What Does Plant-Based Vaccine Technology Offer to the Fight against COVID-19?



 ,
,
Abstract
1. Introduction
2. Development of Therapeutic and Prophylactic Treatments against COVID-19
3. Molecular Farming and the Plant-based Vaccines Technology
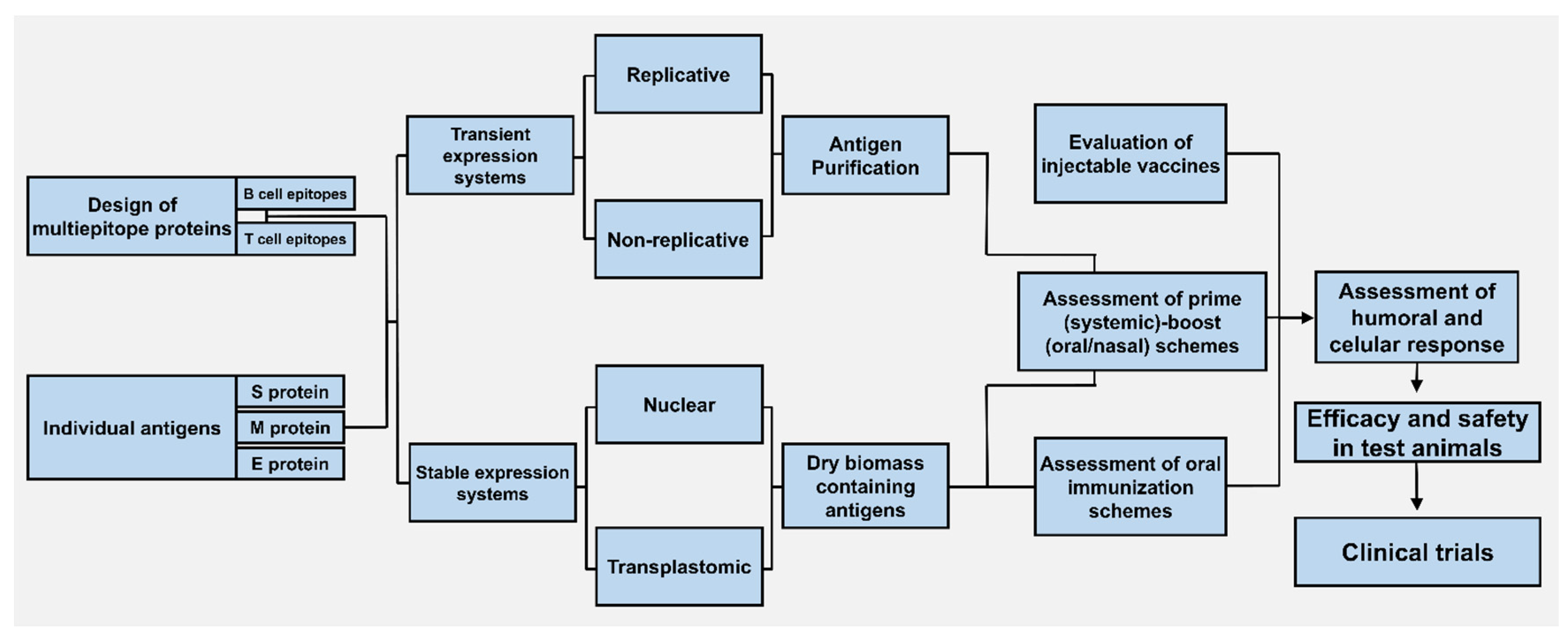
3.1. Approaches for Recombinant Protein Expression in Plants
3.2. Current Scenario of Plant-Based Vaccines
4. Possibilities to Develop Anti-COVID-19 Plant-based Vaccines
4.1. Virus-Like Particles (VLPs)
4.2. Multiepitopic Vaccines
4.3. Immune Complexes
4.4. Elastin-Like Polypeptide Fusions
5. Efforts to Develop Plant-Based Vaccines against MERS and SARS-CoV-1
6. Relevance of Mucosal Vaccines and Prime-Boosting Immunization Schemes
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO (World Health Organization). Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 25 March 2020).
- Chen, Y.; Liu, Q.; Guo, D. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef]
- Tian, X.; Li, C.; Huang, A.; Xia, S.; Lu, S.; Shi, Z.; Lu, L.; Jiang, S.; Yang, Z.; Wu, Y.; et al. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg. Microbes Infect. 2020, 9, 382–385. [Google Scholar] [CrossRef]
- Wu, A.; Peng, Y.; Huang, B.; Ding, X.; Wang, X.; Niu, P.; Meng, J.; Zhu, Z.; Zhang, Z.; Wang, J.; et al. Genome composition and divergence of the novel coronavirus (2019-nCoV) originating in China. Cell Host Microbe 2020, 27, 325–328. [Google Scholar] [CrossRef]
- Rothan, H.A.; Byrareddy, S.N. The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J. Autoimmun. 2020, 26, 102433. [Google Scholar] [CrossRef] [PubMed]
- Coronavirus COVID-19 Global Cases by the Center for Systems Science and Engineering (CSSE) at Johns Hopkins University (JHU). Available online: https://gisanddata.maps.arcgis.com/apps/opsdashboard/index.html#/bda7594740fd40299423467b48e9ecf6 (accessed on 25 March 2020).
