Development of Fish Parasite Vaccines in the OMICs Era: Progress and Opportunities



Abstract
1. Introduction
2. Data Sources, Searches and Study Selection
3. Economically Important Fish Parasites
4. Progress in Understanding of Host-Parasite Interactions
4.1. Innate Immune Response during Selected Parasitic Infections
4.2. Adaptive Immune Response during Selected Parasite Infection
5. Parasite Vaccines: Status and Prospects
5.1. Ectoparasite Vaccine Trials
5.2. Endoparasite Vaccine Trials
6. Perspectives in Fish Parasite Vaccine Development
6.1. Vaccination Strategy
6.2. Vaccination Routes
6.3. Protective Immune Response
6.4. Long-Term Immunity
7. Challenges in Vaccine Development
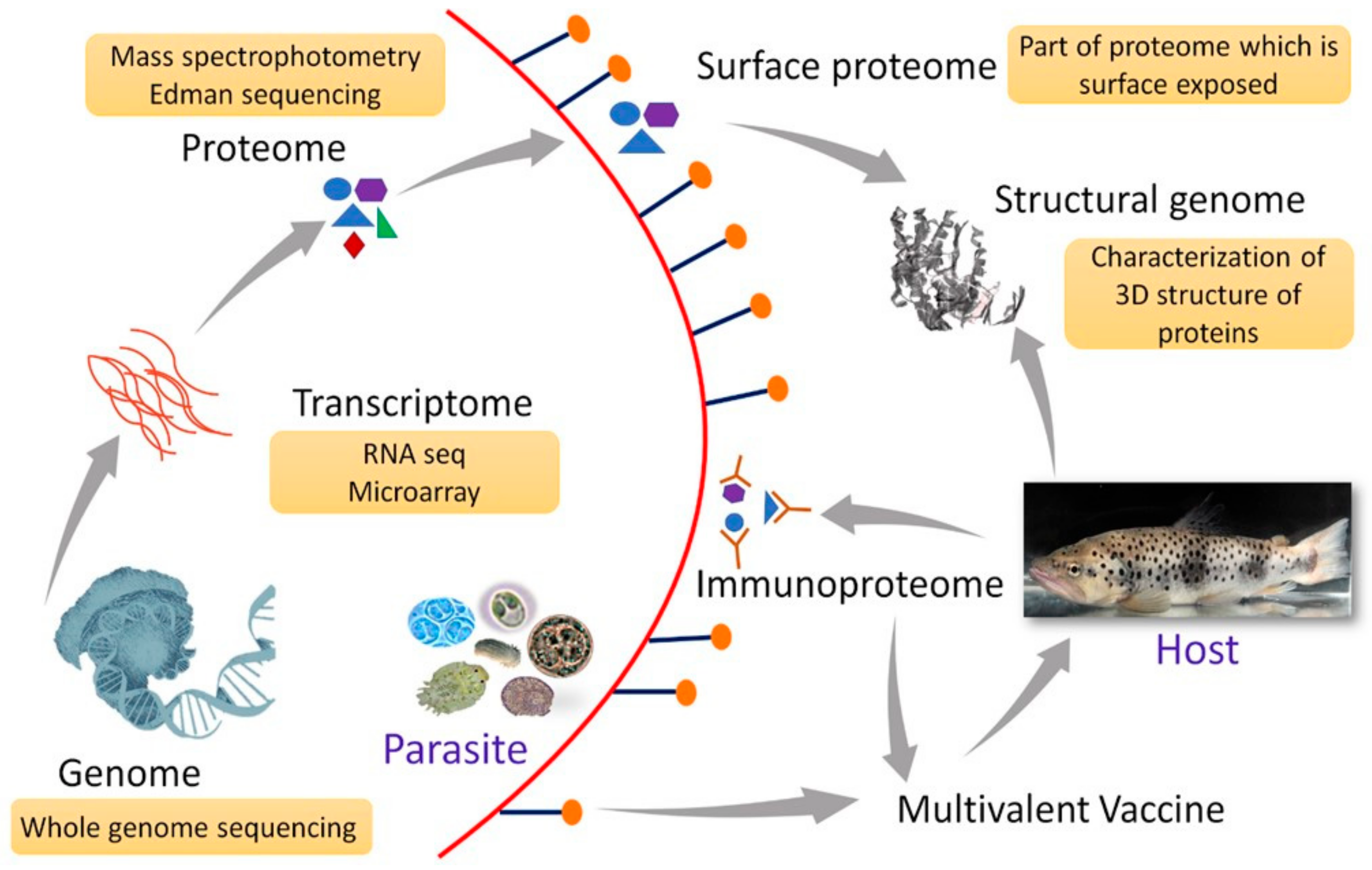
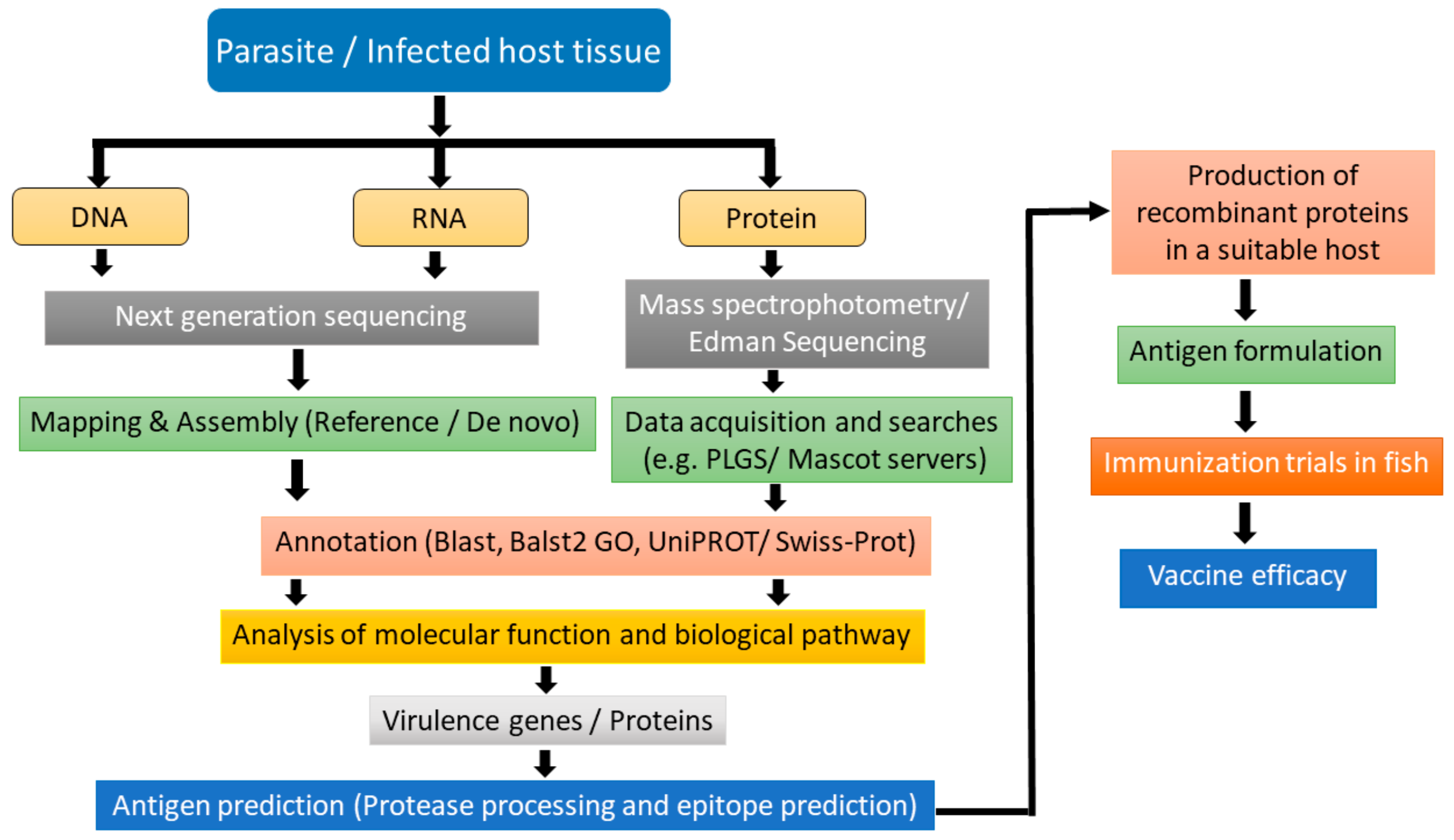
8. Role of Omics Technologies in Vaccine Development
8.1. Transcriptomics
8.2. Proteomics and Genomics
9. Approach of Multivalent Vaccines
10. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- Murray, A.G.; Peeler, E.J. A framework for understanding the potential for emerging diseases in aquaculture. Prev. Vet. Med. 2005, 67, 223–235. [Google Scholar] [CrossRef]
- Shinn, A.P.; Pratoomyot, J.; Bron, J.E.; Paladini, G.; Brooker, E.E.; Brooker, A.J. Economic costs of protistan and metazoan parasites to global mariculture. Parasitology 2015, 142, 196–270. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, H. A review: Gaps in our knowledge on myxozoan parasites of fishes. Fish Pathol. 2003, 38, 125–136. [Google Scholar] [CrossRef]
- Hoffman, G.L. Myxobolus cerebralis, a worldwide cause of salmonid whirling disease. J. Aquat. Anim. Health 1990, 2, 30–37. [Google Scholar] [CrossRef]
- Bravo, S.; Sevatdal, S.; Horsberg, T.E. Sensitivity assessment of Caligus rogercresseyi to emamectin benzoate in Chile. Aquaculture 2008, 282, 7–12. [Google Scholar] [CrossRef]
- Sevatdal, S.; Copley, L.; Wallace, C.; Jackson, D.; Horsberg, T.E. Monitoring of the sensitivity of sea lice (Lepeophtheirus salmonis) to pyrethroids in Norway, Ireland and Scotland using bioassays and probit modelling. Aquaculture 2005, 244, 19–27. [Google Scholar] [CrossRef]
- Boxall, A.B.A. The environmental side effects of medication: How are human and veterinary medicines in soils and water bodies affecting human and environmental health? EMBO Rep. 2004, 5, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.C.; Jürgens, M.D.; Williams, R.J.; Kümmerer, K.; Kortenkamp, A.; Sumpter, J.P. Do cytotoxic chemotherapy drugs discharged into rivers pose a risk to the environment and human health? An overview and UK case study. J. Hydrol. 2008, 348, 167–175. [Google Scholar] [CrossRef]
- Valladão, G.M.R.; Gallani, S.U.; Pilarski, F. Phytotherapy as an alternative for treating fish disease. J. Vet. Pharmacol. Ther. 2015, 38, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Neurol. 2014, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, H.; Ogawa, K.; Wakabayashi, H. Chemotherapy with fumagillin and toltrazuril against kidney enlargement disease of goldfish caused by the myxosporean Hoferellus carassii. Fish. Pathol. 1990, 25, 157–163. [Google Scholar] [CrossRef]
- Morris, D.J.; Adams, A.; Smith, P.; Richards, R.H. Effects of oral treatment with TNP-470 on rainbow trout (Oncorhynchus mykiss) infected with Tetracapsuloides bryosalmonae (Malacosporea), the causative agent of proliferative kidney disease. Aquaculture 2003, 221, 51–64. [Google Scholar] [CrossRef]
- Flamarique, I.N.; Gulbransen, C.; Galbraith, M.; Stucchi, D. Monitoring and potential control of sea lice using an LED-based light trap. Can. J. Fish. Aquat. Sci. 2009, 66, 1371–1382. [Google Scholar] [CrossRef]
- Skilton, D.C.; Saunders, R.J.; Hutson, K.S. Parasite attractants: Identifying trap baits for parasite management in aquaculture. Aquaculture 2020, 516, 734557. [Google Scholar] [CrossRef]
- Adams, A. Progress, challenges and opportunities in fish vaccine development. Fish Shellfish Immunol. 2019, 90, 210–214. [Google Scholar] [CrossRef]
- Villegas, A. First Ever Sea Lice Vaccine Launched in Chile. Available online: https://www.undercurrentnews.com/2015/11/24/first-ever-sea-lice-vaccine-launched-in-chile/ (accessed on 26 February 2020).
- Woo, P.T.K.; Buchmann, K. Fish Parasites:Pathobiology and Protection; CAB International: Wallingford, UK, 2012; ISBN 9781845938062. [Google Scholar]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-omics approaches to disease. Genome Biol. 2017, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kavallaris, M.; Marshall, G.M. Proteomics and disease: Opportunities and challenges. Med. J. Aust. 2005, 182, 575–579. [Google Scholar] [CrossRef]
- Hegde, P.S.; White, I.R.; Debouck, C. Interplay of transcriptomics and proteomics. Curr. Opin. Biotechnol. 2003, 14, 647–651. [Google Scholar] [CrossRef]
- Lowe, R.; Shirley, N.; Bleackley, M.; Dolan, S.; Shafee, T. Transcriptomics technologies. PLoS Comput. Biol. 2017, 13, e1005457. [Google Scholar] [CrossRef]
- Feder, M.E.; Walser, J.C. The biological limitations of transcriptomics in elucidating stress and stress responses. J. Evol. Biol. 2005, 18, 901–910. [Google Scholar] [CrossRef]
- Sudhagar, A.; Kumar, G.; El-Matbouli, M. Transcriptome analysis based on RNA-Seq in understanding pathogenic mechanisms of diseases and the immune system of fish: A comprehensive review. Int. J. Mol. Sci. 2018, 19, 245. [Google Scholar] [CrossRef]
- Feist, S.W.; Longsaw, M. Phylum myxozoa. In Fish Diseases and Disorders; Woo, P.T.K., Ed.; CAB International: Wallingford, UK, 2006; pp. 230–296. [Google Scholar]
