
Mucosal Vaccination Against SARS-CoV-2 Using Human Probiotic Bacillus subtilis Spores as an Adjuvant Induces Potent Systemic and Mucosal Immunity
 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Vaccine Formulation and Inoculation
2.3. Blood Sampling
2.4. Saliva Collection
2.5. Transcardial Perfusion
2.6. Spleen Single-Cell Suspension
2.7. Lung Single-Cell Suspension
2.8. BALF Collection
2.9. NALT Isolation and Culture
2.10. Specific Antibody Determinations
2.11. Avidity Index
2.12. Neutralization Assay
2.13. CFSE-Proliferation Assay
2.14. Flow Cytometry
2.15. Experimental Design and Statistical Analyses
3. Results
3.1. SARS-CoV-2 RBD Intranasal Immunization Adjuvanted with Human Probiotic Bacillus subtilis Spores Induces Specific Systemic and Mucosal Responses
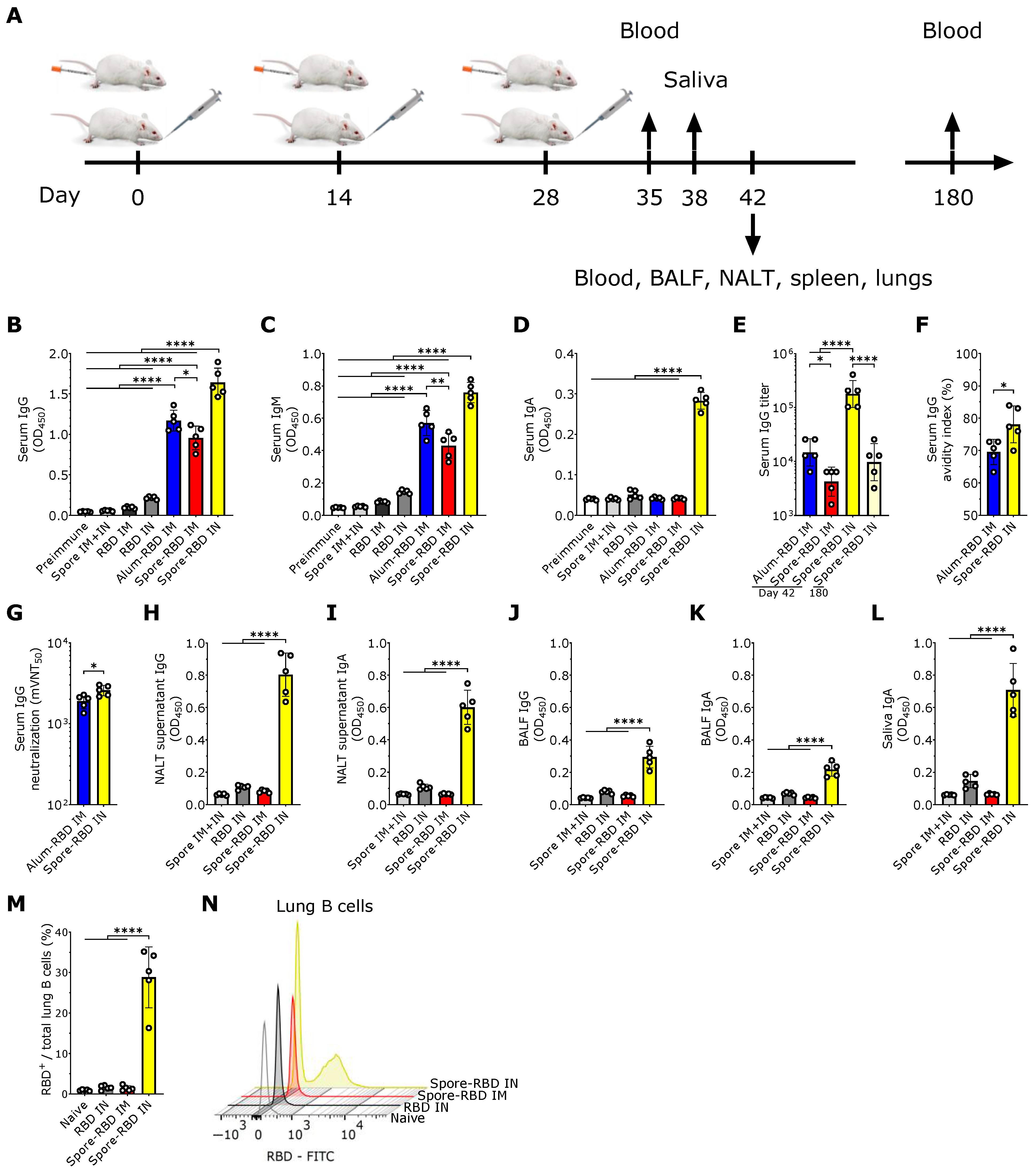
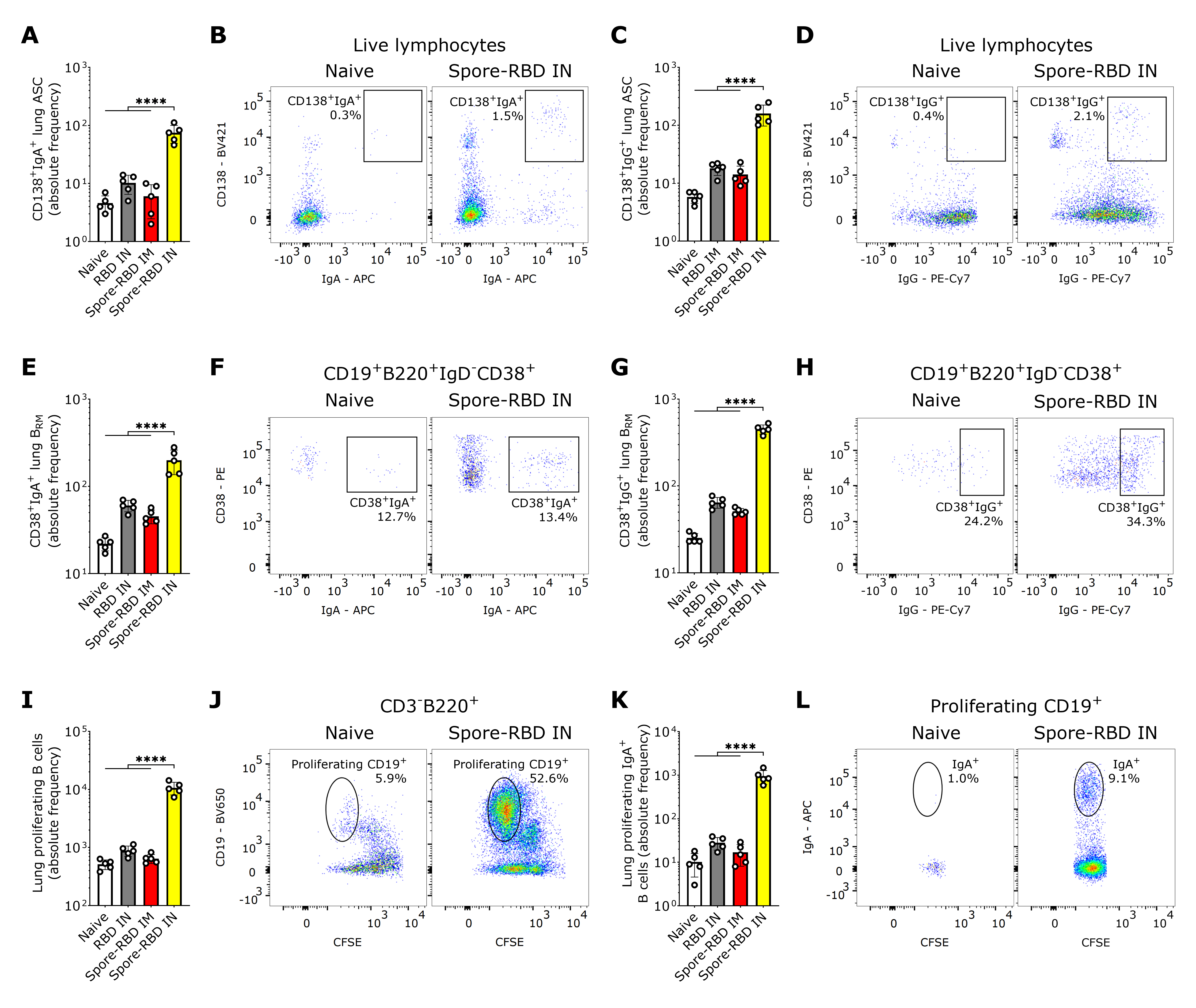
3.2. SARS-CoV-2 RBD Intranasal Immunization Adjuvanted with Human Probiotic Bacillus subtilis Spores Induces Mucosal B Cell Memory in the Lungs
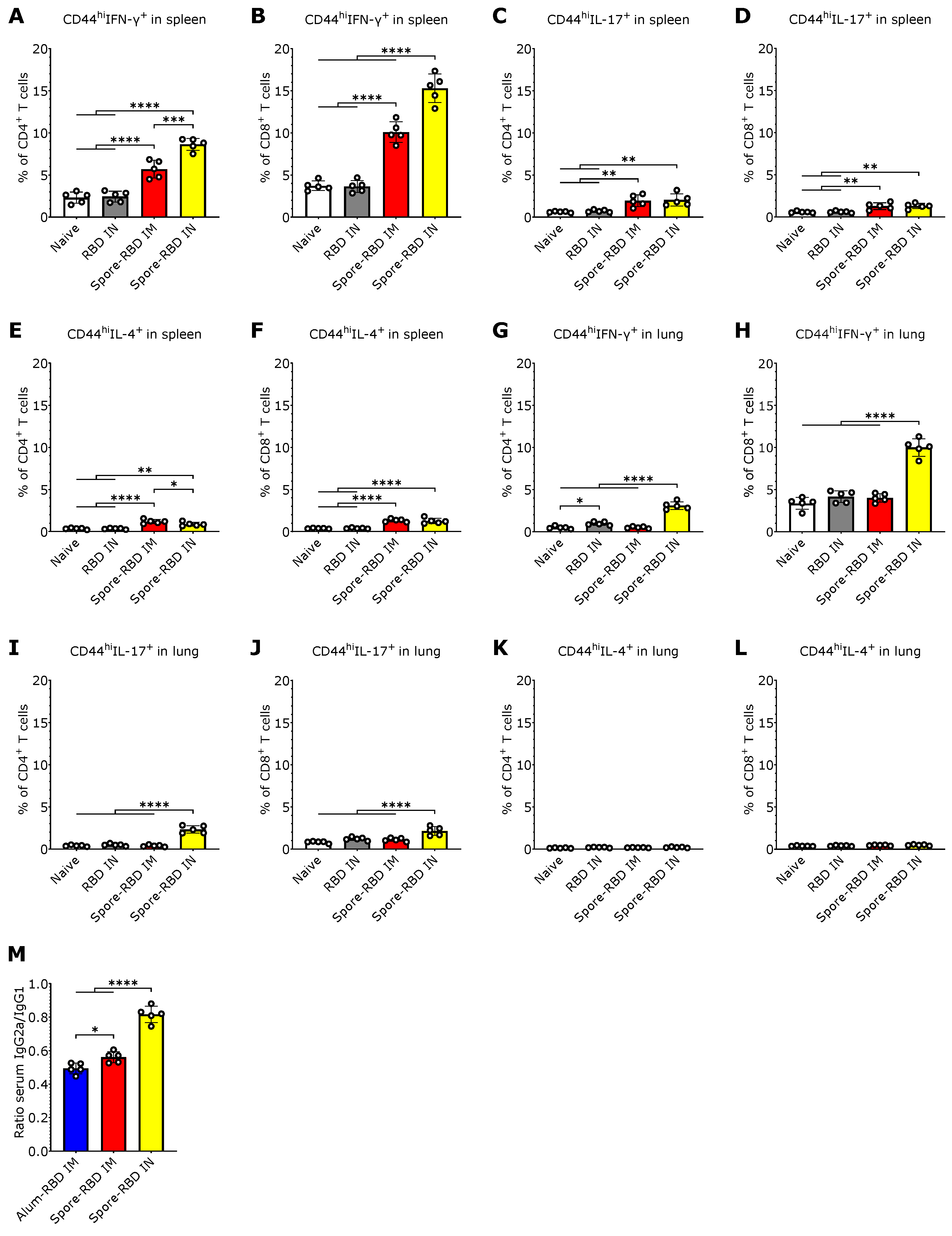
3.3. SARS-CoV-2 RBD Intranasal Immunization Adjuvanted with Human Probiotic Bacillus subtilis Spores Induces Systemic and Mucosal T Cell Immunity with a Th1 Bias
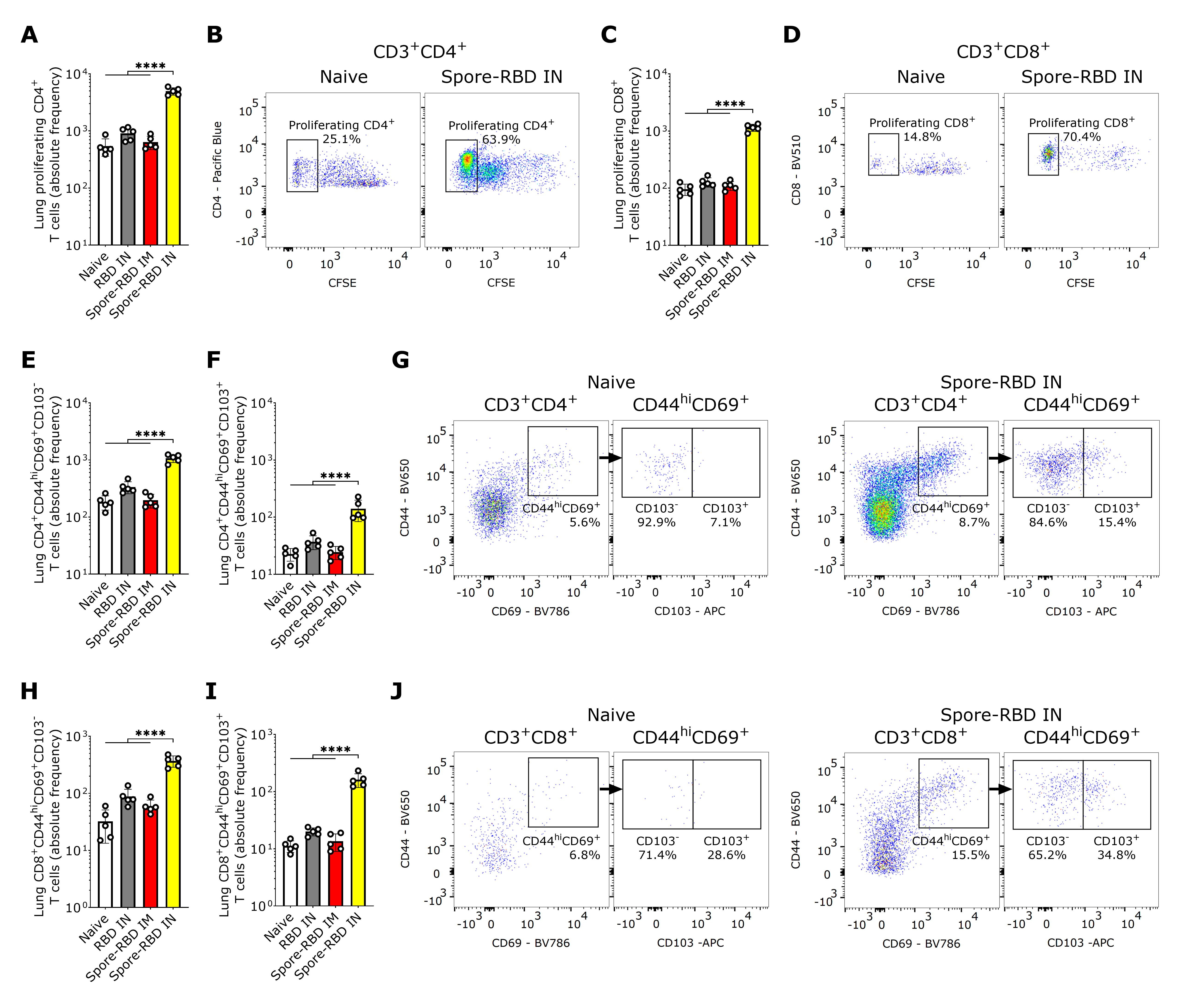
3.4. SARS-CoV-2 RBD Intranasal Immunization Adjuvanted with Human Probiotic Bacillus subtilis Spores Induces Mucosal T Cell Memory in the Lungs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Y.; Peng, F.; Wang, R.; Guan, K.; Jiang, T.; Xu, G.; Sun, J.; Chang, C. The deadly coronaviruses: The 2003 SARS pandemic and the 2020 novel coronavirus epidemic in China. J. Autoimmun. 2020, 109, 102434. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Chen, Y.; Qin, Q. Unique epidemiological and clinical features of the emerging 2019 novel coronavirus pneumonia (COVID-19) implicate special control measures. J. Med. Virol. 2020, 92, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef]
- World Health Organization. Weekly Epidemiological Update on COVID-19. Available online: https://data.who.int/dashboards/covid19 (accessed on 11 February 2025).
- Wang, H.; Paulson, K.R.; Pease, S.A.; Watson, S.; Comfort, H.; Zheng, P.; Aravkin, A.Y.; Bisignano, C.; Barber, R.M.; Alam, T. Estimating excess mortality due to the COVID-19 pandemic: A systematic analysis of COVID-19-related mortality, 2020–2021. Lancet 2022, 399, 1513–1536. [Google Scholar] [CrossRef] [PubMed]
- Fenner, R.; Cernev, T. The implications of the Covid-19 pandemic for delivering the Sustainable Development Goals. Futures 2021, 128, 102726. [Google Scholar] [CrossRef]
- World Health Organization. COVID-19–Landscape of Novel Coronavirus Candidate Vaccine Development Worldwide. Available online: https://www.who.int/teams/blueprint/covid-19/covid-19-vaccine-tracker-and-landscape (accessed on 11 February 2025).
- Mambelli, F.; de Araujo, A.C.V.; Farias, J.P.; de Andrade, K.Q.; Ferreira, L.C.; Minoprio, P.; Leite, L.C.; Oliveira, S.C. An Update on Anti-COVID-19 Vaccines and the Challenges to Protect Against New SARS-CoV-2 Variants. Pathogens 2025, 14, 23. [Google Scholar] [CrossRef]
- Hossain, M.K.; Hassanzadeganroudsari, M.; Feehan, J.; Apostolopoulos, V. The race for a COVID-19 vaccine: Where are we up to? Expert Rev. Vaccines 2022, 21, 355–376. [Google Scholar] [CrossRef]
- World Health Organization. Summary Status of COVID-19 Vaccines Within WHO EUL/PQ Evaluation Process—09 December 2024; WHO: Geneva, Switzerland, 2024. [Google Scholar]
- Singh, J.A.; Upshur, R.E. The granting of emergency use designation to COVID-19 candidate vaccines: Implications for COVID-19 vaccine trials. Lancet Infect. Dis. 2021, 21, e103–e109. [Google Scholar] [CrossRef]
- Ahirwar, K.; Rohila, A.; Shukla, R. Regulatory consideration and pathways for vaccine development. In Advanced Vaccination Technologies for Infectious and Chronic Diseases; Elsevier: Amsterdam, The Netherlands, 2024; pp. 325–339. [Google Scholar]