- Deng, S.Q.; Peng, H.J. Characteristics of and public health responses to the coronavirus disease 2019 outbreak in China. J. Clin. Med. 2020, 9, 575. [Google Scholar] [CrossRef]
- Peng, X.; Xu, X.; Li, Y.; Cheng, L.; Zhou, X.; Ren, B. Transmission routes of 2019-nCoV and controls in dental practice. Int. J. Oral. Sci. 2020, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Torok, M.E. Taking the right measures to control COVID-19. Lancet Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Mitjà, O.; Clotet, B. Use of antiviral drugs to reduce COVID-19 transmission. Lancet Glob. Health 2020. [Google Scholar] [CrossRef]
- Gurwitz, D. Angiotensin receptor blockers as tentative SARS-CoV-2 therapeutics. Drug Dev. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Wang, Z.; Zhao, F.; Yang, Y.; Li, J.; Yuan, J.; Wang, F.; Li, D.; Yang, M.; Xing, L.; et al. Treatment of 5 critically Ill patients with COVID-19 with convalescent plasma. JAMA 2020. [Google Scholar] [CrossRef]
- Menachery, V.D.; Gralinski, L.E.; Mitchell, H.D.; Dinnon, K.H., III; Leist, S.R.; Yount, B.L., Jr.; McAnarney, E.T.; Graham, R.L.; Waters, K.M.; Baric, R.S. Combination attenuation offers strategy for live attenuated coronavirus vaccines. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Kotomina, T.; Isakova-Sivak, I.; Matyushenko, V.; Kim, K.H.; Lee, Y.; Jung, Y.J.; Kang, S.M.; Rudenko, L. Recombinant live attenuated influenza vaccine viruses carrying CD8 T-cell epitopes of respiratory syncytial virus protect mice against both pathogens without inflammatory disease. Antivir. Res. 2019, 168, 9–17. [Google Scholar] [CrossRef]
- Kobinger, G.P.; Figueredo, J.M.; Rowe, T.; Zhi, Y.; Gao, G.; Sanmiguel, J.C.; Bell, P.; Wivel, N.A.; Zitzow, L.A.; Flieder, D.B.; et al. Adenovirus-based vaccine prevents pneumonia in ferrets challenged with the SARS coronavirus and stimulates robust immune responses in macaques. Vaccine 2007, 25, 5220–5231. [Google Scholar] [CrossRef]
- Takano, T.; Yamada, S.; Doki, T.; Hohdatsu, T. Pathogenesis of oral type I feline infectious peritonitis virus (FIPV) infection: Antibody-dependent enhancement infection of cats with type I FIPV via the oral route. J. Vet. Med. Sci. 2019, 81, 911–915. [Google Scholar] [CrossRef]
- Jiang, S. Don’t rush to deploy COVID-19 vaccines and drugs without sufficient safety guarantees. Nature 2020, 579, 321. [Google Scholar] [CrossRef]
- Wu, T.; Guan, J.; Handel, A.; Tscharke, D.C.; Sidney, J.; Sette, A.; Wakim, L.M.; Sng, X.Y.X.; Thomas, P.G.; Croft, N.P.; et al. Quantification of epitope abundance reveals the effect of direct and cross-presentation on influenza CTL responses. Nat. Commun. 2019, 10, 2846. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, N.; Jasny, E.; Petsch, B. Advances in RNA vaccines for preventive indications: A case study of a vaccine against rabies. Vaccines 2019, 7, 132. [Google Scholar] [CrossRef] [PubMed]
- CureVac Focuses on the Development of mRNA-based Coronavirus Vaccine to Protect People Worldwide. Available online: https://www.curevac.com/news/curevac-focuses-on-the-development-of-mrna-based-coronavirus-vaccine-to-protect-people-worldwide (accessed on 25 March 2020).
- Safety and Immunogenicity Study of 2019-nCoV Vaccine (mRNA-1273) to Prevent SARS-CoV-2 Infection. Available online: https://clinicaltrials.gov/ct2/show/NCT04283461?term=vaccine&cond=coronavirus&draw=2&rank=3 (accessed on 25 March 2020).
- Safety and Immunity of Covid-19 aAPC Vaccine. Available online: https://clinicaltrials.gov/ct2/show/NCT04299724?term=vaccine&cond=coronavirus&draw=2&rank=6 (accessed on 25 March 2020).