- Clifton-Hadley, R.S.; Richards, R.H.; Bucke, D. Proliferative kidney disease (PKD) in rainbow trout Salmo gairdneri: Further observations on the effects of water temperature. Aquaculture 1986, 55, 165–171. [Google Scholar] [CrossRef]
- Rognlie, M.C.; Knapp, S.E. Myxobolus cerebralis in Tubifex tubifex from a Whirling Disease Epizootic in Montana. J. Parasitol. 1998, 84, 711–713. [Google Scholar]
- Lom, J.; Dyková, I.; Pavlásková, M.; Grupcheva, G. Sphaerospora molnari sp.nov. (Myxozoa:Myxosporea) an Agent of Gill, Skin and Blood Sphaerosporosis of Common Carp in Europe. Parasitology 1983, 86, 529–535. [Google Scholar] [CrossRef]
- Körting, W. Larval development of Bothriocephalus sp. (Cestoda: Pseudophyllidea) from carp (Cyprinm carpio L.) in Germany. J. Fish Biol. 1975, 7, 727–733. [Google Scholar] [CrossRef]
- Matthews, R.A. Ichthyophthirius multifiliis fouquet and ichthyophthiriosis in freshwater teleosts. Adv. Parasitol. 2005, 59, 159–241. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, P.K.; Kar, B.; Mohapatra, A.; Mohanty, J. De novo whole transcriptome analysis of the fish louse, Argulus siamensis: First molecular insights into characterization of Toll downstream signalling molecules of crustaceans. Exp. Parasitol. 2013, 135, 629–641. [Google Scholar] [CrossRef]
- Bakke, T.A.; Cable, J.; Harris, P.D. The Biology of Gyrodactylid Monogeneans: The “Russian-Doll Killers”. Adv. Parasitol. 2007, 64. [Google Scholar] [CrossRef]
- Braden, L.M.; Rasmussen, K.J.; Purcell, S.L.; Ellis, L.; Mahony, A.; Cho, S.; Whyte, S.K.; Jones, S.R.M.; Fast, M.D. Acquired protective immunity in Atlantic salmon Salmo salar against the myxozoan Kudoa thyrsites involves induction of MHIIβ+ CD83+ antigen-presenting cells. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef]
- Kent, M.L.; Margolis, L.; Whitaker, D.J.; Hoskins, G.E.; McDonald, T.E. Review of Myxosporea of importance in salmonid fisheries and aquaculture in British Columbia. Folia Parasitol. 1994, 41, 27–37. [Google Scholar]
- Piazzon, M.C.; Estensoro, I.; Calduch-Giner, J.A.; Del Pozo, R.; Picard-Sánchez, A.; Pérez-Sánchez, J.; Sitjà-Bobadilla, A. Hints on T cell responses in a fish-parasite model: Enteromyxum leei induces differential expression of T cell signature molecules depending on the organ and the infection status. Parasites Vectors 2018, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Piazzon, M.C.; Leiro, J.; Lamas, J. Reprint of “Fish immunity to scuticociliate parasites”. Dev. Comp. Immunol. 2014, 43, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.C.; Treasurer, J.W.; Bravo, S.; Nagasawa, K.; Kabata, Z. A review of the impact of parasitic copepods on marine aquaculture. Zool. Stud. 2004, 43, 229–243. [Google Scholar]
- Jones, S.R.M. The occurrence and mechanisms of innate immunity against parasites in fish. Dev. Comp. Immunol. 2001, 25, 841–852. [Google Scholar] [CrossRef]
- Sitja-Bobadilla, A.; Palenzuela, O. Enteromyxum species. In Fish Parasites: Pathobiology and Protection; Woo, P.T.K., Buchmann, K., Eds.; CAB International: Oxfordshire, UK, 2012; p. 163. [Google Scholar]
- Frazer, L.N. Sea-cage aquaculture, sea lice, and declines of wild fish. Conserv. Biol. 2009, 23, 599–607. [Google Scholar] [CrossRef]
- Moller, O.S. Argulus foliaceus. In Fish Parasites: Pathobiology and Protection; Woo, P.T.K., Buchmann, K., Eds.; CAB International: Oxfordshire, UK, 2012; pp. 327–330. [Google Scholar]
- Bartholomew, J.L.; Reno, P.W. The history and dissemination of whirling disease. Am. Fish. Soc. Symp. 2002, 2002, 3–24. [Google Scholar]
- Sudhagar, A.; Kumar, G.; El-Matbouli, M. The malacosporean myxozoan parasite Tetracapsuloides bryosalmonae: A threat to wild salmonids. Pathogens 2020, 9, 16. [Google Scholar] [CrossRef]
- Hallett, S.L.; Ray, R.A.; Hurst, C.N.; Holt, R.A.; Buckles, G.R.; Atkinson, S.D.; Bartholomew, J.L. Density of the waterborne parasite Ceratomyxa shasta and its biological effects on salmon. Appl. Environ. Microbiol. 2012, 78, 3724–3731. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, H.W.; Findly, R.C. Immunity to Ichthyophthirius infections in fish: A synopsis. Dev. Comp. Immunol. 2014, 43, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Barker, S.E.; Bricknell, I.R.; Covello, J.; Purcell, S.; Fast, M.D.; Wolters, W.; Bouchard, D.A. Sea lice, Lepeophtheirus salmonis (Krøyer 1837), infected Atlantic salmon (Salmo salar L.) are more susceptible to infectious salmon anemia virus. PLoS ONE 2019, 14, e0209178. [Google Scholar] [CrossRef]
- Smith, N.C.; Rise, M.L.; Christian, S.L. A Comparison of the Innate and Adaptive Immune Systems in Cartilaginous Fish, Ray-Finned Fish, and Lobe-Finned Fish. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Rauta, P.R.; Nayak, B.; Das, S. Immune system and immune responses in fish and their role in comparative immunity study: A model for higher organisms. Immunol. Lett. 2012, 148, 23–33. [Google Scholar] [CrossRef]
- Picard-Sánchez, A.; Estensoro, I.; del Pozo, R.; Piazzon, M.C.; Palenzuela, O.; Sitjà-Bobadilla, A. Acquired protective immune response in a fish-myxozoan model encompasses specific antibodies and inflammation resolution. Fish Shellfish Immunol. 2019, 90, 349–362. [Google Scholar] [CrossRef]
- Rigos, G.; Pavlidis, M.; Divanach, P. Host susceptibility to Cryptocaryon sp. infection of Mediterranean marine broodfish held under intensive culture conditions: A case report. Bull. Eur. Assoc. Fish. Pathol. 2001, 21, 33–36. [Google Scholar]
- Takano, T.; Kondo, H.; Hirono, I.; Endo, M.; Saito-Taki, T.; Aoki, T. Toll-like receptors in teleosts. In Diseases in Asian Aquaculture VII; Fish Health Section, Asian Fisheries Society: Selangor, Malaysia, 2011; Volume 385, pp. 197–208. [Google Scholar]
