Increased Immunogenicity of Full-Length Protein Antigens through Sortase-Mediated Coupling on the PapMV Vaccine Platform


Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Production of the Recombinant Proteins
2.2. Coupling Reactions with SrtA
2.3. SDS-PAGE for Coupling Efficacy and Western Blot
2.4. Immunization and Immune Response
2.5. Statistics
2.6. Ethics Statement
3. Results
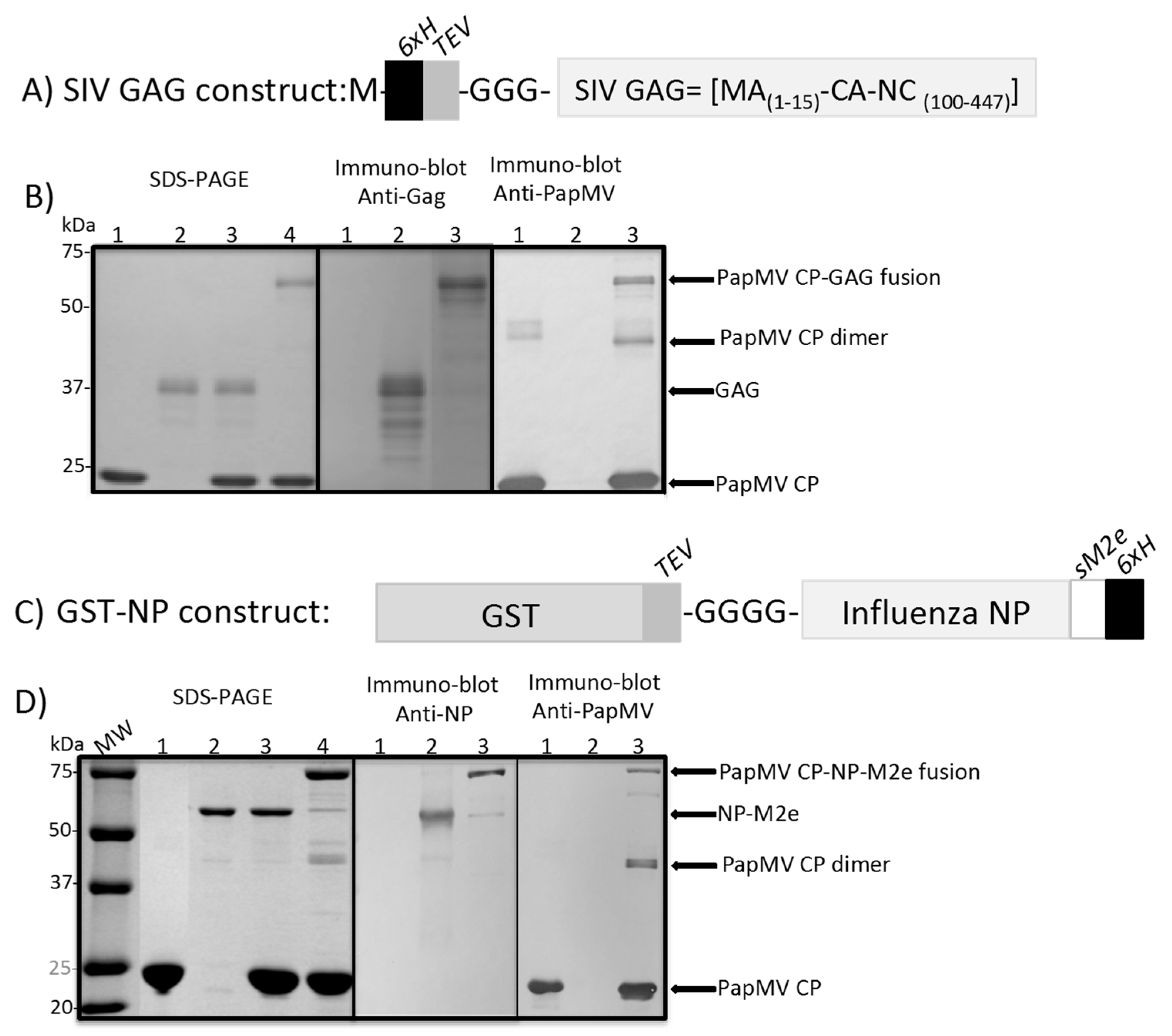
3.1. Coupling of GAG and NP to the PapMV Nanoparticles
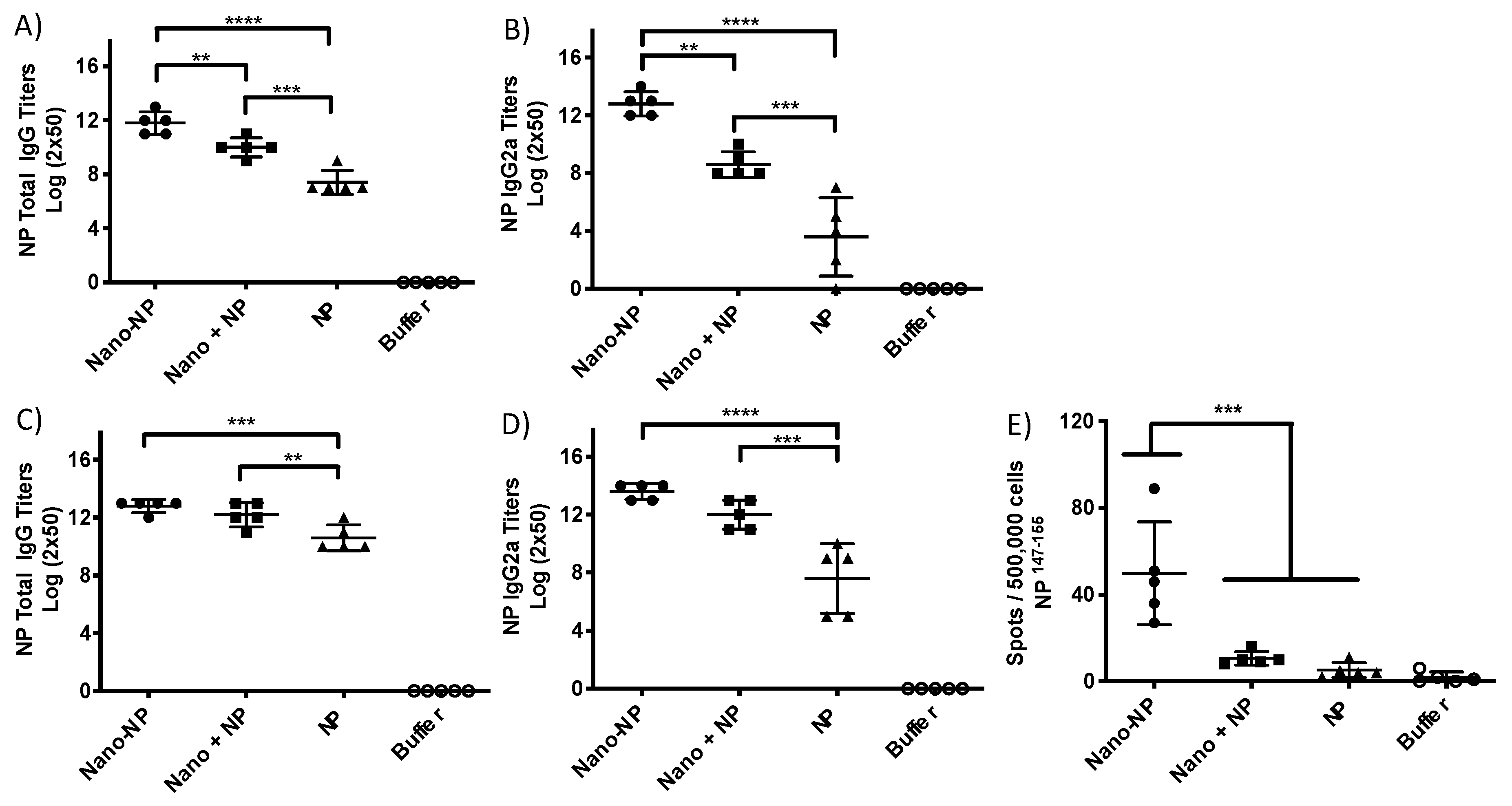
3.2. Assessment of the Immune Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CP | coat proteins |
| CTL | cytotoxic T lymphocyte |
| ELISA | enzyme-linked immunosorbent assay |
| ELISPOT | enzyme-linked immunospot |
| GAG | group-specific antigen |
| HPV | Human papilloma virus |
| M2e | matrix protein 2 ectodomain |
| Nano | nanoparticles |
| NP | nucleoprotein |
| PapMV | papaya mosaic virus |
| SIV | simian immunodeficiency virus |
| SrtA | sortase A |
| ssRNA | single stranded RNA |
| TLR | toll like receptor |
| VLPs | virus-like particles |
References
- Al-Halifa, S.; Gauthier, L.; Arpin, D.; Bourgault, S.; Archambault, D. Nanoparticle-Based Vaccines Against Respiratory Viruses. Front Immunol. 2019, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Lebel, M.E.; Daudelin, J.F.; Chartrand, K.; Tarrab, E.; Kalinke, U.; Savard, P.; Labrecque, N.; Leclerc, D.; Lamarre, A. Nanoparticle Adjuvant Sensing by TLR7 enhances CD8+ T Cell-Mediated protection from listeria Monocytogenes infection. J. Immunol. 2014, 192, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, D. Plant Viral Epitope Display Systems for Vaccine Development. Curr. Top. Microbiol. Immunol. 2014, 183, 2–13. [Google Scholar]