- Mao, Q.Y.; Wang, Y.; Bian, L.; Xu, M.; Liang, Z. EV71 vaccine, a new tool to control outbreaks of hand, foot and mouth disease (HFMD). Expert Rev. Vaccines 2016, 15, 599–606. [Google Scholar] [CrossRef]
- Grifoni, A.; Sidney, J.; Zhang, Y.; Scheuermann, R.H.; Peters, B.; Sette, A. A sequence homology and bioinformatic approach can predict candidate targets for immune responses to SARS-CoV-2. Cell Host Microbe 2020. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 2020. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Buyel, J.F. Molecular farming-The slope of enlightenment. Biotechnol. Adv. 2020, 16, 107519. [Google Scholar] [CrossRef] [PubMed]
- Tekoah, Y.; Shulman, A.; Kizhner, T.; Ruderfer, I.; Fux, L.; Nataf, Y.; Bartfeld, D.; Ariel, T.; Gingis-Velitski, S.; Hanania, U.; et al. Large-scale production of pharmaceutical proteins in plant cell culture-the Protalix experience. Plant Biotechnol. J. 2015, 13, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Salazar, J.A.; Bañuelos, B.; Rosales, S. Current status of viral expression systems in plants and perspectives for oral vaccines development. Plant Mol. Biol. 2015, 87, 203–217. [Google Scholar] [CrossRef] [PubMed]
- García, A.L.; Rubio, N.; Moreno, L. Mucosal immunology and oral vaccination. In Genetically Engineered Plants as a Source of Vaccines Against Widespread Diseases—An Integrated View; Springer: Berlin, Germany, 2014; pp. 15–42. [Google Scholar]
- Bilichak, A.; Gaudet, D.; Laurie, J. Emerging genome engineering tools in crop research and breeding. Methods Mol. Biol. 2020, 2072, 165–181. [Google Scholar]
- Pogrebnyak, N.; Golovkin, M.; Andrianov, V.; Spitsin, S.; Smirnov, Y.; Egolf, R.; Koprowski, H. Severe acute respiratory syndrome (SARS) S protein production in plants: Development of recombinant vaccine. Proc. Natl. Acad. Sci. USA 2005, 102, 9062–9067. [Google Scholar] [CrossRef]
- Li, H.Y.; Ramalingam, S.; Chye, M.L. Accumulation of recombinant SARS-CoV spike protein in plant cytosol and chloroplasts indicate potential for development of plant-derived oral vaccines. Exp. Biol. Med. 2006, 231, 1346–1352. [Google Scholar] [CrossRef]
- Zheng, N.; Xia, R.; Yang, C.; Yin, B.; Li, Y.; Duan, C.; Liang, L.; Guo, H.; Xie, Q. Boosted expression of the SARS-CoV nucleocapsid protein in tobacco and its immunogenicity in mice. Vaccine 2009, 27, 5001–5007. [Google Scholar] [CrossRef]
- Demurtas, O.C.; Massa, S.; Illiano, E.; De Martinis, D.; Chan, P.K.; Di Bonito, P.; Franconi, R. Antigen production in plant to tackle infectious diseases flare up: The case of SARS. Front. Plant Sci. 2016, 7, 54. [Google Scholar] [CrossRef]
- Daniell, H.; Chan, H.T.; Pasoreck, E.K. Vaccination via chloroplast genetics: Affordable protein drugs for the prevention and treatment of inherited or infectious human diseases. Annu. Rev. Genet. 2016, 23, 595–618. [Google Scholar] [CrossRef]
- Olejniczak, S.A.; Łojewska, E.; Kowalczyk, T.; Sakowicz, T. Chloroplasts: State of research and practical applications of plastome sequencing. Planta 2016, 244, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Arai, Y.; Shikanai, T.; Doi, Y.; Yoshida, S.; Yamaguchi, I.; Nakashita, H. Production of polyhydroxybutyrate by polycistronic expression of bacterial genes in tobacco plastid. Plant Cell Physiol. 2004, 45, 1176–1184. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Salazar, J.A.; Monreal, E.; Herrera, S.; Koop, H.U.; Rosales, S. Plastid-based expression strategie. In Genetically Engineered Plants as a Source of Vaccines Against Wide Spread Diseases-An Integrated View; Springer: Berlin, Germany, 2014; pp. 265–280. [Google Scholar]
- Peyret, H.; Lomonossoff, G.P. When plant virology met Agrobacterium: The rise of the deconstructed clones. Plant Biotechnol. J. 2015, 13, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Govea-Alonso, D.O.; Cardineau, G.A.; Rosales-Mendoza, S. Principles of plant-based vaccines. In Genetically Engineered Plants as a Source of Vaccines Against Wide Spread Diseases—An Integrated View; Mendoza, S.R., Ed.; Springer: New York, NY, USA, 2014; ISBN 978-1-4939-0850-9. [Google Scholar]
- McNulty, M.J.; Gleba, Y.; Tusé, D.; Hahn-Löbmann, S.; Giritch, A.; Nandi, S.; McDonald, K.A. Techno-economic analysis of a plant-based platform for manufacturing antimicrobial proteins for food safety. Biotechnol. Prog. 2020, 36, 1–15. [Google Scholar] [CrossRef]
- Peyret, H.; Brown, J.K.M.; Lomonossoff, G.P. Improving plant transient expression through the rational design of synthetic 5’ and 3’ untranslated regions. Plant Methods 2019, 15, 108. [Google Scholar] [CrossRef]
- Márquez-Escobar, V.A.; Rosales-Mendoza, S.; Beltrán-López, J.I.; González-Ortega, O. Plant-based vaccines against respiratory diseases: Current status and future prospects. Expert Rev. Vaccines 2017, 16, 137–149. [Google Scholar] [CrossRef]