- Sudhagar, A.; El-Matbouli, M.; Kumar, G. Identification and expression profiling of toll-like receptors of brown trout (Salmo trutta) during proliferative kidney disease. Int. J. Mol. Sci. 2020, 21, 3755. [Google Scholar] [CrossRef]
- Zhao, F.; Li, Y.W.; Pan, H.J.; Shi, C.B.; Luo, X.C.; Li, A.X.; Wu, S.Q. Expression profiles of toll-like receptors in channel catfish (Ictalurus punctatus) after infection with Ichthyophthirius multifiliis. Fish Shellfish Immunol. 2013, 35, 993–997. [Google Scholar] [CrossRef]
- Li, Y.W.; Xu, D.D.; Li, X.; Mo, Z.Q.; Luo, X.C.; Li, A.X.; Dan, X.M. Identification and characterization of three TLR1 subfamily members from the orange-spotted grouper, Epinephelus coioides. Dev. Comp. Immunol. 2016, 61, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Muñoz, V.; Boltaña, S.; Gallardo-Escárate, C. Comparative immunity of Salmo salar and Oncorhynchus kisutch during infestation with the sea louse Caligus rogercresseyi: An enrichment transcriptome analysis. Fish Shellfish Immunol. 2016, 59, 276–287. [Google Scholar] [CrossRef]
- Becker, I.; Salaiza, N.; Aguirre, M.; Delgado, J.; Carrillo-Carrasco, N.; Kobeh, L.G.; Ruiz, A.; Cervantes, R.; Torres, A.P.; Cabrera, N.; et al. Leishmania lipophosphoglycan (LPG) activates NK cells through toll-like receptor. Mol. Biochem. Parasitol. 2003, 130, 65–74. [Google Scholar] [CrossRef]
- Sepulcre, M.; Pelegrín, P.; Mulero, V.; Meseguer, J. Characterisation of gilthead seabream acidophilic granulocytes by a monoclonal antibody unequivocally points to their involvement in fish phagocytic response. Cell Tissue Res. 2002, 308, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Palikova, M.; Papezikova, I.; Markova, Z.; Navratil, S.; Mares, J.; Mares, L.; Vojtek, L.; Hyrsl, P.; Jelinkova, E.; Schmidt-Posthaus, H. Proliferative kidney disease in rainbow trout (Oncorhynchus mykiss) under intensive breeding conditions: Pathogenesis and haematological and immune parameters. Vet. Parasitol. 2017, 238, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, P.; Álvarez-Pellitero, P.; Sitja-Bobadilla, A. Modulation of the in vitro activity of European sea bass (Dicentrarchus labrax L.) phagocytes by the myxosporean parasite Sphaerospora dicentrarchi (Myxosporea: Bivalvulida). Fish Shellfish Immunol. 2000, 10, 567–581. [Google Scholar] [CrossRef]
- Chin, A.; Woo, P.T.K. Innate cell-mediated immune response and peripheral leukocyte populations in Atlantic salmon, Salmo salar L.; to a live Cryptobia salmositica vaccine. Parasitol. Res. 2005, 95, 299–304. [Google Scholar] [CrossRef]
- Korytář, T.; Wiegertjes, G.F.; Zusková, E.; Tomanová, A.; Lisnerová, M.; Patra, S.; Sieranski, V.; Šíma, R.; Born-Torrijos, A.; Wentzel, A.S.; et al. The kinetics of cellular and humoral immune responses of common carp to presporogonic development of the myxozoan Sphaerospora molnari. Parasites Vectors 2019, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Graves, S.S.; Evans, D.L.; Dawe, D.L. Mobilization and activation of nonspecific cytotoxic cells (ncc) in the channel catfish (Ictalurus punctatus) infected with Ichthyophthirius multifiliis. Comp. Immunol. Microbiol. Infect. Dis. 1985, 8, 43–51. [Google Scholar] [CrossRef]
- Mali, P.; Sanyal, K.B.; Mukherjee, D.; Guchhait, A.; Dash, G. Nonspecific cytotoxic cells (NCC) in fish: A review. J. Interacad. 2017, 21, 372–378. [Google Scholar]
- Jaso-Friedmann, L.; Leary, J.H.; Weisman, Z.; Evans, D.L. Activation of nonspecific cytotoxic cells with a multiple antigenic peptide: Specificity and requirements for receptor crosslinkage. Cell. Immunol. 1996, 170, 195–201. [Google Scholar] [CrossRef]
- Leiro, J.; Piazzón, M.C.; Budiño, B.; Sanmartín, M.L.; Lamas, J. Complement-mediated killing of Philasterides dicentrarchi (Ciliophora) by turbot serum: Relative importance of alternative and classical pathways. Parasite Immunol. 2008, 30, 535–543. [Google Scholar] [CrossRef]
- Agius, C.; Roberts, R.J. Melano-macrophage centres and their role in fish pathology. J. Fish Dis. 2003, 26, 499–509. [Google Scholar] [CrossRef]
- Cuesta, A.; Muñoz, P.; Rodríguez, A.; Salinas, I.; Sitjà-Bobadilla, A.; Álvarez-Pellitero, P.; Esteban, M.A.; Meseguer, J. Gilthead seabream (Sparus aurata L.) innate defence against the parasite Enteromyxum leei (Myxozoa). Parasitology 2006, 132, 95–104. [Google Scholar] [CrossRef]
- Holzer, A.S.; Schachner, O. Myxosporidia and macrophage centres in chub (Leuciscus cephalus)-Quantitative interactions focus on Myxobolus cyprini. Parasitology 2001, 122, 55–62. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Redondo, M.J.; Bermúdez, R.; Palenzuela, O.; Ferreiro, I.; Riaza, A.; Quiroga, I.; Nieto, J.M.; Alvarez-Pellitero, P. Innate and adaptive immune responses of turbot, Scophthalmus maximus (L.), following experimental infection with Enteromyxum scophthalmi (Myxosporea: Myxozoa). Fish Shellfish Immunol. 2006, 21, 485–500. [Google Scholar] [CrossRef] [PubMed]
- Davey, G.C.; Calduch-Giner, J.A.; Houeix, B.; Talbot, A.; Sitjà-Bobadilla, A.; Prunet, P.; Pérez-Sánchez, J.; Cairns, M.T. Molecular profiling of the gilthead sea bream (Sparus aurata L.) response to chronic exposure to the myxosporean parasite Enteromyxum leei. Mol. Immunol. 2011, 48, 2102–2112. [Google Scholar] [CrossRef]
- Baerwald, M.R. Temporal expression patterns of rainbow trout immune-related genes in response to Myxobolus cerebralis exposure. Fish Shellfish Immunol. 2013, 35, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Bjork, S.J.; Zhang, Y.A.; Hurst, C.N.; Alonso-Naveiro, M.E.; Alexander, J.D.; Sunyer, J.O.; Bartholomew, J.L. Defenses of susceptible and resistant Chinook salmon (Onchorhynchus tshawytscha) against the myxozoan parasite Ceratomyxa shasta. Fish Shellfish Immunol. 2014, 37, 87–95. [Google Scholar] [CrossRef][Green Version]
