Impact of Helminth Infections during Pregnancy on Vaccine Immunogenicity in Gabonese Infants


 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Vaccination Schedule
2.3. Assessment of Parasite Infection
2.4. Assessment of Vaccine-Specific Antibody Levels (Immunoglobulin IgG)
2.5. Ethical Approval
2.6. Statistical Analysis
3. Results
3.1. Baseline Characteristics
3.2. Assessment of Vaccine-Specific IgG Levels
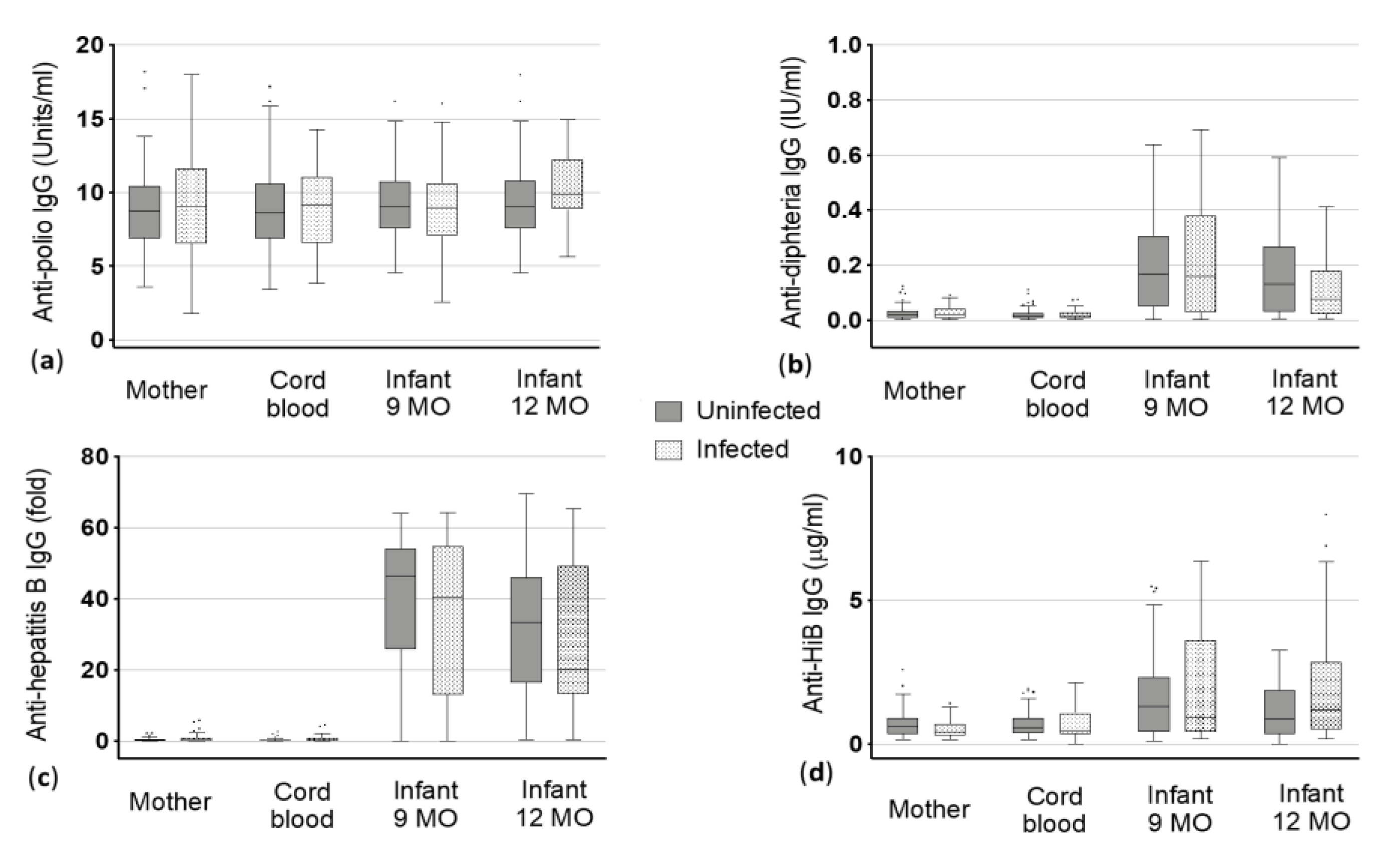
3.2.1. Polio
3.2.2. Diphtheria
3.2.3. Hepatitis B
3.2.4. HiB
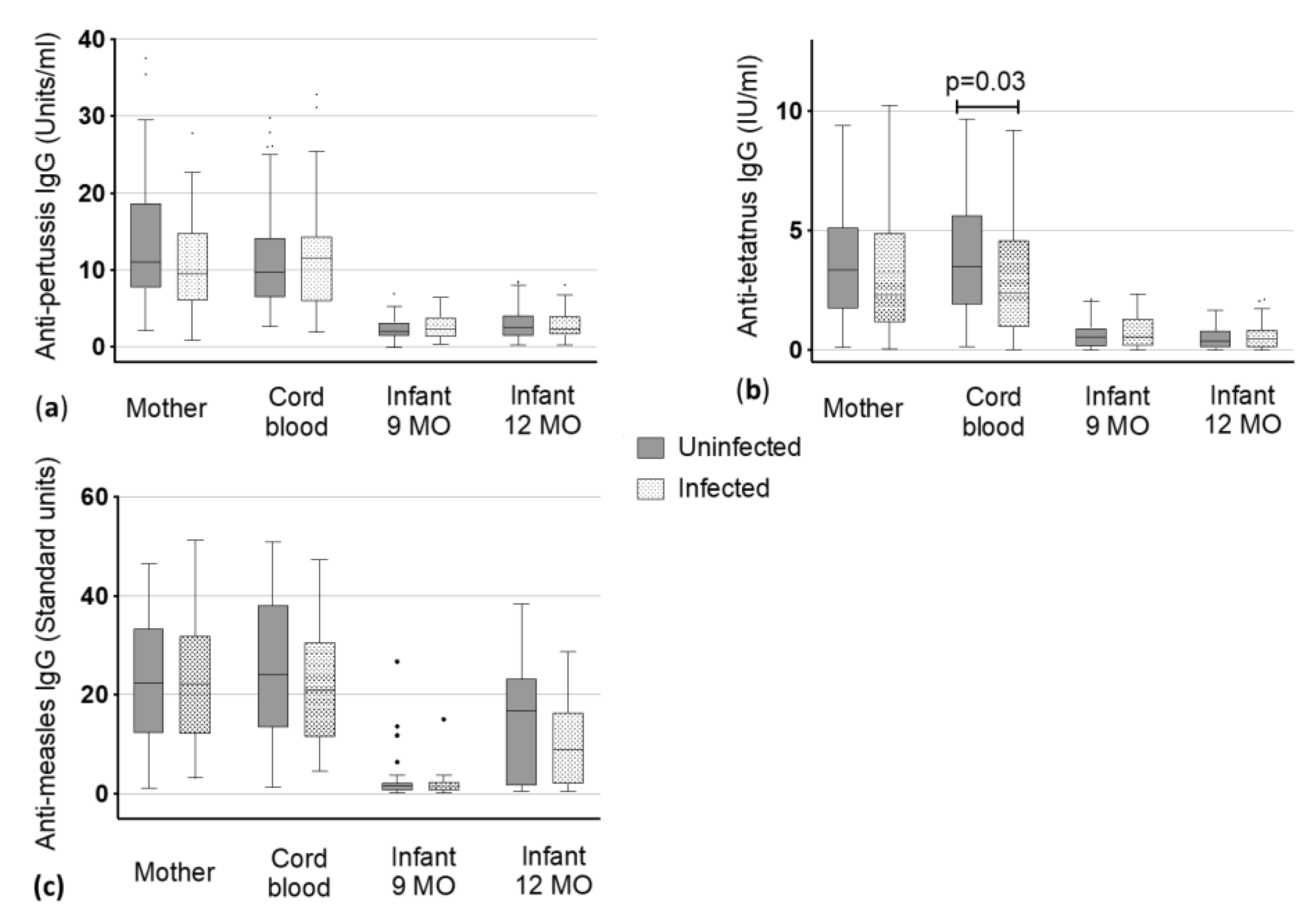
3.2.5. Pertussis
3.2.6. Tetanus
3.2.7. Measles
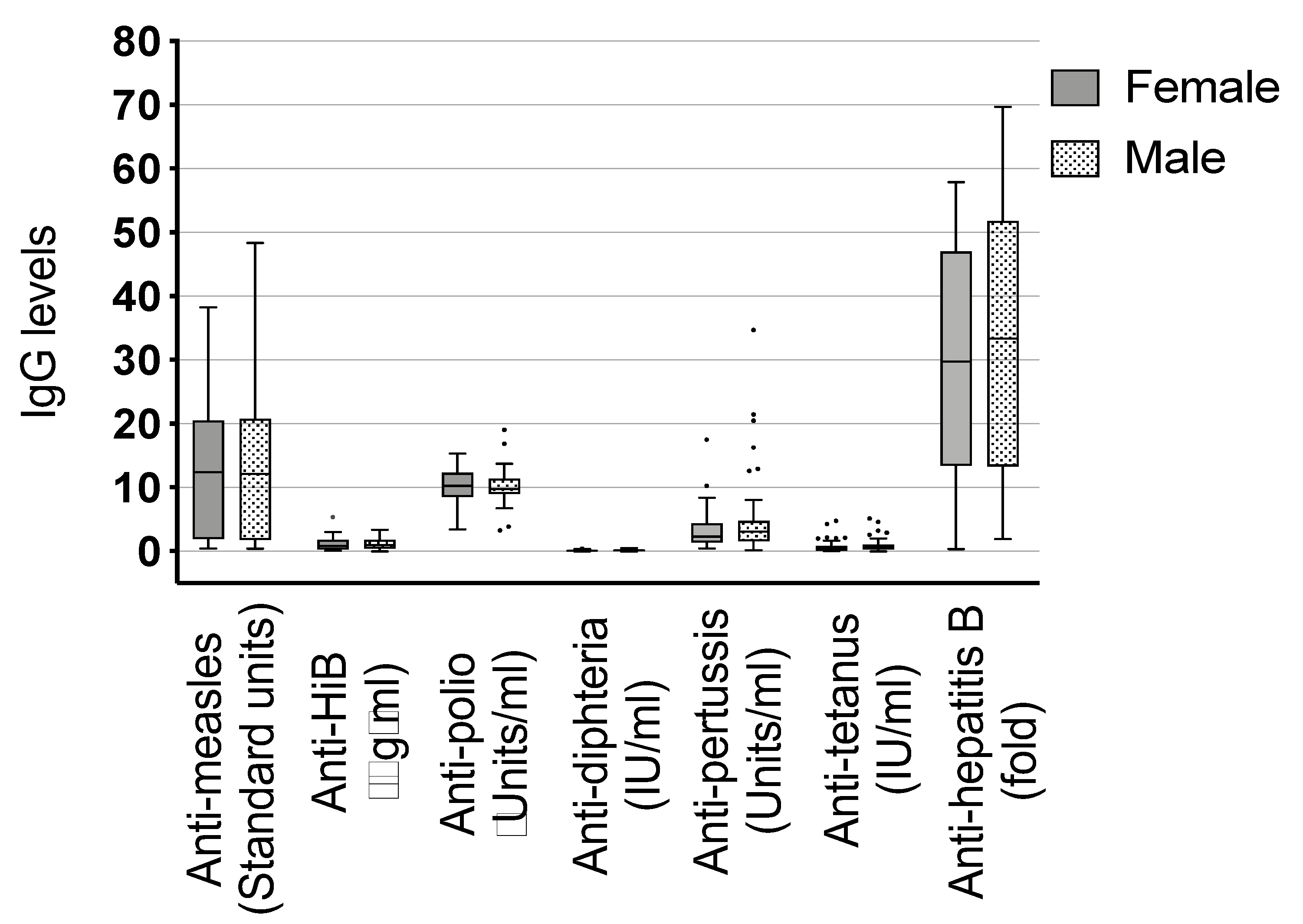
3.3. Additional Considerations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Colditz, G.A.; Brewer, T.F.; Berkey, C.S.; Wilson, M.E.; Burdick, E.; Fineberg, H.V.; Mosteller, F. Efficacy of BCG vaccine in the prevention of tuberculosis. Meta-analysis of the published literature. JAMA 1994, 271, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Patriarca, P.A.; Wright, P.F.; John, T.J. Factors affecting the immunogenicity of oral poliovirus vaccine in developing countries: Review. Rev. Infect. Dis. 1991, 13, 926–939. [Google Scholar] [CrossRef] [PubMed]
- Yazdanbakhsh, M.; Luty, A.J. Wormy mothers, healthy babies: Case closed or conundrum? Lancet 2011, 377, 6–8. [Google Scholar] [CrossRef]
- Elias, D.; Wolday, D.; Akuffo, H.; Petros, B.; Bronner, U.; Britton, S. Effect of deworming on human T cell responses to mycobacterial antigens in helminth-exposed individuals before and after bacille Calmette-Guerin (BCG) vaccination. Clin. Exp. Immunol. 2001, 123, 219–225. [Google Scholar] [CrossRef]
- Gruber, J.F.; Hille, D.A.; Liu, G.F.; Kaplan, S.S.; Nelson, M.; Goveia, M.G.; Mast, T.C. Heterogeneity of Rotavirus Vaccine Efficacy Among Infants in Developing Countries. Pediatr. Infect. Dis. J. 2017, 36, 72–78. [Google Scholar] [CrossRef]
- Levine, M.M. Immunogenicity and efficacy of oral vaccines in developing countries: Lessons from a live cholera vaccine. BMC Biol. 2010, 8, 129. [Google Scholar] [CrossRef]
- Parker, E.P.; Ramani, S.; Lopman, B.A.; Church, J.A.; Iturriza-Gomara, M.; Prendergast, A.J.; Grassly, N.C. Causes of impaired oral vaccine efficacy in developing countries. Future Microbiol. 2018, 13, 97–118. [Google Scholar] [CrossRef]
- van Riet, E.; Adegnika, A.A.; Retra, K.; Vieira, R.; Tielens, A.G.; Lell, B.; Issifou, S.; Hartgers, F.C.; Rimmelzwaan, G.F.; Kremsner, P.G.; et al. Cellular and humoral responses to influenza in gabonese children living in rural and semi-urban areas. J. Infect. Dis. 2007, 196, 1671–1678. [Google Scholar] [CrossRef]
- Casapia, M.; Joseph, S.A.; Nunez, C.; Rahme, E.; Gyorkos, T.W. Parasite and maternal risk factors for malnutrition in preschool-age children in Belen, Peru using the new WHO Child Growth Standards. Br. J. Nutr. 2007, 98, 1259–1266. [Google Scholar] [CrossRef][Green Version]
- Lindsey, B.B.; Armitage, E.P.; Kampmann, B.; de Silva, T.I. The efficacy, effectiveness, and immunogenicity of influenza vaccines in Africa: A systematic review. Lancet Infect. Dis. 2019, 19, e110–e119. [Google Scholar] [CrossRef]