- Hodgins, B.; Pillet, S.; Landry, N.; Ward, B.J. Prime-pull vaccination with a plant-derived virus-like particle influenza vaccine elicits a broad immune response and protects aged mice from death and frailty after challenge. Immun. Ageing 2019, 16, 27. [Google Scholar] [CrossRef]
- Pillet, S.; Couillard, J.; Trépanier, S.; Poulin, J.F.; Yassine-Diab, B.; Guy, B.; Ward, B.J.; Landry, N. Immunogenicity and safety of a quadrivalent plant-derived virus like particle influenza vaccine candidate-Two randomized Phase II clinical trials in 18 to 49 and ≥50 years old adults. PLoS ONE 2019, 14, e0216533. [Google Scholar] [CrossRef]
- Makarkov, A.I.; Golizeh, M.; Ruiz-Lancheros, E.; Gopal, A.; Costas-Cancelas, I.N.; Chierzi, S.; Pillet, S.; Charland, N.; Landry, N.; Rouiller, I.; et al. Plant-derived virus-like particle vaccines drive cross-presentation of influenza A hemagglutinin peptides by human monocyte-derived macrophages. NPJ Vaccines 2019, 4, 17. [Google Scholar] [CrossRef]
- Hodgins, B.; Pillet, S.; Landry, N.; Ward, B.J. A plant-derived VLP influenza vaccine elicits a balanced immune response even in very old mice with co-morbidities. PLoS ONE 2019, 14, e0210009. [Google Scholar] [CrossRef]
- Won, S.Y.; Hunt, K.; Guak, H.; Hasaj, B.; Charland, N.; Landry, N.; Ward, B.J.; Krawczyk, C.M. Characterization of the innate stimulatory capacity of plant-derived virus-like particles bearing influenza hemagglutinin. Vaccine 2018, 36, 8028–8038. [Google Scholar] [CrossRef]
- Lindsay, B.J.; Bonar, M.M.; Costas-Cancelas, I.N.; Hunt, K.; Makarkov, A.I.; Chierzi, S.; Krawczyk, C.M.; Landry, N.; Ward, B.J.; Rouiller, I. Morphological characterization of a plant-made virus-like particle vaccine bearing influenza virus hemagglutinins by electron microscopy. Vaccine 2018, 36, 2147–2154. [Google Scholar] [CrossRef] [PubMed]
- Rosales, S.; Salazar, J.A. Immunological aspects of using plant cells as delivery vehicles for oral vaccines. Expert Rev. Vaccines 2014, 13, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Magnifection—A new platform for expressing recombinant vaccines in plants. Vaccine 2005, 23, 2042–2048. [Google Scholar] [CrossRef] [PubMed]
- Pniewski, T.; Milczarek, M.; Wojas-Turek, J.; Paitasz-Piasecka, E.; Wietrzyk, J.; Czyz, M. Plant lyophilisate carrying S-HBsAg as an oral booster vaccine against HBV. Vaccine 2018, 36, 6070–6076. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.F.; Chiang, C.H.; Li, Y.L.; Wang, N.M.; Song, P.P.; Lin, S.J.; Chen, Y.H. Oral edible plant vaccine containing hypoallergen of American cockroach major allergen Per a 2 prevents roach-allergic asthma in a murine model. PLoS ONE 2018, 13, 1–16. [Google Scholar] [CrossRef]
- Yusibov, V.; Streatfield, S.J.; Kushnir, N. Clinical development of plant-produced recombinant pharmaceuticals: Vaccines, antibodies and beyond. Hum. Vaccines 2011, 7, 313–321. [Google Scholar] [CrossRef]
- Rigano, M.M.; Sala, F.; Arntzen, C.J.; Walmsley, A.M. Targeting of plant-derived vaccine antigens to immunoresponsive mucosal sites. Vaccine 2003, 21, 809–811. [Google Scholar] [CrossRef]
- Rosales, S.; Nieto, R. Green therapeutic biocapsules: Using plant cells to orally deliver biopharmaceuticals. Trends Biotechnol. 2018, 36, 1054–1067. [Google Scholar] [CrossRef]
- Rybicki, E.P. Plant molecular farming of virus-like nanoparticles as vaccines and reagents. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2020, 12, e1587. [Google Scholar] [CrossRef]
- Chargelegue, D.; Drake, P.M.; Obregon, P.; Prada, A.; Fairweather, N.; Ma, J.K. Highly immunogenic and protective recombinant vaccine candidate expressed in transgenic plants. Infect. Immun. 2005, 73, 5915–5922. [Google Scholar] [CrossRef]
- Yeboah, A.; Cohen, R.I.; Rabolli, C.; Yarmush, M.L.; Berthiaume, F. Elastin-like polypeptides: A strategic fusion partner for biologics. Biotechnol. Bioeng. 2016, 113, 1617–1627. [Google Scholar] [CrossRef] [PubMed]
- Chansaenroj, J.; Chuchaona, W.; Thanusuwannasak, T.; Duang-In, A.; Puenpa, J.; Vutithanachot, V.; Vongpunsawad, S.; Poovorawan, Y. Prevalence of poliovirus vaccine strains in randomized stool samples from 2010 to 2018: Encompassing transition from the trivalent to bivalent oral poliovirus vaccine. Virusdisease 2019, 30, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Chabeda, A.; Van Zyl, A.R.; Rybicki, E.P.; Hitzeroth, I.I. Substitution of human papillomavirus type 16 L2 neutralizing epitopes into L1 surface loops: The Effect on virus-like particle assembly and immunogenicity. Front. Plant Sci. 2019, 10, 779. [Google Scholar] [CrossRef] [PubMed]
