

Next-Generation Vaccine Platforms: Integrating Synthetic Biology, Nanotechnology, and Systems Immunology for Improved Immunogenicity
 , ,
, , 
 and
and
Abstract
1. The Evolution of Vaccines: From Traditional to Next-Generation Platforms
2. Synthetic Biology in Vaccine Design
2.1. Integrated Perspective on Vaccine Adjuvants: Common Mechanisms and Strategic Insights
2.2. Key Functional Characteristics of Vaccine Adjuvants
3. Nanoparticle Properties and Their Immunological Impacts
3.1. Size
3.2. Shape
3.3. Surface Charge
3.4. Composition
3.5. Molecular and Cellular Mechanisms
4. Nanotechnology as a Vaccine Delivery System
5. Systems Immunology: Decoding Immune Responses
6. Synergistic Integration of the Three Pillars

{kind=link}
{kind=link}
{kind=link}
| Platform Type | Product/Trial Name | Technology Used | Target Disease | Clinical Phase/Status |
|---|---|---|---|---|
| mRNA-based (LNP) | BNT162b2 (Pfizer–BioNTech) | Synthetic mRNA + lipid nanoparticles | COVID-19 | Approved (EMA, FDA) |
| saRNA | ARCT-154 (Arcturus) | Self-amplifying RNA + LNP | COVID-19 | Phase 3 |
| DNA Vaccine | INO-4800 | Synthetic plasmid DNA | COVID-19 | Phase 3 |
| Nanoparticle Protein-based | NVX-CoV2373 (Novavax) | Recombinant protein + Matrix-M (saponin NP) | COVID-19 | Approved (e.g., WHO, EU) |
| VLP-based | Mosquirix (RTS,S/AS01) | Hepatitis B-based VLP + AS01 adjuvant | Malaria | Approved (WHO, 2021) |
| Personalized Neoantigen | mRNA-4157/V940 + Keytruda | mRNA encoding patient-specific tumor neoantigens | Melanoma (Cancer) | Phase 2 (Positive results) |
7. Integrated Conceptual Framework: Synergistic Interaction of Synthetic Biology, Nanotechnology, and Systems Immunology
8. Challenges and Ethical Considerations
8.1. Regulatory Challenges
8.2. Manufacturing Scalability and Complexity
8.3. Cost Implications and Global Access
8.4. Case Studies on Regulatory Approval
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Glossary of Key Technical Terms
| Term | Definition |
| Codon Optimization | The modification of gene codons to improve the efficiency of protein expression in the target host organism. |
| Virus-like Particles (VLPs) | VLPs are structures that resemble viruses but lack genetic material, making them a safe and effective tool for triggering immune responses to vaccines. |
| Self-amplifying RNA (saRNA) | A synthetic RNA platform that replicates within host cells, producing higher antigen levels from lower vaccine doses. |
| Pattern Recognition Receptors (PRRs) | Cellular receptors that recognize pathogen-associated molecular patterns (PAMPs) to trigger immune responses. |
| Multi-omics | An integrative approach combining datasets from various “omics” fields, such as genomics, transcriptomics, and proteomics. |
| Codon Usage Bias | The preferential usage of certain synonymous codons over others in a gene, which can affect protein translation efficiency. |
| Nanovaccine | A vaccine platform that utilizes nanoparticles to deliver antigens directly and efficiently to immune cells. |
| Adjuvant | A substance added to vaccines to enhance the body’s immune response to the presented antigen. |
| Transcriptomics | The comprehensive analysis of all RNA transcripts in a cell or tissue used to study gene activity and regulation. |
| Immunopeptidomics | A proteomics subfield that is focused on identifying the peptides presented by MHC molecules to inform precise vaccine design. |
References
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Plotkin, S. History of vaccination. Proc. Natl. Acad. Sci. USA 2014, 111, 12283–12287. [Google Scholar] [CrossRef]
- Tahamtan, A.; Charostad, J.; Hoseini Shokouh, S.J.; Barati, M. An overview of history, evolution, and manufacturing of various generations of vaccines. J. Arch. Mil. Med. 2017, 5, e12315. [Google Scholar] [CrossRef]
- Levine, M.M.; Lagos, R. Vaccines and vaccination in historical perspective. In New Generation Vaccines; CRC Press: Boca Raton, FL, USA, 2016; pp. 29–39. [Google Scholar]
- Sandle, T. The Manufacture and Quality Control of Immunological Products. In Hugo and Russell’s Pharmaceutical Microbiology; John Wiley & Sons Ltd.: London, UK, 2023. [Google Scholar]
- Kozak, M.; Hu, J. The Integrated Consideration of Vaccine Platforms, Adjuvants, and Delivery Routes for Successful Vaccine Development. Vaccines 2023, 11, 695. [Google Scholar] [CrossRef]