- de Souza Fernandes, E.; Correia do Nascimento, W.R.; Alves dos Santos, P.; de Oliveira Nóbrega, C.G.; Barros Lorena, V.; Simões Barbosa, C.; Medeiros, D.; Wanick Sarinho, S.; Assis Costa, V.M.; Oliveira de Souza, V.M. Immune response to purified protein derivative in infants from helminth-sensitized mothers–A cases series. J. Microbiol. Immunol. Infect. 2018, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Lule, S.A.; Mawa, P.A.; Nkurunungi, G.; Nampijja, M.; Kizito, D.; Akello, F.; Muhangi, L.; Elliott, A.M.; Webb, E.L. Factors associated with tuberculosis infection, and with anti-mycobacterial immune responses, among five year olds BCG-immunised at birth in Entebbe, Uganda. Vaccine 2015, 33, 796–804. [Google Scholar] [CrossRef]
- Bethony, J.; Brooker, S.; Albonico, M.; Geiger, S.M.; Loukas, A.; Diemert, D.; Hotez, P.J. Soil-transmitted helminth infections: Ascariasis, trichuriasis, and hookworm. Lancet 2006, 367, 1521–1532. [Google Scholar] [CrossRef]
- Hamid, F.; Amoah, A.S.; van Ree, R.; Yazdanbakhsh, M. Helminth-induced IgE and protection against allergic disorders. Curr. Top. Microbiol. Immunol. 2015, 388, 91–108. [Google Scholar]
- LaBeaud, A.D.; Nayakwadi Singer, M.; McKibben, M.; Mungai, P.; Muchiri, E.M.; McKibben, E.; Gildengorin, G.; Sutherland, L.J.; King, C.H.; King, C.L.; et al. Parasitism in Children Aged Three Years and Under: Relationship between Infection and Growth in Rural Coastal Kenya. PLoS Negl. Trop. Dis. 2015, 9, e0003721. [Google Scholar] [CrossRef] [PubMed]
- Foy, H.; Nelson, G.S. Helminths in the Etiology of Anemia in the Tropics, with Special Reference to Hookworms and Schistosomes. Exp. Parasitol. 1963, 14, 240–262. [Google Scholar] [CrossRef]
- Gyorkos, T.W.; Gilbert, N.L.; Larocque, R.; Casapia, M. Trichuris and hookworm infections associated with anaemia during pregnancy. Trop. Med. Int. Health 2011, 16, 531–537. [Google Scholar] [CrossRef]
- Jardim-Botelho, A.; Raff, S.; Rodrigues Rde, A.; Hoffman, H.J.; Diemert, D.J.; Correa-Oliveira, R.; Bethony, J.M.; Gazzinelli, M.F. Hookworm, Ascaris lumbricoides infection and polyparasitism associated with poor cognitive performance in Brazilian schoolchildren. Trop. Med. Int. Health 2008, 13, 994–1004. [Google Scholar] [CrossRef]
- Lobato, L.; Miranda, A.; Faria, I.M.; Bethony, J.M.; Gazzinelli, M.F. Development of cognitive abilities of children infected with helminths through health education. Rev. Soc. Bras Med. Trop. 2012, 45, 514–519. [Google Scholar] [CrossRef]
- Mascarini-Serra, L. Prevention of Soil-transmitted Helminth Infection. J. Glob. Infect. Dis. 2011, 3, 175–182. [Google Scholar] [CrossRef]
- Mintsa Nguema, R.; Mavoungou, J.F.; Me Ngou-Milama, K.M.; Mamfoumbi, M.M.; Koumba, A.A.; Lamine, M.S.; Diarra, A.; Asseko, G.N.; Mourou, J.R.; Bouyou Akotet, M.K.; et al. Baseline Mapping of Schistosomiasis and Soil Transmitted Helminthiasis in the Northern and Eastern Health Regions of Gabon, Central Africa: Recommendations for Preventive Chemotherapy. Trop. Med. Infect. Dis. 2018, 3, 119. [Google Scholar] [CrossRef] [PubMed]
- Veletzky, L.; Hergeth, J.; Stelzl, D.R.; Mischlinger, J.; Manego, R.Z.; Mombo-Ngoma, G.; McCall, M.B.B.; Adegnika, A.A.; Agnandji, S.T.; Metzger, W.G.; et al. Burden of disease in Gabon caused by loiasis: A cross-sectional survey. Lancet Infect. Dis. 2020, S1473-3099, 30256–30265. [Google Scholar]