- Shah, S.S. Health Economics of Vaccine Development and Distribution: Lessons from the COVID-19 Pandemic. Public Health 2024, 1, 100015. [Google Scholar] [CrossRef]
- Lin, L.; Liu, Y.; Tang, X.; He, D. The disease severity and clinical outcomes of the SARS-CoV-2 variants of concern. Front. Public Health 2021, 9, 775224. [Google Scholar] [CrossRef]
- Charitos, I.A.; Ballini, A.; Lovero, R.; Castellaneta, F.; Colella, M.; Scacco, S.; Cantore, S.; Arrigoni, R.; Mastrangelo, F.; Dioguardi, M. Update on COVID-19 and Effectiveness of a Vaccination Campaign in a Global Context. Int. J. Environ. Res. Public Health 2022, 19, 10712. [Google Scholar] [CrossRef] [PubMed]
- McLean, G.; Kamil, J.; Lee, B.; Moore, P.; Schulz, T.F.; Muik, A.; Sahin, U.; Türeci, Ö.; Pather, S. The impact of evolving SARS-CoV-2 mutations and variants on COVID-19 vaccines. mBio 2022, 13, e0297921. [Google Scholar] [CrossRef]
- Focosi, D.; Quiroga, R.; McConnell, S.; Johnson, M.C.; Casadevall, A. Convergent evolution in SARS-CoV-2 spike creates a variant soup from which new COVID-19 waves emerge. Int. J. Mol. Sci. 2023, 24, 2264. [Google Scholar] [CrossRef]
- Callaway, E. The next generation of coronavirus vaccines. Nature 2023, 614, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Sarker, R.; Roknuzzaman, A.; Nazmunnahar; Shahriar, M.; Hossain, M.J.; Islam, M.R. The WHO has declared the end of pandemic phase of COVID-19: Way to come back in the normal life. Health Sci. Rep. 2023, 6, e1544. [Google Scholar] [CrossRef] [PubMed]
- Rathore, A.P.; John, A.L.S. Promises and challenges of mucosal COVID-19 vaccines. Vaccine 2023, 41, 4042–4049. [Google Scholar] [CrossRef]
- Alqahtani, S.A.M. Mucosal immunity in COVID-19: A comprehensive review. Front. Immunol. 2024, 15, 1433452. [Google Scholar] [CrossRef]
- Christensen, D.; Polacek, C.; Sheward, D.J.; Hanke, L.; Moliner-Morro, A.; McInerney, G.; Murrell, B.; Hartmann, K.T.; Jensen, H.E.; Jungersen, G.; et al. Protection against SARS-CoV-2 transmission by a parenteral prime—Intranasal boost vaccine strategy. EBioMedicine 2022, 84, 104248. [Google Scholar] [CrossRef]
- Wagstaffe, H.R.; Thwaites, R.S.; Reynaldi, A.; Sidhu, J.K.; McKendry, R.; Ascough, S.; Papargyris, L.; Collins, A.M.; Xu, J.; Lemm, N.M. Mucosal and systemic immune correlates of viral control after SARS-CoV-2 infection challenge in seronegative adults. Sci. Immunol. 2024, 9, eadj9285. [Google Scholar] [CrossRef]
- Leñini, C.; Rodriguez Ayala, F.; Goñi, A.J.; Rateni, L.; Nakamura, A.; Grau, R.R. Probiotic properties of Bacillus subtilis DG101 isolated from the traditional Japanese fermented food nattō. Front. Microbiol. 2023, 14, 1253480. [Google Scholar] [CrossRef]
- Fricke, C.; Pfaff, F.; Ulrich, L.; Halwe, N.J.; Schön, J.; Timm, L.; Hoffmann, W.; Rauch, S.; Petsch, B.; Hoffmann, D.; et al. SARS-CoV-2 variants of concern elicit divergent early immune responses in hACE2 transgenic mice. Eur. J. Immunol. 2023, 53, e2250332. [Google Scholar] [CrossRef] [PubMed]
- Ayala, F.; Cardinali, N.; Grau, R. Effectiveness of the probiotic Bacillus subtilis DG101 to treat type 2 diabetes mellitus triggered by SARS-CoV-2 infection. J. Clin. Images Med. Case Rep. 2022, 3, 1847. [Google Scholar] [CrossRef]
- Cardinali, N.; Ayala, F.; Leñini, C.; Perez, O.; Grau, R. Efficacy of The Probiotic Bacillus subtilis DG101 Against Intestinal Discomfort and Constipation in Healthy Adults: A Double-Blind, Placebo-Controlled Study. Am. J. Clin. Med. Res. 2024, 4, 129. [Google Scholar]
- Rosales-Mendoza, S.; Angulo, C. Bacillus subtilis comes of age as a vaccine production host and delivery vehicle. Expert Rev. Vaccines 2015, 14, 1135–1148. [Google Scholar] [PubMed]
- De Souza, R.D.; Batista, M.T.; Luiz, W.B.; Cavalcante, R.C.M.; Amorim, J.H.; Bizerra, R.S.P.; Martins, E.G.; de Souza Ferreira, L.C. Bacillus subtilis spores as vaccine adjuvants: Further insights into the mechanisms of action. PLoS ONE 2014, 9, e87454. [Google Scholar] [CrossRef]