- Rioux, G.; Babin, C.; Majeau, N.; Leclerc, D. Engineering of papaya mosaic virus (PapMV) nanoparticles through fusion of the HA11 peptide to several putative surface-exposed sites. PLoS ONE 2012, 7, e31925. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.M. Human papillomavirus and vaccination. Mayo Clin. Proc. 2008, 83, 701–707. [Google Scholar] [CrossRef]
- Stanley, M. Immunobiology of HPV and HPV vaccines. Gynecol. Oncol. 2008, 109, S15–S21. [Google Scholar] [CrossRef] [PubMed]
- Thérien, A.; Bédard, M.; Carignan, D.; Rioux, G.; Gauthier-Landry, L.; Laliberté-Gagné, M.E.; Bolduc, M.; Savard, P.; Leclerc, D. A versatile papaya mosaic virus (PapMV) vaccine platform based on sortase-mediated antigen coupling. J. Nanobiotechnol. 2017, 15, 54. [Google Scholar] [CrossRef]
- Mathieu, C.; Rioux, G.; Dumas, M.C.; Leclerc, D. Induction of innate immunity inlungs with virus-like nanoparticles leads to protection against influenza and Streptococcus pneumoniae challenge. Nanomedecine 2013, 9, 839–848. [Google Scholar] [CrossRef]
- Carignan, D.; Herblot, S.; Laliberté-Gagné, M.E.; Bolduc, M.; Duval, M.; Savard, P.; Leclerc, D. Activation of innate immunity in primary human cells using a plant virus derived nanoparticle TLR7/8 agonist. Nanomedecine 2018, 14, 2317–2327. [Google Scholar] [CrossRef]
- Bolduc, M.; Baz, M.; Laliberté-Gagné, M.E.; Carignan, D.; Garneau, C.; Russel, A.; Boivin, G.; Savard, P.; Leclerc, D. The Quest for a nanoparticle-based vaccine inducing broad protection to influenza viruses. Nanomedecine 2018, 14, 2563–2574. [Google Scholar] [CrossRef]
- Carignan, D.; Thérien, A.; Rioux, G.; Paquet, G.; Laliberté-Gagné, M.E.; Bolduc, M.; Savard, P.; Leclerc, D. Engineering of the PapMV vaccine platform with a shortened M2e peptide leads to an effective one dose influenza vaccine. Vaccine 2015, 16, 7245–7253. [Google Scholar] [CrossRef] [PubMed]
- Denis, J.; Acosta-Ramirez, E.; Zhao, Y.; Hamelin, M.E.; Koukavica, I.; Baz, M.; Abed, Y.; Savard, C.; Pare, C.; Lopez Macias, C.; et al. Development of a Universal Influenza A vaccine based on the M2e peptide fused to the Papaya Mosaic Virus (PapMV) vaccine platform. Vaccine 2008, 26, 3395–3403. [Google Scholar] [CrossRef] [PubMed]