- Adegnika, A.A.; Ramharter, M.; Agnandji, S.T.; Ateba Ngoa, U.; Issifou, S.; Yazdanbahksh, M.; Kremsner, P.G. Epidemiology of parasitic co-infections during pregnancy in Lambarene, Gabon. Trop. Med. Int. Health 2010, 15, 1204–1209. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, E.; Harder, J.; Lacorcia, M.; Honkpehedji, Y.J.; Nouatin, O.P.; van Dam, G.J.; Corstjens, P.; Sartono, E.; Esen, M.; Lobmaier, S.M.; et al. Placental gene expression and antibody levels of mother-neonate pairs reveal an enhanced risk for inflammation in a helminth endemic country. Sci. Rep. 2019, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Dejon-Agobe, J.C.; Honkpehedji, Y.J.; Zinsou, J.F.; Edoa, J.R.; Adégbitè, B.R.; Mangaboula, A.; Agnandji, S.T.; Mombo-Ngoma, G.; Ramharter, M.; Kremsner, P.G.; et al. Epidemiology of Schistosomiasis and Soil-Transmitted Helminth Coinfections among Schoolchildren Living in Lambarene, Gabon. Am. J. Trop. Med. Hyg. 2020, 19, 0835. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, C.A.; Barreto, M.L.; Rodrigues, L.C.; Cooper, P.J.; Silva, N.B.; Amorim, L.D.; Alcantara-Neves, N.M. Chronic intestinal helminth infections are associated with immune hyporesponsiveness and induction of a regulatory network. Infect Immun. 2010, 78, 3160–3167. [Google Scholar] [CrossRef]
- Gazzinelli-Guimaraes, P.H.; Nutman, T.B. Helminth parasites and immune regulation. F1000Res 2018, 7, 15596. [Google Scholar] [CrossRef]
- Dauby, N.; Goetghebuer, T.; Kollmann, T.R.; Levy, J.; Marchant, A. Uninfected but not unaffected: Chronic maternal infections during pregnancy, fetal immunity, and susceptibility to postnatal infections. Lancet Infect. Dis. 2012, 12, 330–340. [Google Scholar] [CrossRef]
- Araujo, M.I.; Lopes, A.A.; Medeiros, M.; Cruz, A.A.; Sousa-Atta, L.; Sole, D.; Carvalho, E.M. Inverse association between skin response to aeroallergens and Schistosoma mansoni infection. Int. Arch. Allergy Immunol. 2000, 123, 145–148. [Google Scholar] [CrossRef]
- Logan, J.; Navarro, S.; Loukas, A.; Giacomin, P. Helminth-induced regulatory T cells and suppression of allergic responses. Curr. Opin. Immunol. 2018, 54, 1–6. [Google Scholar] [CrossRef]
- Lynch, N.R.; Hagel, I.; Perez, M.; Di Prisco, M.C.; Lopez, R.; Alvarez, N. Effect of anthelmintic treatment on the allergic reactivity of children in a tropical slum. J. Allergy Clin. Immunol. 1993, 92, 404–411. [Google Scholar] [CrossRef]
- Maizels, R.M.; Balic, A.; Gomez-Escobar, N.; Nair, M.; Taylor, M.D.; Allen, J.E. Helminth parasites—Masters of regulation. Immunol. Rev. 2004, 201, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, J.V.; Elliott, D.E. Helminth infections decrease host susceptibility to immune-mediated diseases. J. Immunol. 2014, 193, 3239–3247. [Google Scholar] [CrossRef]
- Kamal, S.M.; Khalifa, K.E. Immune modulation by helminthic infections: Worms and viral infections. Parasite Immunol. 2006, 28, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, W.; Brunn, M.L.; Stetter, N.; Gagliani, N.; Muscate, F.; Stanelle-Bertram, S.; Gabriel, G.; Breloer, M. Helminth Infections Suppress the Efficacy of Vaccination against Seasonal Influenza. Cell Rep. 2019, 29, 2243–2256.e4. [Google Scholar] [CrossRef] [PubMed]