- Amuguni, H.; Tzipori, S. Bacillus subtilis: A temperature resistant and needle free delivery system of immunogens. Hum. Vaccines Immunother. 2012, 8, 979–986. [Google Scholar] [CrossRef]
- Arnaouteli, S.; Bamford, N.C.; Stanley-Wall, N.R.; Kovács, Á.T. Bacillus subtilis biofilm formation and social interactions. Nat. Rev. Microbiol. 2021, 19, 600–614. [Google Scholar] [CrossRef] [PubMed]
- Afzaal, M.; Saeed, F.; Islam, F.; Ateeq, H.; Asghar, A.; Shah, Y.A.; Ofoedu, C.E.; Chacha, J.S. Nutritional health perspective of natto: A critical review. Biochem. Res. Int. 2022, 2022, 5863887. [Google Scholar] [CrossRef] [PubMed]
- Khodavirdipour, A.; Chamanrokh, P.; Alikhani, M.Y.; Alikhani, M.S. Potential of Bacillus subtilis Against SARS-CoV-2—A Sustainable Drug Development Perspective. Front. Microbiol. 2022, 13, 718786. [Google Scholar] [CrossRef]
- Chan, B.C.-L.; Li, P.; Tsang, M.S.-M.; Sung, J.C.-C.; Kwong, K.W.-Y.; Zheng, T.; Hon, S.S.-M.; Lau, C.-P.; Cheng, W.; Chen, F.; et al. Creating a Vaccine-like Supplement against Respiratory Infection Using Recombinant Bacillus subtilis Spores Expressing SARS-CoV-2 Spike Protein with Natural Products. Molecules 2023, 28, 4996. [Google Scholar] [CrossRef]
- Xu, R.; Hong, H.A.; Khandaker, S.; Baltazar, M.; Allehyani, N.; Beentjes, D.; Prince, T.; Ho, Y.-L.; Nguyen, L.H.; Hynes, D.; et al. Nasal delivery of killed Bacillus subtilis spores protects against influenza, RSV and SARS-CoV-2. Front. Immunol. 2025, 16, 1501907. [Google Scholar] [CrossRef]
- Setlow, P. Spores of Bacillus subtilis: Their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Liu, H.; Wu, N.C.; Wilson, I.A. Recognition of the SARS-CoV-2 receptor binding domain by neutralizing antibodies. Biochem. Biophys. Res. Commun. 2021, 538, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Min, L.; Sun, Q. Antibodies and vaccines target RBD of SARS-CoV-2. Front. Mol. Biosci. 2021, 8, 671633. [Google Scholar] [CrossRef]
- Kleanthous, H.; Silverman, J.M.; Makar, K.W.; Yoon, I.K.; Jackson, N.; Vaughn, D.W. Scientific rationale for developing potent RBD-based vaccines targeting COVID-19. NPJ Vaccines 2021, 6, 128. [Google Scholar] [CrossRef] [PubMed]
- Valdes-Balbin, Y.; Santana-Mederos, D.; Quintero, L.; Fernández, S.; Rodriguez, L.; Sanchez Ramirez, B.; Perez-Nicado, R.; Acosta, C.; Méndez, Y.; Ricardo, M.G. SARS-CoV-2 RBD-tetanus toxoid conjugate vaccine induces a strong neutralizing immunity in preclinical studies. ACS Chem. Biol. 2021, 16, 1223–1233. [Google Scholar] [CrossRef]
- Zang, J.; Zhu, Y.; Zhou, Y.; Gu, C.; Yi, Y.; Wang, S.; Xu, S.; Hu, G.; Du, S.; Yin, Y. Yeast-produced RBD-based recombinant protein vaccines elicit broadly neutralizing antibodies and durable protective immunity against SARS-CoV-2 infection. Cell Discov. 2021, 7, 71. [Google Scholar] [CrossRef]
- Dashti, N.; Golsaz-Shirazi, F.; Soltanghoraee, H.; Zarnani, A.-H.; Mohammadi, M.; Imani, D.; Jeddi-Tehrani, M.; Amiri, M.M.; Shokri, F. Preclinical assessment of a recombinant RBD-Fc fusion protein as SARS-CoV-2 candidate vaccine. Eur. J. Microbiol. Immunol. 2024, 14, 228–242. [Google Scholar] [CrossRef]
- Pollet, J.; Strych, U.; Chen, W.H.; Versteeg, L.; Keegan, B.; Zhan, B.; Wei, J.; Liu, Z.; Lee, J.; Kundu, R. Receptor-binding domain recombinant protein on alum-CpG induces broad protection against SARS-CoV-2 variants of concern. Vaccine 2022, 40, 3655–3663. [Google Scholar] [CrossRef]
- Hernández-Bernal, F.; Ricardo-Cobas, M.C.; Martín-Bauta, Y.; Rodríguez-Martínez, E.; Urrutia-Pérez, K.; Urrutia-Pérez, K.; Quintana-Guerra, J.; Navarro-Rodríguez, Z.; Piñera-Martínez, M.; Rodríguez-Reinoso, J.L. A phase 3, randomised, double-blind, placebo-controlled clinical trial evaluation of the efficacy and safety of a SARS-CoV-2 recombinant spike RBD protein vaccine in adults (ABDALA-3 study). Lancet Reg. Health–Am. 2023, 21, 100497. [Google Scholar] [CrossRef]