- Wang, B.; Wangkahart, E.; Secombes, C.J.; Wang, T. Insights into the evolution of the suppressors of cytokine signaling (SOCS) gene family in vertebrates. Mol. Biol. Evol. 2019, 36, 393–411. [Google Scholar] [CrossRef]
- Kotob, M.H.; Kumar, G.; Saleh, M.; Gorgoglione, B.; Abdelzaher, M.; El-Matbouli, M. Differential modulation of host immune genes in the kidney and cranium of the rainbow trout (Oncorhynchus mykiss) in response to Tetracapsuloides bryosalmonae and Myxobolus cerebralis co-infections. Parasites Vectors 2018, 11, 1–10. [Google Scholar] [CrossRef]
- Jørgensen, L.v.G. The fish parasite Ichthyophthirius multifiliis—Host immunology, vaccines and novel treatments. Fish Shellfish Immunol. 2017, 67, 586–595. [Google Scholar] [CrossRef]
- Mustafa, A.; MacWilliams, C.; Fernandez, N.; Matchett, K.; Conboy, G.A.; Burka, J.F. Effects of sea lice (Lepeophtheirus salmonis Kröyer, 1837) infestation on macrophage functions in Atlantic salmon (Salmo salar L.). Fish Shellfish Immunol. 2000, 10, 47–59. [Google Scholar] [CrossRef]
- Dalvin, S.; Jørgensen, L.v.G.; Kania, P.W.; Grotmol, S.; Buchmann, K.; Øvergård, A.C. Rainbow trout Oncorhynchus mykiss skin responses to salmon louse Lepeophtheirus salmonis: From copepodid to adult stage. Fish Shellfish Immunol. 2020, 103, 200–210. [Google Scholar] [CrossRef]
- Wallace, J.L. Nitric oxide as a regulator of inflammatory processes. Mem. Inst. Oswaldo Cruz 2005, 100, 5–9. [Google Scholar] [CrossRef]
- Salinas, I.; Zhang, Y.A.; Sunyer, J.O. Mucosal immunoglobulins and B cells of teleost fish. Dev. Comp. Immunol. 2011, 35, 1346–1365. [Google Scholar] [CrossRef] [PubMed]
- Saulnier, D.; De Kinkelin, P. Antigenic and biochemical study of PKX, the myxosporean causative agent of proliferative kidney disease of salmonid fish. Dis. Aquat. Organ. 1996, 27, 103–114. [Google Scholar] [CrossRef]
- Kumar, G.; Abd-Elfattah, A.; El-Matbouli, M. Differential modulation of host genes in the kidney of brown trout Salmo trutta during sporogenesis of Tetracapsuloides bryosalmonae (Myxozoa). Vet. Res. 2014, 45, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, R.P.; El-Matbouli, M.; Adkison, M.A.; MacConnell, E. Whirling disease: Re-emergence among wild trout. Immunol. Rev. 1998, 166, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, P.; Palenzuela, O.; Sitjà-Bobadilla, A.; Álvarez-Pellitero, P. Immunohistochemical reactivity of polyclonal antibodies against Sphaerospora testicularis and Ceratomyxa labracis (Myxosporea: Bivalvulida), with other myxosporean parasites. Int. J. Parasitol. 1999, 29, 521–525. [Google Scholar] [CrossRef]
- Bartholomew, J.L. Host resistance to infection by the myxosporean parasite Ceratomyxa shasta: A review. J. Aquat. Anim. Health 1998, 10, 112–120. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Redondo, M.J.; Macias, M.A.; Ferreiro, I.; Riaza, A.; Alvarez-Pellitero, P. Development of immunohistochemistry and enzyme-linked immunosorbent assays for the detection of circulating antibodies against Enteromyxum scophthalmi (Myxozoa) in turbot (Scophthalmus maximus L.). Fish Shellfish Immunol. 2004, 17, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, X.; Sato, H.; Zhang, Q.; Li, A.; Zhang, J. RNA-seq analysis of local tissue of Carassius auratus gibelio with pharyngeal myxobolosis: Insights into the pharyngeal mucosal immune response in a fish-parasite dialogue. Fish Shellfish Immunol. 2019, 94, 99–112. [Google Scholar] [CrossRef]
- Zhang, Y.A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; Xu, Z.; Lapatra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol. 2010, 11, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Parra, D.; Gómez, D.; Salinas, I.; Zhang, Y.A.; Von Gersdorff Jørgensen, L.; Heinecke, R.D.; Buchmann, K.; LaPatra, S.; Oriol Sunyer, J. Teleost skin, an ancient mucosal surface that elicits gut-like immune responses. Proc. Natl. Acad. Sci. USA 2013, 110, 13097–13102. [Google Scholar] [CrossRef]
- Abos, B.; Estensoro, I.; Perdiguero, P.; Faber, M.; Hu, Y.; Rosales, P.D.; Granja, A.G.; Secombes, C.J.; Holland, J.W.; Tafalla, C. Dysregulation of B cell activity during proliferative kidney disease in rainbow trout. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- MacKinnon, B.M. Host Response of Atlantic Salmon (Salmo salar) to Infection by Sea Lice (Caligus elongatus). Can. J. Fish. Aquat. Sci. 1993, 50, 789–792. [Google Scholar] [CrossRef]
- Nakanishi, T.; Shibasaki, Y.; Matsuura, Y. T cells in fish. Biology 2015, 4, 640–663. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.; Montero, R.; Kumar, G.; Sudhagar, A.; Friedl, A.; Köllner, B.; El-Matbouli, M. Kinetics of local and systemic immune cell responses in whirling disease infection and resistance in rainbow trout. Parasites Vectors 2019, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tarleton, R.L. New approaches in vaccine development for parasitic infections. Cell. Microbiol. 2005, 7, 1379–1386. [Google Scholar] [CrossRef]
- Clem, A.S. Fundamentals of vaccine immunology. J. Glob. Infect. Dis. 2011, 3, 73–78. [Google Scholar] [CrossRef]
- Goven, B.A.; Dawe, D.L.; Gratzek, J.B. Protection of channel catfish, Ictalurus punctatus Rafmesque, against Ichthyophthirius multifiliis Fouquet by immunization. J. Fish. Biol. 1980, 17, 311–316. [Google Scholar] [CrossRef]
- Gibbons, I.R. Studies on the Protein Components of Cilia From Tetrahymena pyriformis. Proc. Natl. Acad. Sci. USA 1963, 50, 1002–1010. [Google Scholar] [CrossRef]
- Sleigh, M.A. Cilia and Flagella; Academic Press: London, UK, 1974. [Google Scholar]