- Esen, M.; Mordmuller, B.; de Salazar, P.M.; Adegnika, A.A.; Agnandji, S.T.; Schaumburg, F.; Hounkpatin, A.B.; Bruckner, S.; Theisen, M.; Belard, S.; et al. Reduced antibody responses against Plasmodium falciparum vaccine candidate antigens in the presence of Trichuris trichiura. Vaccine 2012, 30, 7621–7624. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.J.; Espinel, I.; Wieseman, M.; Paredes, W.; Espinel, M.; Guderian, R.H.; Nutman, T.B. Human onchocerciasis and tetanus vaccination: Impact on the postvaccination antitetanus antibody response. Infect. Immun. 1999, 67, 5951–5957. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.J.; Chico, M.E.; Losonsky, G.; Sandoval, C.; Espinel, I.; Sridhara, R.; Aguilar, M.; Guevara, A.; Guderian, R.H.; Levine, M.M.; et al. Albendazole treatment of children with ascariasis enhances the vibriocidal antibody response to the live attenuated oral cholera vaccine CVD 103-HgR. J. Infect. Dis. 2000, 182, 1199–1206. [Google Scholar] [CrossRef]
- Bruckner, S.; Agnandji, S.T.; Berberich, S.; Bache, E.; Fernandes, J.F.; Schweiger, B.; Massinga Loembe, M.; Engleitner, T.; Lell, B.; Mordmuller, B.; et al. Effect of Antihelminthic Treatment on Vaccine Immunogenicity to a Seasonal Influenza Vaccine in Primary School Children in Gabon: A Randomized Placebo-Controlled Trial. PLoS Negl. Trop. Dis. 2015, 9, e0003768. [Google Scholar] [CrossRef]
- Bruckner, S.; Agnandji, S.T.; Elias, J.; Berberich, S.; Bache, E.; Fernandes, J.; Loembe, M.M.; Hass, J.; Lell, B.; Mordmuller, B.; et al. A single-dose antihelminthic treatment does not influence immunogenicity of a meningococcal and a cholera vaccine in Gabonese school children. Vaccine 2016, 34, 5384–5390. [Google Scholar] [CrossRef]
- Labeaud, A.D.; Malhotra, I.; King, M.J.; King, C.L.; King, C.H. Do antenatal parasite infections devalue childhood vaccination? PLoS Negl. Trop. Dis. 2009, 3, e442. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, I.; Ouma, J.; Wamachi, A.; Kioko, J.; Mungai, P.; Omollo, A.; Elson, L.; Koech, D.; Kazura, J.W.; King, C.L. In utero exposure to helminth and mycobacterial antigens generates cytokine responses similar to that observed in adults. J. Clin. Investig. 1997, 99, 1759–1766. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, I.; Mungai, P.; Wamachi, A.; Kioko, J.; Ouma, J.H.; Kazura, J.W.; King, C.L. Helminth- and Bacillus Calmette-Guerin-induced immunity in children sensitized in utero to filariasis and schistosomiasis. J. Immunol. 1999, 162, 6843–6848. [Google Scholar] [PubMed]
- Malhotra, I.; Mungai, P.L.; Wamachi, A.N.; Tisch, D.; Kioko, J.M.; Ouma, J.H.; Muchiri, E.; Kazura, J.W.; King, C.L. Prenatal T cell immunity to Wuchereria bancrofti and its effect on filarial immunity and infection susceptibility during childhood. J. Infect. Dis. 2006, 193, 1005–1013. [Google Scholar] [CrossRef]
- Nash, S.; Mentzer, A.J.; Lule, S.A.; Kizito, D.; Smits, G.; van der Klis, F.R.; Elliott, A.M. The impact of prenatal exposure to parasitic infections and to anthelminthic treatment on antibody responses to routine immunisations given in infancy: Secondary analysis of a randomised controlled trial. PLoS Negl. Trop. Dis. 2017, 11, e0005213. [Google Scholar] [CrossRef]