- Babin, C.; Majeau, N.; Leclerc, D. Engineering of papaya mosaic virus (PapMV) nanoparticles with a CTL epitope derived from influenza NP. J. Nanobiotechnol. 2013, 11, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Clancy, K.W.; Melvin, J.; McCafferty, D.G. Sortases transpeptidases: Insights into mechanism, substrate specificity, and inhibition. Biopolymers 2010, 4, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Ritzefeld, M. Sortagging: A robust and efficient chemoenzymatic ligation strategy. Chemistry 2014, 20, 8516–8529. [Google Scholar] [CrossRef] [PubMed]
- Spirig, T.; Weiner, E.M.; Clubb, R.T. Sortase enzymes in gram positive bacteria. Mol. Microbiol. 2011, 82, 1044–1059. [Google Scholar] [CrossRef] [PubMed]
- Rioux, G.; Mathieu, C.; Russell, A.; Bolduc, M.; Laliberté-Gagné, M.E.; Savard, P.; Leclerc, D. PapMV nanoparticles Improve Mucosal Immune Responses to the Trivalent Inactivated Flu Vaccine. J. Nanobiotechnol. 2014, 12, 19. [Google Scholar] [CrossRef]
- Rioux, G.; Carignan, D.; Russell, A.; Bolduc, M.; Laliberté-Gagné, M.E.; Savard, P.; Leclerc, D. Influence of PapMV nanoparticles on the kinetics of the antibody response to flu vaccine. J. Nanobiotechnol. 2016, 14, 43. [Google Scholar] [CrossRef]
- Savard, C.; Guérin, A.; Drouin, K.; Bolduc, M.; Laliberté-Gagné, M.E.; Dumas, M.C.; Majeau, N.; Leclerc, D. Improvement of the Trivalent Inactivated Flu Vaccine Using PapMV Nanoparticles. PLoS ONE 2011, 6, e21522. [Google Scholar] [CrossRef]
- Kleijmeer, M.J.; Escola, J.M.; UytdeHaag, F.G.; Jakobobson, E.; Griffith, J.M.; Osterhous, A.D.; Stoorvogel, W.; Melief, C.J.; Rabouille, C.; Geuze, H.J. Antigen loading of MHC class I molecules in the endocytic tract. Traffic 2001, 2, 124–137. [Google Scholar] [CrossRef]