- Burkart, M.A.; Clark, T.G.; Dickerson, H.W. Immunization of channel catfish, Ictalurus punctatus Rafinesque, against Ichthyophthirius multifiliis (Fouquet): Killed versus live vaccines. J. Fish Dis. 1990, 13, 401–410. [Google Scholar] [CrossRef]
- Xu, D.H.; Zhang, D.; Shoemaker, C.; Beck, B. Immune response of channel catfish (Ictalurus punctatus) against Ichthyophthirius multifiliis post vaccination using DNA vaccines encoding immobilization antigens. Fish Shellfish Immunol. 2019, 94, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.H.; Klesius, P.H. Comparison of serum antibody responses and host protection against parasite Ichthyophthirius multifiliis between channel catfish and channel × blue hybrid catfish. Fish Shellfish Immunol. 2013, 34, 1356–1359. [Google Scholar] [CrossRef]
- Von Gersdorff Jørgensen, L.; Kania, P.W.; Rasmussen, K.J.; Mattsson, A.H.; Schmidt, J.; Al-Jubury, A.; Sander, A.; Salanti, A.; Buchmann, K. Rainbow trout (Oncorhynchus mykiss) immune response towards a recombinant vaccine targeting the parasitic ciliate Ichthyophthirius multifiliis. J. Fish. Dis. 2017, 40, 1815–1821. [Google Scholar] [CrossRef]
- von Gersdorff Jørgensen, L.; Sigh, J.; Kania, P.W.; Holten-Andersen, L.; Buchmann, K.; Clark, T.; Rasmussen, J.S.; Einer-Jensen, K.; Lorenzen, N. Approaches towards DNA Vaccination against a Skin Ciliate Parasite in Fish. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Jørgensen, L.V.G.; Nemli, E.; Heinecke, R.D.; Raida, M.K.; Buchmann, K. Immune-relevant genes expressed in rainbow trout following immunisation with a live vaccine against Ichthyophthirius multifiliis. Dis. Aquat. Organ. 2008, 80, 189–197. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dalgaard, M.; Buchmann, K.; Li, A. Immunization of rainbow trout fry with Ichthyophthirius multifiliis sonicate: Protection of host and immunological changes. Bull. Eur. Assoc. Fish Pathol. 2002, 22, 288–297. [Google Scholar]
- Xu, D.H.; Klesius, P.H.; Shoemaker, C.A. Protective immunity of Nile tilapia against Ichthyophthirius multifiliis post-immunization with live theronts and sonicated trophonts. Fish Shellfish Immunol. 2008, 25, 124–127. [Google Scholar] [CrossRef]
- Yambot, A.V.; Song, Y.L. Immunization of grouper, Epinephelus coioides, confers protection against a protozoan parasite, Cryptocaryon irritans. Aquaculture 2006, 260, 1–9. [Google Scholar] [CrossRef]
- Grayson, T.H.; John, R.J.; Wadsworth, S.; Greaves, K.; Cox, D.; Roper, J.; Wrathmell, A.B.; Gilpin, M.L.; Harris, J.E. Immunization of Atlantic salmon against the salmon louse: Identification of antigens and effects on louse fecundity. J. Fish Biol. 1995, 47, 85–94. [Google Scholar] [CrossRef]
- Faber, M.N.; Holland, J.W.; Secombes, C.J. Vaccination strategies and IgM responses against PKD in rainbow trout. Fish Shellfish Immunol. 2019, 91, 423. [Google Scholar] [CrossRef]
- Kane, S.N.; Mishra, A.; Dutta, A.K. Preface: International Conference on Recent Trends in Physics (ICRTP 2016). J. Phys. Conf. Ser. 2016, 755. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.S. Poly d,l-lactide-co-glycolic acid (PLGA)-encapsulated vaccine on immune system in Epinephelus bruneus against Uronema marinum. Exp. Parasitol. 2012, 131, 325–332. [Google Scholar] [CrossRef]
- Fontenla, F.; Blanco-Abad, V.; Pardo, B.G.; Folgueira, I.; Noia, M.; Gómez-Tato, A.; Martínez, P.; Leiro, J.M.; Lamas, J. Vaccine-induced modulation of gene expression in turbot peritoneal cells. A microarray approach. Mol. Immunol. 2016, 75, 188–199. [Google Scholar] [CrossRef]
- Méndez, S.; Gurunathan, S.; Kamhawi, S.; Belkaid, Y.; Moga, M.A.; Skeiky, Y.A.W.; Campos-Neto, A.; Reed, S.; Seder, R.A.; Sacks, D. The Potency and Durability of DNA- and Protein-Based Vaccines Against Leishmania major Evaluated Using Low-Dose, Intradermal Challenge. J. Immunol. 2001, 166, 5122–5128. [Google Scholar] [CrossRef]
- Knox, D.P.; Redmond, D.L. Parasite vaccines-Recent progress and problems associated with their development. Parasitology 2006, 133. [Google Scholar] [CrossRef]
- Mutoloki, S.; Munang’andu, H.M.; Evensen, Ø. Oral vaccination of fish-antigen preparations, uptake, and immune induction. Front. Immunol. 2015, 6, 1–10. [Google Scholar] [CrossRef]
- Bøgwald, J.; Dalmo, R.A. Review on immersion vaccines for fish: An update. Microorganisms 2019, 7, 627. [Google Scholar] [CrossRef]
- Plant, K.P.; LaPatra, S.E. Advances in fish vaccine delivery. Dev. Comp. Immunol. 2011, 35, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Horne, M.T. Technical aspects of the administration of vaccines. Dev. Biol. Stand. 1997, 90, 79–89. [Google Scholar]
- Sitjà-Bobadilla, A.; Estensoro, I.; Pérez-Sánchez, J. Immunity to gastrointestinal microparasites of fish. Dev. Comp. Immunol. 2016, 64, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.T.K. Cryptobia (Trypanoplasma) salmositica and salmonid cryptobiosis. J. Fish. Dis. 2003, 26, 627–646. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Palenzuela, O.; Alvarez-Pellitero, P. Immune response of turbot, Psetta maxima (L.) (Pisces: Teleostei), to formalin-killed scuticociliates (Ciliophora) and adjuvanted formulations. Fish Shellfish Immunol. 2008, 24, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Buchmann, K.; Lindenstrøm, T.; Bresciani, J. Defence mechanisms against parasites in fish and the prospect for vaccines. Acta Parasitol 2001, 46, 76–81. [Google Scholar]
- Katzenback, B.A.; Plouffe, D.A.; Haddad, G.; Belosevic, M. Administration of recombinant parasite β-tubulin to goldfish (Carassius auratus L.) confers partial protection against challenge infection with Trypanosoma danilewskyi Laveran and Mesnil. Vet. Parasitol. 2008, 151, 36–45. [Google Scholar] [CrossRef]