- Law, J.L.M.; Logan, M.; Joyce, M.A.; Landi, A.; Hockman, D.; Crawford, K.; Johnson, J.; LaChance, G.; Saffran, H.A.; Shields, J. SARS-COV-2 recombinant Receptor-Binding-Domain (RBD) induces neutralizing antibodies against variant strains of SARS-CoV-2 and SARS-CoV-1. Vaccine 2021, 39, 5769–5779. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.-I.; Chung, M.-S. Bacillus spores: A review of their properties and inactivation processing technologies. Food Sci. Biotechnol. 2020, 29, 1447–1461. [Google Scholar] [CrossRef] [PubMed]
- Ahimou, F.; Paquot, M.; Jacques, P.; Thonart, P.; Rouxhet, P.G. Influence of electrical properties on the evaluation of the surface hydrophobicity of Bacillus subtilis. J. Microbiol. Methods 2001, 45, 119–126. [Google Scholar] [CrossRef]
- Boggiano-Ayo, T.; Palacios-Oliva, J.; Lozada-Chang, S.; Relova-Hernandez, E.; Gomez-Perez, J.; Oliva, G.; Hernandez, L.; Bueno-Soler, A.; Montes de Oca, D.; Mora, O. Development of a scalable single process for producing SARS-CoV-2 RBD monomer and dimer vaccine antigens. Front. Bioeng. Biotechnol. 2023, 11, 1287551. [Google Scholar] [CrossRef]
- Kehagia, E.; Papakyriakopoulou, P.; Valsami, G. Advances in intranasal vaccine delivery: A promising non-invasive route of immunization. Vaccine 2023, 41, 3589–3603. [Google Scholar] [CrossRef]
- Ramvikas, M.; Arumugam, M.; Chakrabarti, S.R.; Jaganathan, K.S. Nasal Vaccine Delivery. In Micro- and Nanotechnology in Vaccine Development; Elsevier: Amsterdam, The Netherlands, 2017; pp. 279–301. [Google Scholar]
- Tub-Chafer, F.; Reyes-Díaz, L.M.; Vega-García, I.G.; González-Aznar, E.; Otero-Alfaro, O.; Lumpuy-Castillo, J.; Ricardo-Grau, R.; Pérez, O. Acción adyuvante de esporas de Bacillus subtilis por vía mucosa. VacciMonitor 2016, 25, 19–29. [Google Scholar]
- Huang, J.-M.; Hong, H.A.; Van Tong, H.; Hoang, T.H.; Brisson, A.; Cutting, S.M. Mucosal delivery of antigens using adsorption to bacterial spores. Vaccine 2009, 28, 1021–1030. [Google Scholar] [CrossRef]
- Song, M.; Hong, H.A.; Huang, J.-M.; Colenutt, C.; Khang, D.D.; Van Anh Nguyen, T.; Park, S.-M.; Shim, B.-S.; Song, H.H.; Cheon, I.S.; et al. Killed Bacillus subtilis spores as a mucosal adjuvant for an H5N1 vaccine. Vaccine 2012, 30, 3266–3277. [Google Scholar] [CrossRef]
- Ricca, E.; Baccigalupi, L.; Cangiano, G.; De Felice, M.; Isticato, R. Mucosal vaccine delivery by non-recombinant spores of Bacillus subtilis. Microb. Cell Factories 2014, 13, 115. [Google Scholar] [CrossRef]
- Isticato, R.; Ricca, E.; Baccigalupi, L. Spore adsorption as a nonrecombinant display system for enzymes and antigens. J. Vis. Exp. 2019, e59102. [Google Scholar] [CrossRef]
- Ricca, E.; Baccigalupi, L.; Isticato, R. Spore-adsorption: Mechanism and applications of a non-recombinant display system. Biotechnol. Adv. 2020, 47, 107693. [Google Scholar] [CrossRef] [PubMed]
- George, A.J.; Harmsen, B.J.; Ford, J.A.; Tadepalli, S.R.; Horton, N.D. Evaluation of submental blood collection in mice (Mus musculus). J. Am. Assoc. Lab. Anim. Sci. 2023, 62, 92–98. [Google Scholar] [CrossRef]
- Parasuraman, S.; Raveendran, R. Biological sample collection from experimental animals. In Introduction to Basics of Pharmacology and Toxicology: Volume 3: Experimental Pharmacology: Research Methodology and Biostatistics; Springer: Berlin/Heidelberg, Germany, 2022; pp. 45–63. [Google Scholar]
- Henderson, M. Transcardial Perfusion in Mouse; protocols.io; Springer: Berlin/Heidelberg, Germany, 2023. [Google Scholar]
- Sam-Yellowe, T.Y.; Sam-Yellowe, T.Y. Exercise 4: Single Suspension of Mouse Spleen Cells, Cell Viability Assays and Identification of Specific Cells Using Cell Surface Antigens. In Immunology: Overview and Laboratory Manual; Springer: Berlin/Heidelberg, Germany, 2021; pp. 255–265. [Google Scholar]