- Webb, E.L.; Mawa, P.A.; Ndibazza, J.; Kizito, D.; Namatovu, A.; Kyosiimire-Lugemwa, J.; Nanteza, B.; Nampijja, M.; Muhangi, L.; Woodburn, P.W.; et al. Effect of single-dose anthelmintic treatment during pregnancy on an infant’s response to immunisation and on susceptibility to infectious diseases in infancy: A randomised, double-blind, placebo-controlled trial. Lancet 2011, 377, 52–62. [Google Scholar] [CrossRef]
- Elliott, A.M.; Mawa, P.A.; Webb, E.L.; Nampijja, M.; Lyadda, N.; Bukusuba, J.; Kizza, M.; Namujju, P.B.; Nabulime, J.; Ndibazza, J.; et al. Effects of maternal and infant co-infections, and of maternal immunisation, on the infant response to BCG and tetanus immunisation. Vaccine 2010, 29, 247–255. [Google Scholar] [CrossRef]
- Ondigo, B.N.; Muok, E.M.O.; Oguso, J.K.; Njenga, S.M.; Kanyi, H.M.; Ndombi, E.M.; Priest, J.W.; Kittur, N.; Secor, W.E.; Karanja, D.M.S.; et al. Impact of Mothers’ Schistosomiasis Status During Gestation on Children’s IgG Antibody Responses to Routine Vaccines 2 Years Later and Anti-Schistosome and Anti-Malarial Responses by Neonates in Western Kenya. Front. Immunol. 2018, 9, 1402. [Google Scholar] [CrossRef]
- Katz, N.; Chaves, A.; Pellegrino, J. A simple device for quantitative stool thick-smear technique in Schistosomiasis mansoni. Rev. Inst. Med. Trop. Sao Paulo 1972, 14, 397–400. [Google Scholar]
- Mbong Ngwese, M.; Prince Manouana, G.; Nguema Moure, P.A.; Ramharter, M.; Esen, M.; Adégnika, A.A. Diagnostic Techniques of Soil-Transmitted Helminths: Impact on Control Measures. Trop. Med. Infect. Dis. 2020, 5, 93. [Google Scholar] [CrossRef]
- Feldmeier, H.; Bienzle, U.; Dietrich, M.; Sievertsen, H.J. Combination of a viability test and a quantification method for Schistosoma haematobium eggs (filtration--trypan blue-staining-technique). Tropenmed. Parasitol. 1979, 30, 417–422. [Google Scholar] [PubMed]
- Noireau, F.; Apembet, J.D. Comparison of thick blood smear and saponin haemolysis for the detection of Loa loa and Mansonella perstans infections. J. Trop. Med. Hyg. 1990, 93, 390–392. [Google Scholar] [PubMed]
- Borkow, G.; Bentwich, Z. Chronic parasite infections cause immune changes that could affect successful vaccination. Trends Parasitol. 2008, 24, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Kabagenyi, J.; Natukunda, A.; Nassuuna, J.; Sanya, R.E.; Nampijja, M.; Webb, E.L.; Elliott, A.M.; Nkurunungi, G. Urban-rural differences in immune responses to mycobacterial and tetanus vaccine antigens in a tropical setting: A role for helminths? Parasitol. Int. 2020, 78, 102132. [Google Scholar] [CrossRef]
- Wajja, A.; Kizito, D.; Nassanga, B.; Nalwoga, A.; Kabagenyi, J.; Kimuda, S.; Galiwango, R.; Mutonyi, G.; Vermaak, S.; Satti, I.; et al. The effect of current Schistosoma mansoni infection on the immunogenicity of a candidate TB vaccine, MVA85A, in BCG-vaccinated adolescents: An open-label trial. PLoS Negl. Trop. Dis. 2017, 11, e0005440. [Google Scholar] [CrossRef] [PubMed]
- Riner, D.K.; Ndombi, E.M.; Carter, J.M.; Omondi, A.; Kittur, N.; Kavere, E.; Korir, H.K.; Flaherty, B.; Karanja, D.; Colley, D.G. Schistosoma mansoni Infection Can Jeopardize the Duration of Protective Levels of Antibody Responses to Immunizations against Hepatitis B and Tetanus Toxoid. PLoS Negl. Trop. Dis. 2016, 10, e0005180. [Google Scholar] [CrossRef]