- Buchmann, K. Immune response to Ichthyophthirius multifiliis and role of IgT. Parasite Immunol. 2020, 42, 2–7. [Google Scholar] [CrossRef]
- Mo, Z.Q.; Li, Y.W.; Wang, H.Q.; Wang, J.L.; Ni, L.Y.; Yang, M.; Lao, G.F.; Luo, X.C.; Li, A.X.; Dan, X.M. Comparative transcriptional profile of the fish parasite Cryptocaryon irritans. Parasites Vectors 2016, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Piazzon, M.C.; Wiegertjes, G.F.; Leiro, J.; Lamas, J. Turbot resistance to Philasterides dicentrarchi is more dependent on humoral than on cellular immune responses. Fish Shellfish Immunol. 2011, 30, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, K.L.; Khader, S.A. Novel vaccine approaches for protection against intracellular pathogens. Curr. Opin. Immunol. 2014, 28, 58–63. [Google Scholar] [CrossRef]
- Bethony, J.; Loukas, A.; Smout, M.; Brooker, S.; Mendez, S.; Plieskatt, J.; Goud, G.; Bottazzi, M.E.; Zhan, B.; Wang, Y.; et al. Antibodies against a secreted protein from hookworm larvae reduce the intensity of hookworm infection in humans and vaccinated laboratory animals. FASEB J. 2005, 19, 1743–1745. [Google Scholar] [CrossRef] [PubMed]
- Riveau, G.; Deplanque, D.; Remoué, F.; Schacht, A.M.; Vodougnon, H.; Capron, M.; Thiry, M.; Martial, J.; Libersa, C.; Capron, A. Safety and immunogenicity of rSh28GST antigen in humans: Phase 1 randomized clinical study of a vaccine candidate against urinary schistosomiasis. PLoS Negl. Trop. Dis. 2012, 6, e1704. [Google Scholar] [CrossRef] [PubMed]
- Lofthouse, S.A.; Andrews, A.E.; Elhay, M.J.; Bowles, V.M.; Meeusen, E.N.T.; Nash, A.D. Cytokines as adjuvants for ruminant vaccines. Int. J. Parasitol. 1996, 26, 835–842. [Google Scholar] [CrossRef]
- Brown, G.V.; Reeder, J.C. Malaria vaccines. Med. J. Aust. 2002, 177, 230–231. [Google Scholar] [CrossRef] [PubMed]
- Arkoosh, M.R.; Kaattari, S.L. Development of immunological memory in rainbow trout (Oncorhynchus mykiss). I. An immunochemical and cellular analysis of the B cell response. Dev. Comp. Immunol. 1991, 15, 279–293. [Google Scholar] [CrossRef]
- Ferguson, H.W.; BalL, H.J. Epidemiological aspects of proliferative kidney disease amongst rainbow trout Salmo gairdneri Richardson in Northern Ireland. J. Fish Dis. 1979, 2, 219–225. [Google Scholar] [CrossRef]
- Findly, R.C.; Zhao, X.; Noe, J.; Camus, A.C.; Dickerson, H.W. B cell memory following infection and challenge of channel catfish with Ichthyophthirius multifiliis. Dev. Comp. Immunol. 2013, 39, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Gorgoglione, B.; Wang, T.; Secombes, C.J.; Holland, J.W. Immune gene expression profiling of Proliferative Kidney Disease in rainbow trout Oncorhynchus mykiss reveals a dominance of anti-inflammatory, antibody and T helper cell-like activities. Vet. Res. 2013, 44, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Fast, M.D. Fish immune responses to parasitic copepod (namely sea lice) infection. Dev. Comp. Immunol. 2014, 43, 300–312. [Google Scholar] [CrossRef]
- Dzikowski, R.; Deitsch, K. Antigenic variation by protozoan parasites: Insights from Babesia bovis. Mol. Microbiol. 2006, 59, 364–366. [Google Scholar] [CrossRef]
- El-Matbouli, M.; Hoffmann, R.W.; Schoel, H.; McDowell, T.S.; Hedrick, R.P. Whirling disease: Host specificity and interaction between the actinosporean stage of Myxobolus cerebralis and rainbow trout Oncorhynchus mykiss. Dis. Aquat. Organ. 1999, 35, 1–12. [Google Scholar] [CrossRef]
- Clark, T.G.; Mcgraw, R.A.; Dickerson, H.W. Developmental expression of surface antigen genes in the parasitic ciliate Ichthyophthirius multifiliis. Proc. Natl. Acad. Sci. USA 1992, 89, 6363–6367. [Google Scholar] [CrossRef]
- Palti, Y. Toll-like receptors in bony fish: From genomics to function. Dev. Comp. Immunol. 2011, 35, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Nóbrega de Sousa, T.; de Menezes Neto, A.; Alves de Brito, C.F. “Omics” in the study of the major parasitic diseases malaria and schistosomiasis. Infect. Genet. Evol. 2013, 19, 258–273. [Google Scholar] [CrossRef]
- Amambua-Ngwa, A.; Tetteh, K.K.A.; Manske, M.; Gomez-Escobar, N.; Stewart, L.B.; Deerhake, M.E.; Cheeseman, I.H.; Newbold, C.I.; Holder, A.A.; Knuepfer, E.; et al. Population Genomic Scan for Candidate Signatures of Balancing Selection to Guide Antigen Characterization in Malaria Parasites. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef]
- Hull, R.; Dlamini, Z. The role played by alternative splicing in antigenic variability in human endo-parasites. Parasites Vectors 2014, 7, 1–19. [Google Scholar] [CrossRef]
- Yeoh, L.M.; Goodman, C.D.; Mollard, V.; McHugh, E.; Lee, V.V.; Sturm, A.; Cozijnsen, A.; McFadden, G.I.; Ralph, S.A. Alternative splicing is required for stage differentiation in malaria parasites. Genome Biol. 2019, 20. [Google Scholar] [CrossRef]
- Haase, S.; Cabrera, A.; Langer, C.; Treeck, M.; Struck, N.; Herrmann, S.; Jansen, P.W.; Bruchhaus, I.; Bachmann, A.; Dias, S.; et al. Characterization of a conserved rhoptry-associated leucine zipper-like protein in the malaria parasite Plasmodium falciparum. Infect. Immun. 2008, 76, 879–887. [Google Scholar] [CrossRef]
- Lindner, S.E.; Swearingen, K.E.; Harupa, A.; Vaughan, A.M.; Sinnis, P.; Moritz, R.L.; Kappe, S.H.I. Total and putative surface proteomics of malaria parasite salivary gland sporozoites. Mol. Cell. Proteom. 2013, 12, 1127–1143. [Google Scholar] [CrossRef]
- Doolan, D.L. Plasmodium Immunomics. Int. J. Parasitol 2011, 41, 3–20. [Google Scholar] [CrossRef]
- Doolan, B.D.L.; Sedegah, M.; Hedstrom, R.C.; Charoenvit, P.H.; Hoffrnan, S.L. Circumventing Genetic Restriction of Protection against Malaria with Multigene DNA Immunization: CD8 + T Cell, Interferon, and Nitric Oxide-Dependent Immunity. J. Exp. Med. 1996, 183. [Google Scholar] [CrossRef] [PubMed]