- Vanlandewijck, M.; Andrae, J.; Gouveia, L.; Betsholtz, C. Preparation of single cell suspensions from the adult mouse lung. Res. Sq. 2018. [Google Scholar] [CrossRef]
- Daubeuf, F.; Frossard, N. Performing bronchoalveolar lavage in the mouse. Curr. Protoc. Mouse Biol. 2012, 2, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Cisney, E.D.; Fernandez, S.; Hall, S.I.; Krietz, G.A.; Ulrich, R.G. Examining the role of nasopharyngeal-associated lymphoreticular tissue (NALT) in mouse responses to vaccines. J. Vis. Exp. JoVE 2012, 3960. [Google Scholar] [CrossRef]
- Oluka, G.K.; Namubiru, P.; Kato, L.; Ankunda, V.; Gombe, B.; Cotten, M.; Team, C.I.; Musenero, M.; Kaleebu, P.; Fox, J. Optimisation and Validation of a conventional ELISA and cut-offs for detecting and quantifying anti-SARS-CoV-2 Spike, RBD, and Nucleoprotein IgG, IgM, and IgA antibodies in Uganda. Front. Immunol. 2023, 14, 1113194. [Google Scholar] [CrossRef]
- Santana-Mederos, D.; Perez-Nicado, R.; Climent, Y.; Rodriguez, L.; Ramirez, B.S.; Perez-Rodriguez, S.; Rodriguez, M.; Labrada, C.; Hernandez, T.; Diaz, M.; et al. A COVID-19 vaccine candidate composed of the SARS-CoV-2 RBD dimer and Neisseria meningitidis outer membrane vesicles. RSC Chem. Biol. 2021, 3, 242–249. [Google Scholar] [CrossRef]
- Manuylov, V.; Dolzhikova, I.; Kudryashova, A.; Cherepovich, B.; Kovyrshina, A.; Iliukhina, A.; Kharchenko, O.; Semashko, M.; Tkachuk, A.; Gushchin, V. Simple ELISA methods to estimate neutralizing antibody Titers to SARS-CoV-2: IgG quantification, the avidity index, and the surrogate virus neutralization test. Arch. Microbiol. Immunol. 2022, 6, 213–220. [Google Scholar] [CrossRef]
- Pi-Estopiñan, F.; Pérez, M.T.; Fraga, A.; Bergado, G.; Díaz, G.D.; Orosa, I.; Díaz, M.; Solozábal, J.A.; Rodríguez, L.M.; Garcia-Rivera, D. A cell-based ELISA as surrogate of virus neutralization assay for RBD SARS-CoV-2 specific antibodies. Vaccine 2022, 40, 1958–1967. [Google Scholar] [CrossRef]
- Muul, L.M.; Heine, G.; Silvin, C.; James, S.P.; Candotti, F.; Radbruch, A.; Worm, M. Measurement of proliferative responses of cultured lymphocytes. Curr. Protoc. Immunol. 2011, 94, 7.10.1–7.10.26. [Google Scholar] [CrossRef]
- Foster, B.; Prussin, C.; Liu, F.; Whitmire, J.K.; Whitton, J.L. Detection of intracellular cytokines by flow cytometry. Curr. Protoc. Immunol. 2007, 78, 6.24.1–6.24.21. [Google Scholar] [CrossRef] [PubMed]
- Rybkina, K.; Davis-Porada, J.; Farber, D.L. Tissue immunity to SARS-CoV-2: Role in protection and immunopathology. Immunol. Rev. 2022, 309, 25–39. [Google Scholar] [CrossRef]
- Allie, S.R.; Bradley, J.E.; Mudunuru, U.; Schultz, M.D.; Graf, B.A.; Lund, F.E.; Randall, T.D. The establishment of resident memory B cells in the lung requires local antigen encounter. Nat. Immunol. 2019, 20, 97–108. [Google Scholar] [CrossRef]
- Hassan, A.O.; Case, J.B.; Winkler, E.S.; Thackray, L.B.; Kafai, N.M.; Bailey, A.L.; McCune, B.T.; Fox, J.M.; Chen, R.E.; Alsoussi, W.B. A SARS-CoV-2 infection model in mice demonstrates protection by neutralizing antibodies. Cell 2020, 182, 744–753.e4. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.F.; Zhao, F.; Huang, D.; Beutler, N.; Burns, A.; He, W.; Limbo, O.; Smith, C.; Song, G.; Woehl, J. Isolation of potent SARS-CoV-2 neutralizing antibodies and protection from disease in a small animal model. Science 2020, 369, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Trichel, A.M. Overview of Nonhuman Primate Models of SARS-CoV-2 Infection. Comp. Med. 2021, 71, 411–432. [Google Scholar] [CrossRef]
- Chen, Z.; Yuan, Y.; Hu, Q.; Zhu, A.; Chen, F.; Li, S.; Guan, X.; Lv, C.; Tang, T.; He, Y. SARS-CoV-2 immunity in animal models. Cell. Mol. Immunol. 2024, 21, 119–133. [Google Scholar] [CrossRef]