- Lebel, M.E.; Chartrand, K.; Leclerc, D.; Lamarre, A. Plant viruses as Nanoparticles-Based Vaccines and Adjuvants. Vaccines 2015, 3, 620–637. [Google Scholar] [CrossRef] [PubMed]
- Lebel, M.E.; Chartrand, K.; Tarrab, E.; Savard, P.; Leclerc, D.; Lamarre, A. Potentiating cancer immunotherapy using Papaya mosaic virus-derived nanoparticles. Nano Lett. 2016, 16, 1826–1832. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.P.; Zhang, R.; Xu, Z.P. Nanoparticle-Based Nanomedicines to Promote Cancer Immunotherapy: Recent Advances and Future Directions. Small 2019, 25, e1900262. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laliberté-Gagné, M.-È.; Bolduc, M.; Thérien, A.; Garneau, C.; Casault, P.; Savard, P.; Estaquier, J.; Leclerc, D. Increased Immunogenicity of Full-Length Protein Antigens through Sortase-Mediated Coupling on the PapMV Vaccine Platform. Vaccines 2019, 7, 49. https://doi.org/10.3390/vaccines7020049
Laliberté-Gagné M-È, Bolduc M, Thérien A, Garneau C, Casault P, Savard P, Estaquier J, Leclerc D. Increased Immunogenicity of Full-Length Protein Antigens through Sortase-Mediated Coupling on the PapMV Vaccine Platform. Vaccines. 2019; 7(2):49. https://doi.org/10.3390/vaccines7020049
Chicago/Turabian StyleLaliberté-Gagné, Marie-Ève, Marilène Bolduc, Ariane Thérien, Caroline Garneau, Philippe Casault, Pierre Savard, Jérome Estaquier, and Denis Leclerc. 2019. "Increased Immunogenicity of Full-Length Protein Antigens through Sortase-Mediated Coupling on the PapMV Vaccine Platform" Vaccines 7, no. 2: 49. https://doi.org/10.3390/vaccines7020049
APA StyleLaliberté-Gagné, M.-È., Bolduc, M., Thérien, A., Garneau, C., Casault, P., Savard, P., Estaquier, J., & Leclerc, D. (2019). Increased Immunogenicity of Full-Length Protein Antigens through Sortase-Mediated Coupling on the PapMV Vaccine Platform. Vaccines, 7(2), 49. https://doi.org/10.3390/vaccines7020049