- Osier, F.H.A.; Fegan, G.; Polley, S.D.; Murungi, L.; Verra, F.; Tetteh, K.K.A.; Lowe, B.; Mwangi, T.; Bull, P.C.; Thomas, A.W.; et al. Breadth and magnitude of antibody responses to multiple Plasmodium falciparum merozoite antigens are associated with protection from clinical malaria. Infect. Immun. 2008, 76, 2240–2248. [Google Scholar] [CrossRef] [PubMed]
- Cecílio, P.; Pérez-Cabezas, B.; Fernández, L.; Moreno, J.; Carrillo, E.; Requena, J.M.; Fichera, E.; Reed, S.G.; Coler, R.N.; Kamhawi, S.; et al. Pre-clinical antigenicity studies of an innovative multivalent vaccine for human visceral leishmaniasis. PLoS Negl. Trop. Dis. 2017, 11, e0005951. [Google Scholar] [CrossRef]
- Jayaraj, R.; Piedrafita, D.; Dynon, K.; Grams, R.; Spithill, T.W.; Smooker, P.M. Vaccination against fasciolosis by a multivalent vaccine of stage-specific antigens. Vet. Parasitol. 2009, 160, 230–236. [Google Scholar] [CrossRef]
- Kumar, G.; Kocour, M. Applications of next-generation sequencing in fisheries research: A review. Fish. Res. 2017, 186, 11–22. [Google Scholar] [CrossRef]
- Sudhagar, A.; Ertl, R.; Kumar, G.; El-Matbouli, M. Transcriptome profiling of posterior kidney of brown trout, Salmo trutta, during proliferative kidney disease. Parasites Vectors 2019, 12, 1–17. [Google Scholar] [CrossRef]
- Kumar, G.; Ertl, R.; Bartholomew, J.L.; El-Matbouli, M. First transcriptome analysis of bryozoan Fredericella sultana, the primary host of myxozoan parasite Tetracapsuloides bryosalmonae. PeerJ 2020. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Ertl, R.; Bartholomew, J.L.; El-Matbouli, M. Transcriptome analysis elucidates the key responses of bryozoan Fredericella sultana during the development of Tetracapsuloides bryosalmonae (Myxozoa). Int. J. Mol. Sci. 2020, 21, 5910. [Google Scholar] [CrossRef]
- Kumar, G.; Ertl, R.; Nilsen, F.; Bartholomew, J.L.; El-Matbouli, M. Data of de novo transcriptome assembly of the myxozoan parasite Tetracapsuloides bryosalmonae. Data Br. 2021, 35, 106831. [Google Scholar] [CrossRef]
- Ronza, P.; Robledo, D.; Bermúdez, R.; Losada, A.P.; Pardo, B.G.; Sitjà-Bobadilla, A.; Quiroga, M.I.; Martínez, P. RNA-seq analysis of early enteromyxosis in turbot (Scophthalmus maximus): New insights into parasite invasion and immune evasion strategies. Int. J. Parasitol. 2016, 46, 507–517. [Google Scholar] [CrossRef]
- Hartigan, A.; Kosakyan, A.; Pecková, H.; Eszterbauer, E.; Holzer, A.S. Transcriptome of Sphaerospora molnari (Cnidaria, Myxosporea) blood stages provides proteolytic arsenal as potential therapeutic targets against sphaerosporosis in common carp. BMC Genom. 2020, 21, 404. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.S.; Neuhof, M.; Rubinstein, N.D.; Diamant, A.; Philippe, H.; Huchon, D.; Cartwright, P. Genomic insights into the evolutionary origin of Myxozoa within Cnidaria. Proc. Natl. Acad. Sci. USA 2015, 112, 14912–14917. [Google Scholar] [CrossRef] [PubMed]
- Mo, Z.Q.; Yang, M.; Wang, H.Q.; Xu, Y.; Huang, M.Z.; Lao, G.F.; Li, Y.W.; Li, A.X.; Luo, X.C.; Dan, X.M. Grouper (Epinephelus coioides) BCR signaling pathway was involved in response against Cryptocaryon irritans infection. Fish Shellfish Immunol. 2016, 57, 198–205. [Google Scholar] [CrossRef]
- Kumar, G.; Gotesman, M.; El-Matbouli, M. Interaction of Tetracapsuloides bryosalmonae, the causative agent of proliferative kidney disease, with host proteins in the kidney of Salmo trutta. Parasitol. Res. 2015, 114, 1721–1727. [Google Scholar] [CrossRef] [PubMed]
- Piriatinskiy, G.; Atkinson, S.D.; Park, S.; Morgenstern, D.; Brekhman, V.; Yossifon, G.; Bartholomew, J.L.; Lotan, T. Functional and proteomic analysis of Ceratonova shasta (Cnidaria: Myxozoa) polar capsules reveals adaptations to parasitism. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Saleh, M.; Kumar, G.; Abdel-Baki, A.A.S.; Dkhil, M.A.; El-Matbouli, M.; Al-Quraishy, S. Quantitative proteomic profiling of immune responses to Ichthyophthirius multifiliis in common carp skin mucus. Fish Shellfish Immunol. 2019, 84, 834–842. [Google Scholar] [CrossRef]
- Mai, Y.Z.; Li, Y.W.; Li, R.J.; Li, W.; Huang, X.Z.; Mo, Z.Q.; Li, A.X. Proteomic analysis of differentially expressed proteins in the marine fish parasitic ciliate Cryptocaryon irritans. Vet. Parasitol. 2015, 211, 1–11. [Google Scholar] [CrossRef]
- Coyne, R.S.; Hannick, L.; Shanmugam, D.; Hostetler, J.B.; Brami, D.; Joardar, V.S.; Johnson, J.; Radune, D.; Singh, I.; Badger, J.H.; et al. Comparative genomics of the pathogenic ciliate Ichthyophthirius multifiliis, its free-living relatives and a host species provide insights into adoption of a parasitic lifestyle and prospects for disease control. Genome Biol. 2011, 12, 1–26. [Google Scholar] [CrossRef]
- Chen, K.; Wang, J.L.; Huang, S.Y.; Yang, W.B.; Zhu, W.N.; Zhu, X.Q. Immune responses and protection after DNA vaccination against Toxoplasma gondii calcium-dependent protein kinase 2 (TgCDPK2). Parasite 2017, 24. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Zhou, A.; Lu, G.; Meng, M.; Sun, M.; Bai, Y.; Han, Y.; Wang, L.; Zhou, H.; Cong, H.; et al. Identification and characterization of Toxoplasma gondii aspartic protease 1 as a novel vaccine candidate against toxoplasmosis. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef]
- Overington, J.P.; Al-Lazikani, B.; Hopkins, A.L. How many drug targets are there? Nat. Rev. Drug Discov. 2006, 5, 993–996. [Google Scholar] [CrossRef] [PubMed]
- National Centre for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA241036/ (accessed on 14 April 2020).