- Adegnika, A.A.; Zinsou, J.F.; Issifou, S.; Ateba-Ngoa, U.; Kassa, R.F.; Feugap, E.N.; Honkpehedji, Y.J.; Dejon Agobe, J.C.; Kenguele, H.M.; Massinga-Loembe, M.; et al. Randomized, controlled, assessor-blind clinical trial to assess the efficacy of single- versus repeated-dose albendazole to treat ascaris lumbricoides, trichuris trichiura, and hookworm infection. Antimicrob. Agents Chemother. 2014, 58, 2535–2540. [Google Scholar] [CrossRef]
- Vouking, M.Z.; Tadenfok, C.N.; Ekani, J.M.E. Strategies to increase immunization coverage of tetanus vaccine among women in Sub Saharan Africa: A systematic review. Pan. Afr. Med. J. 2017, 27 (Suppl. 3), 25. [Google Scholar] [CrossRef]
- Ahmad, S.M.; Alam, J.; Afsar, N.A.; Huda, N.; Kabir, Y.; Qadri, F.; Raqib, R.; Stephensen, C.B. Comparisons of the effect of naturally acquired maternal pertussis antibodies and antenatal vaccination induced maternal tetanus antibodies on infant’s antibody secreting lymphocyte responses and circulating plasma antibody levels. Hum. Vaccines Immunother. 2016, 12, 886–893. [Google Scholar] [CrossRef]
- Munoz, F.M.; Bond, N.H.; Maccato, M.; Pinell, P.; Hammill, H.A.; Swamy, G.K.; Walter, E.B.; Jackson, L.A.; Englund, J.A.; Edwards, M.S.; et al. Safety and immunogenicity of tetanus diphtheria and acellular pertussis (Tdap) immunization during pregnancy in mothers and infants: A randomized clinical trial. JAMA 2014, 311, 1760–1769. [Google Scholar] [CrossRef]
- Maertens, K.; Cabore, R.N.; Huygen, K.; Hens, N.; Van Damme, P.; Leuridan, E. Pertussis vaccination during pregnancy in Belgium: Results of a prospective controlled cohort study. Vaccine 2016, 34, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Ladhani, S.N.; Andrews, N.J.; Southern, J.; Jones, C.E.; Amirthalingam, G.; Waight, P.A.; England, A.; Matheson, M.; Bai, X.; Findlow, H.; et al. Antibody responses after primary immunization in infants born to women receiving a pertussis-containing vaccine during pregnancy: Single arm observational study with a historical comparator. Clin. Infect Dis. 2015, 61, 1637–1644. [Google Scholar] [CrossRef] [PubMed]
- Amirthalingam, G.; Campbell, H.; Ribeiro, S.; Fry, N.K.; Ramsay, M.; Miller, E.; Andrews, N. Sustained Effectiveness of the Maternal Pertussis Immunization Program in England 3 Years Following Introduction. Clin. Infect. Dis. 2016, 63 (Suppl. 4), S236–S243. [Google Scholar] [CrossRef]
- Bellido-Blasco, J.; Guiral-Rodrigo, S.; Miguez-Santiyan, A.; Salazar-Cifre, A.; Gonzalez-Moran, F. A case-control study to assess the effectiveness of pertussis vaccination during pregnancy on newborns, Valencian community, Spain, 1 March 2015 to 29 February 2016. Euro. Surveill 2017, 22, 30545. [Google Scholar] [CrossRef] [PubMed]
- Saul, N.; Wang, K.; Bag, S.; Baldwin, H.; Alexander, K.; Chandra, M.; Thomas, J.; Quinn, H.; Sheppeard, V.; Conaty, S. Effectiveness of maternal pertussis vaccination in preventing infection and disease in infants: The NSW Public Health Network case-control study. Vaccine 2018, 36, 1887–1892. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Marriott, I.; Fish, E.N. Sex-based differences in immune function and responses to vaccination. Trans. R. Soc. Trop Med. Hyg. 2015, 109, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, K.L.; Fink, A.L.; Plebanski, M.; Klein, S.L. Sex and Gender Differences in the Outcomes of Vaccination over the Life Course. Annu. Rev. Cell Dev. Biol. 2017, 33, 577–599. [Google Scholar] [CrossRef]