- Koch, T.; Mellinghoff, S.C.; Shamsrizi, P.; Addo, M.M.; Dahlke, C. Correlates of vaccine-induced protection against SARS-CoV-2. Vaccines 2021, 9, 238. [Google Scholar] [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Cromer, D.; Steain, M.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Kent, S.J.; Triccas, J.A.; Khoury, D.S.; Davenport, M.P. Neutralising antibody titres as predictors of protection against SARS-CoV-2 variants and the impact of boosting: A meta-analysis. Lancet Microbe 2022, 3, e52–e61. [Google Scholar] [CrossRef]
- Pilapitiya, D.; Wheatley, A.K.; Tan, H.X. Mucosal vaccines for SARS-CoV-2: Triumph of hope over experience. EBioMedicine 2023, 92, 104585. [Google Scholar] [CrossRef]
- Yahyaei, S.; Abdoli, A.; Jamali, A.; Teimoori, A.; Arefian, E.; Eftekhari, Z.; Jamur, P. Targeting respiratory viruses: The efficacy of intranasal mRNA vaccination in generating protective mucosal and systemic immunity against influenza A (H1N1). Influenza Other Respir. Viruses 2025, 19, e70093. [Google Scholar] [CrossRef]
- Anthi, A.K.; Kolderup, A.; Vaage, E.B.; Bern, M.; Benjakul, S.; Tjärnhage, E.; Ruso-Julve, F.; Jensen, K.-R.; Lode, H.E.; Vaysburd, M.; et al. An intranasal subunit vaccine induces protective systemic and mucosal antibody immunity against respiratory viruses in mouse models. Nat. Commun. 2025, 16, 3999. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.O.; Shrihari, S.; Gorman, M.J.; Ying, B.; Yuan, D.; Raju, S.; Chen, R.E.; Dmitriev, I.P.; Kashentseva, E.; Adams, L.J.; et al. An intranasal vaccine durably protects against SARS-CoV-2 variants in mice. Cell Rep. 2021, 36, 109452. [Google Scholar] [CrossRef]
- Gallo, O.; Locatello, L.G.; Mazzoni, A.; Novelli, L.; Annunziato, F. The central role of the nasal microenvironment in the transmission, modulation, and clinical progression of SARS-CoV-2 infection. Mucosal Immunol. 2020, 14, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Mahla, R.S.; Reddy, M.C.; Prasad, D.V.R.; Kumar, H. Sweeten PAMPs: Role of sugar complexed PAMPs in innate immunity and vaccine biology. Front. Immunol. 2013, 4, 248. [Google Scholar] [CrossRef] [PubMed]
- Sterlin, D.; Mathian, A.; Miyara, M.; Mohr, A.; Anna, F.; Claër, L.; Quentric, P.; Fadlallah, J.; Devilliers, H.; Ghillani, P. IgA dominates the early neutralizing antibody response to SARS-CoV-2. Sci. Transl. Med. 2021, 13, eabd2223. [Google Scholar] [CrossRef]
- Kurano, M.; Morita, Y.; Nakano, Y.; Yokoyama, R.; Shimura, T.; Qian, C.; Xia, F.; He, F.; Zheng, L.; Ohmiya, H. Response kinetics of different classes of antibodies to SARS-CoV2 infection in the Japanese population: The IgA and IgG titers increased earlier than the IgM titers. Int. Immunopharmacol. 2022, 103, 108491. [Google Scholar] [CrossRef]
- Takamatsu, Y.; Omata, K.; Shimizu, Y.; Kinoshita-Iwamoto, N.; Terada, M.; Suzuki, T.; Morioka, S.; Uemura, Y.; Ohmagari, N.; Maeda, K. SARS-CoV-2-neutralizing humoral IgA response occurs earlier but is modest and diminishes faster than IgG response. Microbiol. Spectr. 2022, 10, e02716-22. [Google Scholar] [CrossRef]
- Stacey, H.D.; Golubeva, D.; Posca, A.; Ang, J.C.; Novakowski, K.E.; Zahoor, M.A.; Kaushic, C.; Cairns, E.; Bowdish, D.M.; Mullarkey, C.E. IgA potentiates NETosis in response to viral infection. Proc. Natl. Acad. Sci. USA 2021, 118, e2101497118. [Google Scholar] [CrossRef]
- Yaugel-Novoa, M.; Noailly, B.; Jospin, F.; Pizzorno, A.; Traversier, A.; Pozzetto, B.; Waeckel, L.; Longet, S.; Pillet, S.; Botelho-Nevers, E. Impaired mucosal IgA response in patients with severe COVID-19. Emerg. Microbes Infect. 2024, 13, 2401940. [Google Scholar] [CrossRef] [PubMed]
- Yaugel-Novoa, M.; Bourlet, T.; Paul, S. Role of the humoral immune response during COVID-19: Guilty or not guilty? Mucosal Immunol. 2022, 15, 1170–1180. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, M.; Mirabella, L.; Cotoia, A.; Faleo, G.; Rauseo, M.; Rizzo, A.C.; Fiore, J.R.; Cinnella, G.; Serviddio, G. A Possible Protective Effect of IgA Against Severe Acute Respiratory Syndrome Corona Virus 2 (SARS-CoV-2) in Bronchoalveolar Lavage in COVID-19 Patients Admitted to Intensive Care Unit. Viruses 2024, 16, 1851. [Google Scholar] [CrossRef]