- Mbewana, S.; Meyers, A.E.; Rybicki, E.P. Chimaeric rift valley fever virus-like particle vaccine candidate production in Nicotiana Benthamiana. Biotechnol. J. 2019, 14, e1800238. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, V.; Baztarrica, J.; Rybicki, E.P.; Meyers, A.E.; Wigdorovitz, A. Minimally processed crude leaf extracts of Nicotiana benthamiana containing recombinant foot and mouth disease virus-like particles are immunogenic in mice. Biotechnol. Rep. (Amst) 2018, 20, e00283. [Google Scholar] [CrossRef]
- Mortola, E.; Roy, P. Efficient assembly and release of SARS coronavirus-like particles by a heterologous expression system. FEBS Lett. 2004, 576, 174–178. [Google Scholar] [CrossRef]
- Ho, Y.; Lin, P.H.; Liu, C.Y.; Lee, S.P.; Chao, Y.C. Assembly of human severe acute respiratory syndrome coronavirus-like particles. Biochem. Biophys. Res. Commun. 2004, 318, 833–838. [Google Scholar] [CrossRef]
- Lu, X.; Chen, Y.; Bai, B.; Hu, H.; Tao, L.; Yang, J.; Chen, J.; Chen, Z.; Hu, Z.; Wang, H. Immune responses against severe acute respiratory syndrome coronavirus induced by virus-like particles in mice. Immunology 2007, 122, 496–502. [Google Scholar] [CrossRef]
- Bai, B.; Hu, Q.; Hu, H.; Zhou, P.; Shi, Z.; Meng, J.; Lu, B.; Huang, Y.; Mao, P.; Wang, H. Virus-like particles of SARS-like coronavirus formed by membrane proteins from different origins demonstrate stimulating activity in human dendritic cells. PLoS ONE 2008, 3, e2685. [Google Scholar] [CrossRef]
- Lu, B.; Huang, Y.; Huang, L.; Li, B.; Zheng, Z.; Chen, Z.; Chen, J.; Hu, Q.; Wang, H. Effect of mucosal and systemic immunization with virus-like particles of severe acute respiratory syndrome coronavirus in mice. Immunology 2010, 130, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.V.; Massare, M.J.; Barnard, D.L.; Kort, T.; Nathan, M.; Wang, L.; Smith, G. Chimeric severe acute respiratory syndrome coronavirus (SARS-CoV) S glycoprotein and influenza matrix 1 efficiently form virus-like particles (VLPs) that protect mice against challenge with SARS-CoV. Vaccine 2011, 29, 6606–6613. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zheng, X.; Gai, W.; Zhao, Y.; Wang, H.; Wang, H.; Feng, N.; Chi, H.; Qiu, B.; Li, N.; et al. MERS-CoV virus-like particles produced in insect cells induce specific humoural and cellular imminity in rhesus macaques. Oncotarget 2017, 8, 12686–12694. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Takami, Y.; Deo, V.K.; Park, E.Y. Preparation of virus-like particle mimetic nanovesicles displaying the S protein of Middle East respiratory syndrome coronavirus using insect cells. J. Biotechnol. 2019, 306, 177–184. [Google Scholar] [CrossRef]
- Kost, T.A.; Condreay, J.P.; Jarvis, D.L. Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Rev. Nat. Biotechnol. 2005, 23, 567–575. [Google Scholar] [CrossRef]
- Altmann, F.; Schwihla, H.; Staudacher, E.; Glössl, J.; März, L. Insect cells contain an unusual, membrane-bound beta-N-acetylglucosaminidase probably involved in the processing of protein N-glycans. J. Biol. Chem. 1995, 270, 17344–17349. [Google Scholar] [CrossRef]
- COVID-19: Medicago’s Development Programs. Available online: https://www.medicago.com/en/covid-19-programs/ (accessed on 25 March 2020).
- Masters, P.S. The molecular biology of coronaviruses. Adv. Virus Res. 2006, 66, 193–292. [Google Scholar]
- Stephen, S.L.; Beales, L.; Peyret, H.; Roe, A.; Stonehouse, N.J.; Rowlands, D.J. Recombinant expression of tandem-HBc virus-like particles (VLPs). Methods Mol. Biol. 2018, 1776, 97–123. [Google Scholar]
- Balke, I.; Zeltins, A. Recent advances in the use of plant virus-like particles as vaccines. Viruses 2020, 12, 270. [Google Scholar] [CrossRef]
- Lerouge, P.; Bardor, M.; Pagny, S.; Gomord, V.; Faye, L. N-glycosylation of recombinant pharmaceutical glycoproteins produced in transgenic plants: Towards an humanisation of plant N-glycans. Curr. Pharm. Biotechnol. 2000, 1, 347–354. [Google Scholar] [CrossRef]
- Hamorsky, K.T.; Kouokam, J.C.; Jurkiewicz, J.M.; Nelson, B.; Moore, L.J.; Husk, A.S.; Kajiura, H.; Fujiyama, K.; Matoba, N. N-glycosylation of cholera toxin B subunit in Nicotiana benthamiana: Impacts on host stress response, production yield and vaccine potential. Sci. Rep. 2015, 5, 8003. [Google Scholar] [CrossRef]
- Schähs, M.; Strasser, R.; Stadlmann, J.; Kunert, R.; Rademacher, T.; Steinkellner, H. Production of a monoclonalantibody in plants with a humanized N-glycosylation pattern. Plant Biotechnol. J. 2007, 5, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, L.; Kuwahara, K.; Li, L.; Liu, Z.; Li, T.; Zhu, H.; Liu, J.; Xu, Y.; Xie, J.; et al. Immunodominant SARS coronavirus epitopes in humans elicited both enhancing and neutralizing effects on infection in non-human primates. ACS Infect. Dis. 2016, 2, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Wu, C.; Li, X.; Song, Y.; Yao, X.; Wu, X.; Duan, Y.; Zhang, H.; Wang, Y.; Qian, Z.; et al. On the origin and continuing evolution of SARS-CoV-2. Natl. Sci. Rev. 2020. [Google Scholar] [CrossRef]