- Klein, S.L.; Jedlicka, A.; Pekosz, A. The Xs and Y of immune responses to viral vaccines. Lancet Infect. Dis. 2010, 10, 338–349. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Overall | Uninfected | Infected |
|---|---|---|---|
| Mother (n) | 123 | 71 | 52 |
| Median age in years (range) | 24 (13–43) | 26 (13–42) | 23 (14–43) |
| Median weight in kg (range) | 63 (37–153) | 65 (45–153) | 63 (37–100) |
| Median height in cm (range) | 160 (141–171) | 160 (147–171) | 159 (141–170) |
| Parity | |||
| 1st parity in % (n) | 22.8 (28) | 18.3 (13) | 28.8 (15) |
| 1 previous parity in % (n) | 18.7 (23) | 18.3 (13) | 19.2 (10) |
| 2-4 previous parities in % (n) | 35 (43) | 38 (27) | 30.8 (16) |
| 5 or more previous parities in % (n) | 17 (21) | 21.1 (15) | 11.6 (6) |
| NA in % (n) | 6.5 (8) | 4.3 (3) | 9.6 (5) |
| Infant (n) | 124 | 71 | 53 |
| Median birth weight in kg (range) | 3.1 (1.7–4.3) | 3.1 (1.7–4.1) | 3.1 (2.1–4.3) |
| Median height in cm (range) | 50 (43–57) | 50 (44–56) | 50 (43–57) |
| Sex * | |||
| Male (n) | 61 | 33 | 28 |
| Female (n) | 59 | 35 | 24 |
| Breastfeeding | |||
| Yes (n) | 48 | 25 | 23 |
| No (n) | 52 | 32 | 20 |
| NA (n) | 24 | 14 | 10 |
| Mother | Cord Blood | |||||||||||
| uninfected | infected | single infection | multiple infection | Trichuris sp. | Schistosoma sp. | uninfected | infected | single infection | multiple infection | Trichuris sp. | Schistosoma sp. | |
| Polio | 8.74 (6.86–10.48) | 9.01 (6.53–11.61) | 9.01 (6.50–11.67) | 8.35 (6.53–10.03) | 9.69 (9.69–12.11) | 8.69 (6.02–10.08) | 8.61 (6.88–10.59) | 9.11 (6.57–11.06) | 9.09 (5.56–10.93) | 9.41 (6.31–11.91) | 9 (6.06–9.89) | 8.40 (6.12–10.34) |
| Diphtheria | 0.02 (0.01–0.03) | 0.02 (0.01–0.04) | 0.02 (0.01–0.05) | 0.0 6(0.01–0.11) | 0.03 (0.01–0.06) | 0.02 (0.01–0.06) | 0.02 (0.01–0.03) | 0.02 (0.01–0.03) | 0.02 (0.004–0.04) | 0.03 (0.01–0.14) | 0.02 (0.01–0.05) | 0.01 (0.006–0.04) |
| Hepatitis B | 0.24 (0.09–0.57) | 0.32 (0.15–1.07 | 1.07 (0.19–24.40) | 0. 66(0.15–6.173) | 0.31 (0.15–24.40) | 0.47 (0.15–2.37) | 0.18 (0.11–0.44) | 0.24 (0.10–0.44) | 0.50 (0.22–26.05) | 0.27 (0.11–6.13) | 0.27 (0.20–12.75) | 0.33 (0.13–2.09) |
| HiB | 0.37 (0.16–0.63) | 0.31 (0.16–0.43) | 0.45 (0.34–0.79) | 0.83 (0.40–1.24) | 0.55 (0.35–1.25) | 0.79 (0.38–1.90) | 0.39 (0.17–0.59) | 0.3 6(0–0.49) | 0.48 (0.34–0.90) | 0.78 (0.38–1.16) | 0.92 (0.45–1.61) | 0.84 (0.40–1.24) |
| Pertussis | 11 (7.68–18.68) | 9.40 (6.08–14.74) | 8.82 (4.24–14.68) | 11.82 (8.81–15.95) | 11.82 (5.77–14.67) | 10.1 6(5.32–14.93) | 9.64 (6.43–14.16) | 11.54 (5.97–14.33) | 11.40 (5.40–12.58) | 13.0 6(8.13–23.28) | 11.53 (7.89–13.10) | 11.69 (6.24–18.97) |
| Tetanus | 3.37 (1.76–5.14) | 2.31 (1.17–4.91) | 2.72 (1.42–4.84) | 1.75 (0.85–5.09) | 2.74 (1.52–5.29) | 1.68 (0.75–3.36) | 3.48 (1.90–5.64) | 2.38 (0.96–4.59) | 2.74 (1.18–4.90) | 1.91 (0.56–4.15) | 3.05 (1.59–5.32) | 1.79 (0.98–3.63) |
| Measles | 22.39 (12.21–33.45) | 22.14 (12.14–31.93) | 22.14 (12.72–31.95) | 14.04 (9.5–30.13) | 25.15 (13.26–30.13) | 17.22 (10.58–30.59) | 24.20 (13.45–38.12) | 20.94 (11.50–30.57) | 23.99 (14.6–33.05) | 16.22 (9.98–25.96) | 23.78 (14.52–34.01) | 16.65 (10.55–34.33) |
| Child 9 MO | Child 12 MO | |||||||||||