- Tyagi, R.; Basu, S.; Dhar, A.; Gupta, S.; Gupta, S.L.; Jaiswal, R.K. Role of Immunoglobulin A in COVID-19 and Influenza Infections. Vaccines 2023, 11, 1647. [Google Scholar] [CrossRef]
- Noh, H.E.; Rha, M.S. Mucosal Immunity against SARS-CoV-2 in the Respiratory Tract. Pathogens 2024, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.; Oh, J.E. Resident memory B cells in barrier tissues. Front. Immunol. 2022, 13, 953088. [Google Scholar] [CrossRef]
- Longet, S.; Paul, S. Pivotal role of tissue-resident memory lymphocytes in the control of mucosal infections: Can mucosal vaccination induce protective tissue-resident memory T and B cells? Front. Immunol. 2023, 14, 1216402. [Google Scholar] [CrossRef]
- Poon, M.M.; Rybkina, K.; Kato, Y.; Kubota, M.; Matsumoto, R.; Bloom, N.I.; Zhang, Z.; Hastie, K.M.; Grifoni, A.; Weiskopf, D. SARS-CoV-2 infection generates tissue-localized immunological memory in humans. Sci. Immunol. 2021, 6, eabl9105. [Google Scholar] [CrossRef]
- Rha, M.S.; Kim, A.R.; Shin, E.C. SARS-CoV-2-specific T cell responses in patients with COVID-19 and unexposed individuals. Immune Netw. 2021, 21, e2. [Google Scholar] [CrossRef]
- Pavel, A.B.; Glickman, J.W.; Michels, J.R.; Kim-Schulze, S.; Miller, R.L.; Guttman-Yassky, E. Th2/Th1 cytokine imbalance is associated with higher COVID-19 risk mortality. Front. Genet. 2021, 12, 706902. [Google Scholar] [CrossRef]
- Yang, Y.; Miller, H.; Byazrova, M.G.; Cndotti, F.; Benlagha, K.; Camara, N.O.S.; Shi, J.; Forsman, H.; Lee, P.; Yang, L. The characterization of CD8+ T-cell responses in COVID-19. Emerg. Microbes Infect. 2024, 13, 2287118. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, J.S.; Bilich, T.; Tandler, C.; Nelde, A.; Maringer, Y.; Marconato, M.; Reusch, J.; Jäger, S.; Denk, M.; Richter, M. A COVID-19 peptide vaccine for the induction of SARS-CoV-2 T cell immunity. Nature 2022, 601, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Gartlan, C.; Tipton, T.; Salguero, F.J.; Sattentau, Q.; Gorringe, A.; Carroll, M.W. Vaccine-associated enhanced disease and pathogenic human coronaviruses. Front. Immunol. 2022, 13, 882972. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.Z.; Wakim, L.M. Tissue resident memory T cells in the respiratory tract. Mucosal Immunol. 2022, 15, 379–388. [Google Scholar] [CrossRef]
- Cheon, I.S.; Son, Y.M.; Sun, J. Tissue-resident memory T cells and lung immunopathology. Immunol. Rev. 2023, 316, 63–83. [Google Scholar] [CrossRef]
- Carbone, F.R. Unique properties of tissue-resident memory T cells in the lungs: Implications for COVID-19 and other respiratory diseases. Nat. Rev. Immunol. 2023, 23, 329–335. [Google Scholar] [CrossRef]
- Mitsi, E.; Diniz, M.O.; Reiné, J.; Collins, A.M.; Robinson, R.E.; Hyder-Wright, A.; Farrar, M.; Liatsikos, K.; Hamilton, J.; Onyema, O. Respiratory mucosal immune memory to SARS-CoV-2 after infection and vaccination. Nat. Commun. 2023, 14, 6815. [Google Scholar] [CrossRef]
- Isticato, R. Bacterial spore-based delivery system: 20 years of a versatile approach for innovative vaccines. Biomolecules 2023, 13, 947. [Google Scholar] [CrossRef]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef]
- Lee, S.; Belitsky, B.R.; Brinker, J.P.; Kerstein, K.O.; Brown, D.W.; Clements, J.D.; Keusch, G.T.; Tzipori, S.; Sonenshein, A.L.; Herrmann, J.E. Development of a Bacillus subtilis-based rotavirus vaccine. Clin. Vaccine Immunol. 2010, 17, 1647–1655. [Google Scholar] [CrossRef]
- Hong, H.A.; Khaneja, R.; Tam, N.M.; Cazzato, A.; Tan, S.; Urdaci, M.; Brisson, A.; Gasbarrini, A.; Barnes, I.; Cutting, S.M. Bacillus subtilis isolated from the human gastrointestinal tract. Res. Microbiol. 2009, 160, 134–143. [Google Scholar] [CrossRef]