- Phan, T. Genetic diversity and evolution of SARS-CoV-2. Infect. Genet. Evol. 2020, 81, 104260. [Google Scholar] [CrossRef]
- Channappanavar, R.; Fett, C.; Zhao, J.; Meyerholz, D.K.; Perlman, S. Virus-specific memory CD8 T cells provide substantial protection from lethal severe acute respiratory syndrome coronavirus infection. J. Virol. 2014, 88, 11034–11044. [Google Scholar] [CrossRef]
- Freytag, L.C.; Clements, J.D. Mucosal adjuvants. Vaccine 2005, 23, 1804–1813. [Google Scholar] [CrossRef]
- Mason, H.S.; Haq, T.A.; Clements, J.D.; Arntzen, C.J. Edible vaccine protects mice against Escherichia coli heat-labile enterotoxin (LT): Potatoes expressing a synthetic LT-B gene. Vaccine 1998, 16, 1336–1343. [Google Scholar] [CrossRef]
- Govea, D.O.; Rubio, N.; García, A.L.; Varona, J.T.; Korban, S.S.; Moreno, L.; Rosales, S. Immunogenic properties of a lettuce-derived C4(V3)6 multiepitopic HIV protein. Planta 2013, 238, 785–792. [Google Scholar] [CrossRef]
- Trujillo, E.; Rosales, S.; Angulo, C. A multi-epitope plant-made chimeric protein (LTBentero) targeting common enteric pathogens is immunogenic in mice. Plant Mol. Biol. 2020, 102, 159–169. [Google Scholar] [CrossRef]
- Orellana, L.; Rosales, S.; Romero, A.; Parsons, J.; Decker, E.L.; Monreal, E.; Moreno, L.; Reski, R. An Env-derived multi-epitope HIV chimeric protein produced in the moss Physcomitrella patens is immunogenic in mice. Plant Cell Rep. 2015, 34, 425–433. [Google Scholar] [CrossRef]
- Monreal, E.; Bañuelos, B.; Hernández, M.; Fragoso, G.; Garate, T.; Sciutto, E.; Rosales, S. Expression of multiple Taenia Solium Immunogens in plant cells through a ribosomal skip mechanism. Mol. Biotechnol. 2015, 57, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Jong, J.M.; Schuurhiuis, D.; Joan-Facsinay, A.; van der Voort, E.; Huizinga, T.; Ossendorp, F.; Toes, R.; Verbeek, J. Murine Fc receptors for IgG are redundant in facilitating presentation of immune complex derived antigen to CD8+ T cells in vivo. Mol. Immunol. 2006, 43, 2045–2050. [Google Scholar] [CrossRef] [PubMed]
- Marusić-Galesić, S.; Marusić, M.; Pokrić, B. Cellular immune response to the antigen administered as an immune complex in vivo. Immunology 1992, 75, 325–329. [Google Scholar] [PubMed]
- Diamos, A.G.; Hunter, J.G.L.; Pardhe, M.D.; Rosenthal, S.H.; Sun, H.; Foster, B.C.; DiPalma, M.P.; Chen, Q.; Mason, H.S. High level production of monoclonal antibodies using an optimized plant expression system. Front. Bioeng. Biotechnol. 2020, 7, 472. [Google Scholar] [CrossRef] [PubMed]
- Pepponi, I.; Diogo, G.R.; Stylianou, E.; van Dolleweerd, C.J.; Drake, P.M.W.; Paul, M.J.; Sibley, L.; Ma, J.K.-C.; Reljic, R. Plant-derived recombinant immune complexes as self-adjuvanting TB immunogens for mucosal boosting of BCG. Plant Biotechnol. J. 2014, 12, 840–850. [Google Scholar] [CrossRef]
- Phoolcharoen, W.; Bhoo, S.H.; Lai, H.; Ma, J.; Arntzen, C.J.; Chen, Q.; Mason, H.S. Expression of an immunogenic Ebola immune complex in Nicotiana benthamiana. Plant Biotechnol. J. 2011, 9, 807–816. [Google Scholar] [CrossRef]
- Floss, D.M.; Schallau, K.; Rose-John, S.; Conrad, U.; Scheller, J. Elastin-like polypeptides revolutionize protein expression and their biomedical application. Trends Biotechnol. 2010, 28, 37–45. [Google Scholar] [CrossRef]
- Floss, D.M.; Mockey, M.; Zanello, G.; Brosson, D.; Diogon, M.; Frutos, R.; Bruel, T.; Rodrigues, V.; Garzon, E.; Chevaleyre, C.; et al. Expression and immunogenicity of the mycobacterial Ag85B/ESAT-6 antigens produced in transgenic plants by elastin-like peptide fusion strategy. Biomed. Biotechnol. 2010, 274346. [Google Scholar] [CrossRef]
- Phan, H.T.; Pohl, J.; Floss, D.M.; Rabenstein, F.; Veits, J.; Le, B.T.; Chu, H.H.; Hause, G.; Mettenleiter, T.; Conrad, U. ELPylated haemagglutinins produced in tobacco plants induce potentially neutralizing antibodies against H5N1 viruses in mice. Plant Biotechnol. J. 2013, 11, 582–593. [Google Scholar] [CrossRef]
- Li, H.Y.; Chye, M.L. Use of GFP to investigate expression of plant-derived vaccines. Methods Mol. Biol. 2009, 515, 275–285. [Google Scholar]
- Rubio, N.; Govea, D.O.; Romero, A.; García, A.L.; Ilhuicatzi, D.; Salazar, J.A.; Korban, S.S.; Rosales-Mendoza, S.; Moreno-Fierros, L. A plant-derived Multi-HIV antigen induces broad immune responses in orally immunized mice. Mol. Biotechnol. 2015, 57, 622–674. [Google Scholar]
- Andrew, M.K.; Bowles, S.K.; Pawelec, G.; Haynes, L.; Kuchel, G.A.; McNeil, S.A.; McElhaney, J.E. Influenza vaccination in older adults: Recent innovations and practical applications. Drugs Aging 2019, 36, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Miquel-Clopés, A.; Bentley, E.G.; Stewart, J.P.; Carding, S.R. Mucosal vaccines and technology. Clin. Exp. Immunol. 2019, 196, 205–214. [Google Scholar] [CrossRef] [PubMed]
- iBio. Available online: https://www.ibioinc.com/ (accessed on 25 March 2020).