| uninfected | infected | single infection | multiple infection | Trichuris sp. | Schistosoma sp. | uninfected | infected | single infection | multiple infection | Trichuris sp. | Schistosoma sp. | |
| Polio | 9.01 (7.53–10.76) | 9 (7.07–10.63) | 8.83 (6.58–10.32) | 9.19 (7.48–10.80) | 9.38 (7.42–10.80) | 9.2 (5.40–11.34) | 9.04 (7.54–10.82) | 9.83 (8.87–12.25) | 9.69 (8.22–11.58) | 9.97 (8.82–12.25) | 9.83 (8.46–12.36) | 9.69 (8.09–12.33) |
| Diphtheria | 0.17 (0.05–0.31) | 0.1 6(0.03–0.27) | 0.17 (0.03–0.37) | 0.05 (0.02–0.40) | 0.05 (0.01–0.42) | 0.14 (0.03–0.36) | 0.13 (0.31–0.27) | 0.08 (0.02–0.18) | 0.11 (0.02–0.19) | 0.08 (0.02–0.49) | 0.1 6(0.03–0.38) | 0.09 (0.04–0.34) |
| Hepatitis B | 46.41 (25.75–54.19) | 40.54 (13.15–54.86) | 43.95 (21.48–55.35) | 28. 66(9.85–55.33) | 22.82 (5.02–54.96) | 38.91 (12–53.17) | 33.43 (16.43–46.21) | 20.15 (13.18–49.28) | 35.31 (7.35–48.77) | 17.28 (10.78–49.65) | 16.14 (5.99–49.65) | 19.32 (5.95–45.27) |
| HiB | 0.43 (0.11–1.31) | 0.44 (0.19–0.93) | 0.91 (0.42–2.64) | 1.50 (0.41–4.93) | 0.90 (0.29–4.02) | 0.57 (0.37–4.98) | 0.35 (0–0.88) | 0.51 (0.20–1.20) | 1.28 (0.67–7.99) | 1.23 (0.24–6.45) | 1. 66(0.49–8.23) | 1.68 (0.31–8.67) |
| Pertussis | 1.99 (1.37–3.16) | 2.35 (1.33–3.81) | 2.14 (1.27–3.06) | 2.97 (2.22–5.84) | 2.5 6(1.20–5.31) | 2.49 (1.71–4.77) | 2.50 (1.41–4.12) | 2.31 (1.57–3.97) | 1.98 (1.27–3.37) | 4.69 (2.45–7.60) | 3.67 (1.71–5.99) | 2.49 (1.80–6.58) |
| Tetanus | 0.54 (0.16–0.92) | 0.54 (0.18–1.30) | 0.54 (0.15–1.27) | 0.44 (0.17–1.28) | 0.48 (0.15–1.37) | 0.51 (0.17–1.28) | 0.39 (0.10–0.815) | 0.47 (0.11–0.85) | 0.62 (0.33–1.02) | 0.44 (0.09–2.16) | 0.65 (0.14–1.09) | 0.45 (0.15–2.06) |
| Measles | 1.60 (0.69–2.18) | 1.4 6(0.71–2.45) | 1.44 (0.86–2.08) | 1.04 (0.54–2.80) | 1.64 (0.67–2.23) | 1.10 (0.52–2.37) | 16.75 (1.69–23.25) | 8.89 (1.93–16.38) | 11.75 (4.16–18.77) | 4.98 (0.68–14.28) | 1.64 (0.67–2.23) | 7.28 (4.51–12.33) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flügge, J.; Adegnika, A.A.; Honkpehedji, Y.J.; Sandri, T.L.; Askani, E.; Manouana, G.P.; Massinga Loembe, M.; Brückner, S.; Duali, M.; Strunk, J.; et al. Impact of Helminth Infections during Pregnancy on Vaccine Immunogenicity in Gabonese Infants. Vaccines 2020, 8, 381. https://doi.org/10.3390/vaccines8030381
Flügge J, Adegnika AA, Honkpehedji YJ, Sandri TL, Askani E, Manouana GP, Massinga Loembe M, Brückner S, Duali M, Strunk J, et al. Impact of Helminth Infections during Pregnancy on Vaccine Immunogenicity in Gabonese Infants. Vaccines. 2020; 8(3):381. https://doi.org/10.3390/vaccines8030381
Chicago/Turabian StyleFlügge, Judith, Ayôla Akim Adegnika, Yabo Josiane Honkpehedji, Thaisa L. Sandri, Esther Askani, Gédéon Prince Manouana, Marguerite Massinga Loembe, Sina Brückner, Mohamed Duali, Johannes Strunk, and et al. 2020. "Impact of Helminth Infections during Pregnancy on Vaccine Immunogenicity in Gabonese Infants" Vaccines 8, no. 3: 381. https://doi.org/10.3390/vaccines8030381
APA StyleFlügge, J., Adegnika, A. A., Honkpehedji, Y. J., Sandri, T. L., Askani, E., Manouana, G. P., Massinga Loembe, M., Brückner, S., Duali, M., Strunk, J., Mordmüller, B., Agnandji, S. T., Lell, B., Kremsner, P. G., & Esen, M. (2020). Impact of Helminth Infections during Pregnancy on Vaccine Immunogenicity in Gabonese Infants. Vaccines, 8(3), 381. https://doi.org/10.3390/vaccines8030381