- National Centre for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/sra/SRX4615721/ (accessed on 14 April 2020).
- Hahn, C.; Fromm, B.; Bachmann, L. Comparative genomics of flatworms (Platyhelminthes) reveals shared genomic features of ecto-and endoparastic neodermata. Genome Biol. Evol. 2014, 6, 1105–1117. [Google Scholar] [CrossRef]
- National Centre for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/sra/?term=SRX2417163 (accessed on 14 April 2020).
- National Centre for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/sra/?term=SRX2417144 (accessed on 14 April 2020).
- Ivory, C.; Chadee, K. DNA vaccines: Designing strategies against parasitic infections. Genet. Vaccines Ther. 2004, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Budiño, B.; Lamas, J.; Pata, M.P.; Arranz, J.A.; Sanmartín, M.L.; Leiro, J. Intraspecific variability in several isolates of Philasterides dicentrarchi (syn. Miamiensis avidus), a scuticociliate parasite of farmed turbot. Vet. Parasitol. 2011, 175, 260–272. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parasite Group | Parasite | Disease | Host | Mortality a/ Economic Loss b | Reference |
|---|---|---|---|---|---|
| Freshwater Endoparasites | |||||
| Cnidaria | T. bryosalmonae | Proliferative kidney disease | Rainbow trout | 95% a | [26] |
| M. cerebralis | Whirling disease | Rainbow trout | 90% a | [27] | |
| S. molnari | Sphaerosporosis | Carps | NA | [28] | |
| Platyhelminthes | Bothriocephalus acheilo gnathi | Ulcer, catarrhal enteritis | Carps | 100% a | [29] |
| Freshwater Ectoparasites | |||||
| Ciliophora | I. multifiliis | White spot | Rainbow trout, Carps | NA | [30] |
| Arthropoda | Argulus | Argulosis | Indian Major Carps | 5.41 million USD b | [31] |
| Platyhelminthes | Gyrodactylus salaris | Gyrodactylosis | Atlantic salmon | NA | [32] |
| Marine Endoparasites | |||||
| Cnidaria | K. thyrsites | Soft flesh syndrome | Salmonids | 6 million CAD b | [33] |
| C. shashta | Ceratomyxosis | Salmonids | NA | [34] | |
| E. leei | Enteromyxosis | Sharpsnout seabream | NA | [35] | |
| Ciliophora | Uronema marinum, Philasterides dicentrarchi | Scuticociliatosis | Seabass, Turbot | NA | [36] |
| Marine Ectoparasites | |||||
| Ciliophora | C. irritans | White spot | Greater Amberjack | 834 USD b | [3] |
| Arthropoda | Caligus rogercresseyi, Lepeophtheirus salmonis | Sea louse disease | Salmonids | 100 million USD b | [37] |
| Disease | Parasite | Fish Host | Vaccine Trial Type | Antigen Target | Delivery Method | Reference |
|---|---|---|---|---|---|---|
| White spot disease (Ich) | Ichthyophthirius multifiliis | Channel catfish | DNA | (i-antigen) immobilization antigen | IM | [99] |
| Channel catfish | Live | Theront | IP | [100] | ||
| Rainbow trout | Subunit | Recombinant proteins (#5, #10, & #11) | IP | [101] | ||
| DNA | (i-antigen) immobilization antigen & Cysteine protease | IM, Needle free injection, Gene gun delivery | [102] | |||
| Live | Theronts | IP | [103] | |||
| Killed | Sonicated formalin killed trophonts | IP | [104] | |||
| Nile Tilapia | Live | Theront and sonicated trophonts | Immersion and IP | [105] | ||
| Channel catfish | Live | Tomites | IP | [98] | ||
| Killed | Trophonts and tomites | IP | ||||
| Marine White spot | Cryptocaryon irritans | Grouper | Live | Theronts | Bath | [106] |
| Killed | Formalin-killed theronts | IP | [107] | |||
| Sea louse infestations | Lepeophtheirus salmonis | Atlantic salmon | Crude parasite extract | Adult female parasite | IP |
| Disease | Parasite | Fish Host | Vaccine Type | Antigen Target | Delivery Method | Reference |
|---|---|---|---|---|---|---|
| Freshwater Endoparasites | ||||||
| PKD | Tetracapsuloides bryosalmonae | Rainbow trout | DNA | Micro-exon gene (TB-MEG1) | Not Available | [108] |
| Myxobolosis | Myxobolus koi | Gold fish | Subunit | Crude protein spore | Immersion | [109] |
| Marine Endoparasites | ||||||
| Scuticociliatosis | Uronema marinum | Grouper | Subunit | (i-antigen) immobilization antigen | IP | [110] |
| Philasterides dicentrarchi | Turbot | Subunit | Membrane proteins | IP | [111] | |
| Parasite | Molecular Data Type | Sequencing Platform | Size/No. of Contigs | Accession No. | Reference |
|---|---|---|---|---|---|
| Endoparasite | |||||
| Tetracapsuloides bryosalmonae | Transcriptome | NextSeq 550 | 25908 c | [155] | |
| Myxobolus cerebralis | Transcriptome | HiSeq 2000 | 52972 c | GBGI00000000 | [158] |
| Kudoa iwatai | Transcriptome | HiSeq 2000 | 1637 c | GBKL00000000 | [158] |
| Ceratomyxa shasta | Transcriptome | Illumina HiSeq 3000 | NA | SRX3741971 | [168] |
| Sphaerospora molnari | Transcriptome | Illumina HiSeq | 29560 c | PRJNA522909 | [157] |
| Myxobolus squamalis | Transcriptome | Illumina HiSeq 3000 | NA | SRX4615721 | [169] |
| Ectoparasite | |||||
| Gyrodactylus salaris | Whole genome | Roche 454 FLX Titanium Illumina GAII | 120 Mb b | JJOG00000000 | [170] |
| Cryptocaryon irritans | Transcriptome (Trophont) | Illumina HiSeq 2000 | 2.6 Gb a | SRX2417163 | [171] |
| Transcriptome (Theront) | Illumina HiSeq 2000 | 3.2 Gb a | SRX2417144 | [172] | |
| Argulus siamensis | Transcriptome | Illumina HiSeq 2000 | 46352 c | SRX150806 | [31] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shivam, S.; El-Matbouli, M.; Kumar, G. Development of Fish Parasite Vaccines in the OMICs Era: Progress and Opportunities. Vaccines 2021, 9, 179. https://doi.org/10.3390/vaccines9020179
Shivam S, El-Matbouli M, Kumar G. Development of Fish Parasite Vaccines in the OMICs Era: Progress and Opportunities. Vaccines. 2021; 9(2):179. https://doi.org/10.3390/vaccines9020179
Chicago/Turabian StyleShivam, Saloni, Mansour El-Matbouli, and Gokhlesh Kumar. 2021. "Development of Fish Parasite Vaccines in the OMICs Era: Progress and Opportunities" Vaccines 9, no. 2: 179. https://doi.org/10.3390/vaccines9020179
APA StyleShivam, S., El-Matbouli, M., & Kumar, G. (2021). Development of Fish Parasite Vaccines in the OMICs Era: Progress and Opportunities. Vaccines, 9(2), 179. https://doi.org/10.3390/vaccines9020179