- Chan, H.T.; Xiao, Y.; Weldon, W.C.; Oberste, S.M.; Chumakov, K.; Daniell, H. Cold chain and virus-free chloroplast-made booster vaccine to confer immunity against different poliovirus serotypes. Plant Biotechnol. J. 2016, 14, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Margolin, E.; Chapman, R.; Meyers, A.E.; van Diepen, M.T.; Ximba, P.; Hermanus, T.; Crowther, C.; Weber, B.; Morris, L.; Williamson, A.L.; et al. Production and immunogenicity of soluble plant-produced HIV-1 subtype C envelope gp140 immunogens. Front. Plant Sci. 2019, 10, 1378. [Google Scholar] [CrossRef]
- Kardani, K.; Bolhassani, A.; Shahbazi, S. Prime-boost vaccine strategy against viral infections: Mechanisms and benefits. Vaccine 2016, 34, 413–423. [Google Scholar] [CrossRef]



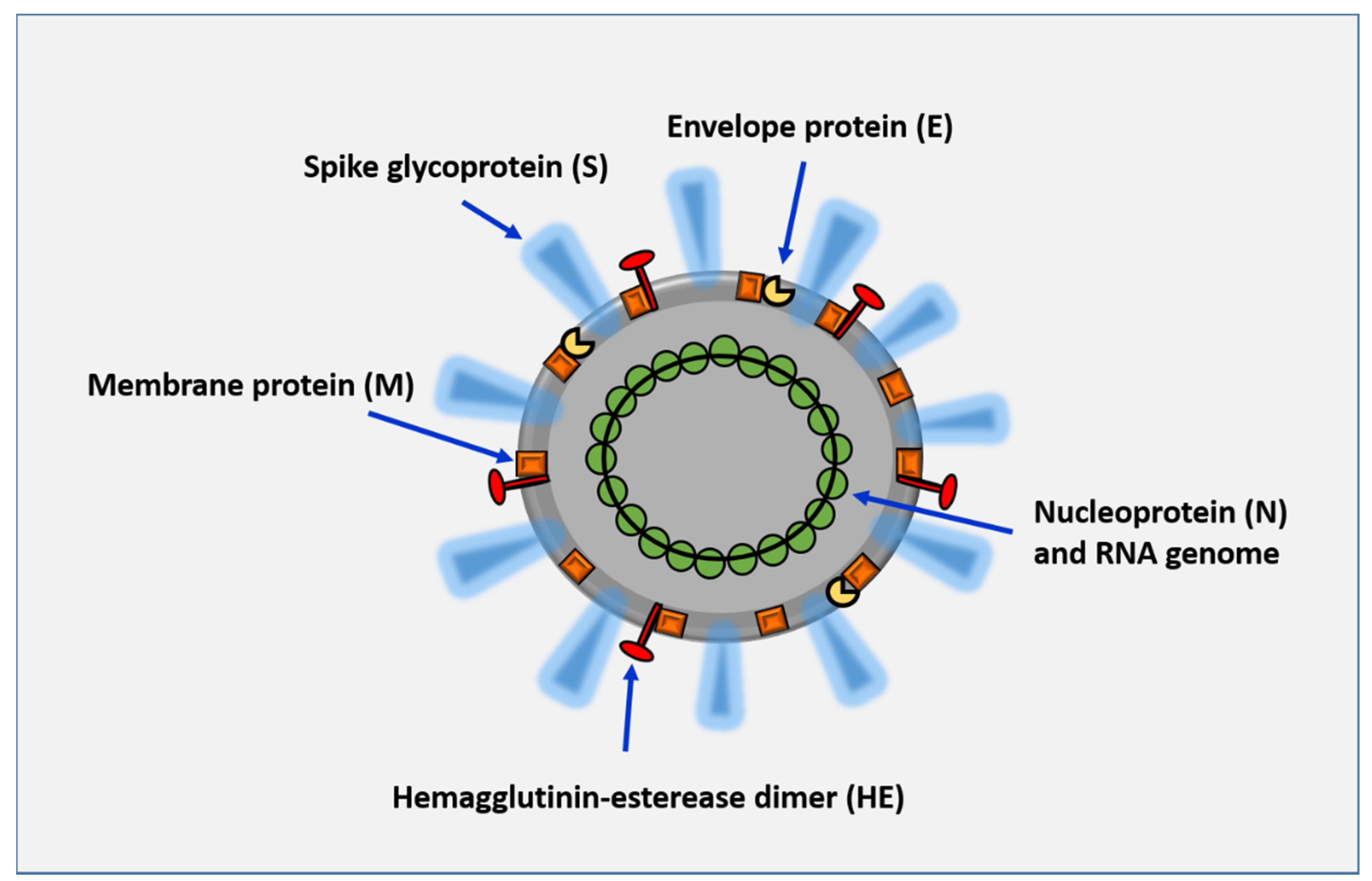{kind=link}
{kind=link}
| Approach | Attractive Features | Drawbacks | Proposed Target Antigens | MERS/SARS Precedents | Reference |
|---|---|---|---|---|---|
| Stable nuclear genome transformation | Inheritable antigen production, allows seed bank generation; post-translational modifications are performed; protocols available for several species including seed crops | Non-site specific transgene insertion; horizontal gene transfer is possible; transgene expression affected by position effects and silencing; transformation takes long time | S protein; multiepitope vaccines | The N-terminal fragment of the SARS-CoV-1 S protein (S1) was expressed in stably transformed tomato and low-nicotine tobacco plants, which induced IgA and IgG responses in mice. | [32] |
| Transient nuclear genome transformation | Rapid production; high productivity; implemented at the industrial level | Seed bank cannot be generated; requires purification of the antigen to eliminate toxic compounds from the host and ag-robacteria residues | S protein; multiepitope vaccines | A chimeric protein of GFP and amino acids 1-658 of the SARS-CoV-1 S protein (S1:GFP) was transiently expressed in tobacco leaves and stably transformed in tobacco and lettuce. No immunization assays were performed The SARS-CoV-1 N protein was transiently expressed in Nicotiana benthamiana, which induced in mice high levels of IgG1 and IgG2a and up regulation of IFN-γ and IL-10 in splenocytes. A chimeric protein of GFP and the SARS-CoV-1 S protein was transiently expressed in tobacco plants. No immunization tests were performed. The SARS-CoV-1 M and N proteins were transiently expressed in N. benthamiana. The N protein was antigenic but immunogenicity was not assessed. | [33,34,35] |
| Transplastomic technologies | High productivity; multigene expression is possible; improved biosafety since the transgene is inherited maternally; site-specific insertion through recombination; not affected by silencing or position effects | Complex post-translational modifications are not performed; transformation protocols available for few species and the generation of lines takes long time. | Multiepitope vaccines | A chimeric protein of GFP and amino acids 1-658 of the SARS-CoV-1 S (S1:GFP) was expressed in transplastomic tobacco plants. | [32,36] |
| VLPs Produced and Immunization Approach | Findings | Reference |
|---|---|---|
| Haemagglutinin (HA) from H1N1 A/California/07/2009 (pdmH1N1). Mice were immunized twice by the intramuscular (i.m.) route or by combining i.m. priming and (intranasal) i.n. boosting. | VLP influenza vaccination exhibited high levels of antibodies titers compared to the inactivated influenza vaccine (IIV). Lung homogenate displayed chemokine/cytokine levels and virus loads lower in the VLP groups compared to the IIV group. | [45] |