- Huang, J.M.; La Ragione, R.M.; Nunez, A.; Cutting, S.M. Immunostimulatory activity of Bacillus spores. FEMS Immunol. Med. Microbiol. 2008, 53, 195–203. [Google Scholar] [CrossRef]
- Zhou, M.; Xiao, H.; Yang, X.; Cheng, T.; Yuan, L.; Xia, N. Novel vaccine strategies to induce respiratory mucosal immunity: Advances and implications. MedComm 2025, 6, e70056. [Google Scholar] [CrossRef] [PubMed]
- So, Y.J.; Park, O.-J.; Kwon, Y.; Im, J.; Lee, D.; Yun, S.-H.; Cho, K.; Yun, C.-H.; Han, S.H. Bacillus subtilis Induces Human Beta Defensin-2 Through its Lipoproteins in Human Intestinal Epithelial Cells. Probiotics Antimicrob. Proteins 2024, 17, 1648–1662. [Google Scholar] [CrossRef] [PubMed]
- Saggese, A.; Baccigalupi, L.; Donadio, G.; Ricca, E.; Isticato, R. The bacterial spore as a mucosal vaccine delivery system. Int. J. Mol. Sci. 2023, 24, 10880. [Google Scholar] [CrossRef] [PubMed]
- Roos, T.B.; de Moraes, C.M.; Sturbelle, R.T.; Dummer, L.A.; Fischer, G.; Leite, F.P.L. Probiotics Bacillus toyonensis and Saccharomyces boulardii improve the vaccine immune response to Bovine herpesvirus type 5 in sheep. Res. Vet. Sci. 2018, 117, 260–265. [Google Scholar] [CrossRef]
- Santos, F.D.S.; Maubrigades, L.R.; Gonçalves, V.S.; Ferreira, M.R.A.; Brasil, C.L.; Cunha, R.C.; Conceição, F.R.; Leite, F.P.L. Immunomodulatory effect of short-term supplementation with Bacillus toyonensis BCT-7112T and Saccharomyces boulardii CNCM I-745 in sheep vaccinated with Clostridium chauvoei. Vet. Immunol. Immunopathol. 2021, 237, 110272. [Google Scholar] [CrossRef]
- Sung, J.C.-C.; Lai, N.C.-Y.; Wu, K.-C.; Choi, M.-C.; Ho-Yi, C.M.A.; Lin, J.; Kuok, C.-N.; Leong, W.-L.; Lam, W.-K.; Hamied, Y.K.; et al. Safety and Immunogenicity of Inactivated Bacillus subtilis Spores as a Heterologous Antibody Booster for COVID-19 Vaccines. Vaccines 2022, 10, 1014. [Google Scholar] [CrossRef]
- Lei, H.; Alu, A.; Yang, J.; Ren, W.; He, C.; Lan, T.; He, X.; Yang, L.; Li, J.; Wang, Z.; et al. Intranasal administration of a recombinant RBD vaccine induces long-term immunity against Omicron-included SARS-CoV-2 variants. Signal Transduct. Target. Ther. 2022, 7, 159. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos Pupo, R.; Reyes Diaz, L.M.; Suarez Formigo, G.M.; Borrego Gonzalez, Y.; Lastre Gonzalez, M.; Saavedra Hernandez, D.; Crombet Ramos, T.; Sanchez Ramirez, B.; Grau, R.; Hellings, N.; et al. Mucosal Vaccination Against SARS-CoV-2 Using Human Probiotic Bacillus subtilis Spores as an Adjuvant Induces Potent Systemic and Mucosal Immunity. Vaccines 2025, 13, 772. https://doi.org/10.3390/vaccines13070772
Ramos Pupo R, Reyes Diaz LM, Suarez Formigo GM, Borrego Gonzalez Y, Lastre Gonzalez M, Saavedra Hernandez D, Crombet Ramos T, Sanchez Ramirez B, Grau R, Hellings N, et al. Mucosal Vaccination Against SARS-CoV-2 Using Human Probiotic Bacillus subtilis Spores as an Adjuvant Induces Potent Systemic and Mucosal Immunity. Vaccines. 2025; 13(7):772. https://doi.org/10.3390/vaccines13070772
Chicago/Turabian StyleRamos Pupo, Raul, Laura M. Reyes Diaz, Gisela M. Suarez Formigo, Yusnaby Borrego Gonzalez, Miriam Lastre Gonzalez, Danay Saavedra Hernandez, Tania Crombet Ramos, Belinda Sanchez Ramirez, Roberto Grau, Niels Hellings, and et al. 2025. "Mucosal Vaccination Against SARS-CoV-2 Using Human Probiotic Bacillus subtilis Spores as an Adjuvant Induces Potent Systemic and Mucosal Immunity" Vaccines 13, no. 7: 772. https://doi.org/10.3390/vaccines13070772
APA StyleRamos Pupo, R., Reyes Diaz, L. M., Suarez Formigo, G. M., Borrego Gonzalez, Y., Lastre Gonzalez, M., Saavedra Hernandez, D., Crombet Ramos, T., Sanchez Ramirez, B., Grau, R., Hellings, N., Stinissen, P., Perez, O., & Bogie, J. F. J. (2025). Mucosal Vaccination Against SARS-CoV-2 Using Human Probiotic Bacillus subtilis Spores as an Adjuvant Induces Potent Systemic and Mucosal Immunity. Vaccines, 13(7), 772. https://doi.org/10.3390/vaccines13070772