| HA from strains A/California/07/2009 H1N1 (H1/Cal), A/Victoria/361/11 H3N2 (H3/Vic), B/Brisbane/60/08 (B/Bris, Victoria lineage), and B/Massachusetts/02/2012 (B/Mass, Yamagata lineage). In a Phase II clinical trial, subjects were immunized with a single i.m. dose using Alhydrogel as an adjuvant. | Both homologous and heterologous antigen-specific CD4+ T cells were elicited. Additionally, production of IFN-γ, IL-2, and/or TNF-α was achieved upon ex vivo antigenic re-stimulation. | [46] |
| HA from A/California/07/2009 H1N1. VLPs were evaluated in vitro using human monocyte-derived macrophages. | The plant-made VLPs were efficiently captured and subjected to endosomal processing and cross-presentation. | [47] |
| HA from A/H1N1/California/07/09 (pdmH1N1). The inactivated H1N1 vaccine (IIV) was included as a reference vaccine. Mice were i.m.-immunized twice. | CD4+ (TNF-α, IFN-γ) and CD8+ (IFN-γ) T cell responses were higher for the plant-made vaccine than the IIV formulation. The plant-made VLP vaccine elicited stronger and more balanced immune responses than IIV. | [48] |
| HA from A/California/7/09 (H1N1) and A/Indonesia/5/05 (H5N1). In vitro assays were performed using mouse and human DCs. Mice were immunized by the i.m. route using Alhydrogel as an adjuvant. | Human DCs exposed to plant-made VLPs showed high stimulation in terms of secretion of IL-6, IL-10, and TNFα and CD83 expression, along with activation of CD4+ and CD8+ T cells. VLPs induced accumulation of both cDC1s and cDC2 in the draining lymph nodes of test mice. Lymphocytes from mice immunized with plant-made VLPs secreted higher concentrations of pro-inflammatory cytokines (including IL-12, IL-6, and TNF-α) upon antigenic stimulation. | [49] |
| HA from A/California/7/2009 or A/Indonesia/5/05 strains. In vitro assays were performed using mouse dendritic cells. | The plant-made VLPs trimmers were morphologically stable over time and interacted with activated antigen-presenting cells similar to the wild type virus. | [50] |
| Approach | Attractive Features | Drawbacks | References |
|---|---|---|---|
| Epitopes fused to B subunits of cholera toxin or the heat labile enterotoxin (CTB/LTB) | Compatible with mucosal immunization | May induce tolerance | [56,57] |
| VLPs (native or chimeric) | Simple purification | Chimeric VLPs often have limitations on the insert size (unrelated antigen) | [58] |
| Immune complexes | Simple purification | High expression is required to form the complexes | [59] |
| ELPylated antigens | Robust expression Simple purification | Low yields when large number of ELP moieties are included | [60] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosales-Mendoza, S.; Márquez-Escobar, V.A.; González-Ortega, O.; Nieto-Gómez, R.; Arévalo-Villalobos, J.I. What Does Plant-Based Vaccine Technology Offer to the Fight against COVID-19? Vaccines 2020, 8, 183. https://doi.org/10.3390/vaccines8020183
Rosales-Mendoza S, Márquez-Escobar VA, González-Ortega O, Nieto-Gómez R, Arévalo-Villalobos JI. What Does Plant-Based Vaccine Technology Offer to the Fight against COVID-19? Vaccines. 2020; 8(2):183. https://doi.org/10.3390/vaccines8020183
Chicago/Turabian StyleRosales-Mendoza, Sergio, Verónica A. Márquez-Escobar, Omar González-Ortega, Ricardo Nieto-Gómez, and Jaime I. Arévalo-Villalobos. 2020. "What Does Plant-Based Vaccine Technology Offer to the Fight against COVID-19?" Vaccines 8, no. 2: 183. https://doi.org/10.3390/vaccines8020183
APA StyleRosales-Mendoza, S., Márquez-Escobar, V. A., González-Ortega, O., Nieto-Gómez, R., & Arévalo-Villalobos, J. I. (2020). What Does Plant-Based Vaccine Technology Offer to the Fight against COVID-19? Vaccines, 8(2), 183. https://doi.org/10.3390/vaccines8020183