- Méthot, P.-O. Bacterial transformation and the origins of epidemics in the interwar period: The epidemiological significance of Fred Griffith’s “transforming experiment”. J. Hist. Biol. 2016, 49, 311–358. [Google Scholar] [CrossRef]
- Hicks, D.; Fooks, A.; Johnson, N. Developments in rabies vaccines. Clin. Exp. Immunol. 2012, 169, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Bröker, M.; Berti, F.; Schneider, J.; Vojtek, I. Polysaccharide conjugate vaccine protein carriers as a “neglected valency”–potential and limitations. Vaccine 2017, 35, 3286–3294. [Google Scholar] [CrossRef] [PubMed]
- Moyle, P.M.; Toth, I. Modern subunit vaccines: Development, components, and research opportunities. ChemMedChem 2013, 8, 360–376. [Google Scholar] [CrossRef]
- Eslami, M.; Arjmand, N.; Mahmoudian, F.; Babaeizad, A.; Tahmasebi, H.; Fattahi, F.; Oksenych, V. Deciphering Host–Virus Interactions and Advancing Therapeutics for Chronic Viral Infection. Viruses 2025, 17, 390. [Google Scholar] [CrossRef]
- Romano’, L.; Zanetti, A.R. Hepatitis B vaccination: A historical overview with a focus on the Italian achievements. Viruses 2022, 14, 1515. [Google Scholar] [CrossRef]
- Hudu, S.A.; Shinkafi, S.H.; Umar, S. An overview of recombinant vaccine technology, adjuvants and vaccine delivery methods. Int. J. Pharm. Pharm. Sci. 2016, 8, 19–24. [Google Scholar] [CrossRef]
- Harper, D.M.; DeMars, L.R. HPV vaccines–a review of the first decade. Gynecol. Oncol. 2017, 146, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Anwari, P. Benefit-Risk and Cost-Utility of Rotavirus Vaccination in Afghanistan: A Modelling Study Informed by Post-Marketing Surveillance Data. Ph.D. Thesis, London School of Hygiene & Tropical Medicine, London, UK, 2025. [Google Scholar]
- Alizon, S.; Michalakis, Y. Adaptive virulence evolution: The good old fitness-based approach. Trends Ecol. Evol. 2015, 30, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S.A.C.; Clemens, R. The need and challenges for development of vaccines against emerging infectious diseases. J. Pediatria 2023, 99, S37–S45. [Google Scholar] [CrossRef]
- Lozano, D.; Larraga, V.; Vallet-Regí, M.; Manzano, M. An Overview of the Use of Nanoparticles in Vaccine Development. Nanomaterials 2023, 13, 1828. [Google Scholar] [CrossRef]
- Kanekiyo, M.; Ellis, D.; King, N.P. New Vaccine Design and Delivery Technologies. J. Infect. Dis. 2019, 219, S88–S96. [Google Scholar] [CrossRef]
- Zhang, R.; Ulery, B. Synthetic Vaccine Characterization and Design. J. Bionanosci. 2018, 12, 1–11. [Google Scholar] [CrossRef]
- Jones, L.H. Recent advances in the molecular design of synthetic vaccines. Nat. Chem. 2015, 7, 952–960. [Google Scholar] [CrossRef]
- Gary, E.N.; Weiner, D.B. DNA vaccines: Prime time is now. Curr. Opin. Immunol. 2020, 65, 21–27. [Google Scholar] [CrossRef]
- Rauch, S.; Jasny, E.; Schmidt, K.E.; Petsch, B. New Vaccine Technologies to Combat Outbreak Situations. Front. Immunol. 2018, 9, 1963. [Google Scholar] [CrossRef]
- Uddin, M.N.; Roni, M.A. Challenges of Storage and Stability of mRNA-Based COVID-19 Vaccines. Vaccines 2021, 9, 1033. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Kwon, M.; Im, S.; Lee, K.; Lee, H. mRNA vaccines: The most recent clinical applications of synthetic mRNA. Arch. Pharm. Res. 2022, 45, 245–262. [Google Scholar] [CrossRef] [PubMed]
- Amanat, F.; Krammer, F. SARS-CoV-2 Vaccines: Status Report. Immunity 2020, 52, 583–589. [Google Scholar] [CrossRef]
- Saade, F.; Petrovsky, N. Technologies for enhanced efficacy of DNA vaccines. Expert. Rev. Vaccines 2012, 11, 189–209. [Google Scholar] [CrossRef]
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-Based Vaccines: Current Progress and Future Challenges. Chem. Rev. 2020, 120, 3210–3229. [Google Scholar] [CrossRef]
- Han, J.H.; Choi, Y.S.; Kim, W.J.; Jeon, Y.H.; Lee, S.K.; Lee, B.J.; Ryu, K.S. Codon optimization enhances protein expression of human peptide deformylase in E. coli. Protein Expr. Purif. 2010, 70, 224–230. [Google Scholar] [CrossRef]
- Liu, B.; Kong, Q.; Zhang, D.; Yan, L. Codon optimization significantly enhanced the expression of human 37-kDa iLRP in Escherichia coli. 3 Biotech 2018, 8, 210. [Google Scholar] [CrossRef]
- Hyatt, D.; LoCascio, P.F.; Hauser, L.J.; Uberbacher, E.C. Gene and translation initiation site prediction in metagenomic sequences. Bioinformatics 2012, 28, 2223–2230. [Google Scholar] [CrossRef] [PubMed]
- Schlub, T.E.; Buchmann, J.P.; Holmes, E.C. A Simple Method to Detect Candidate Overlapping Genes in Viruses Using Single Genome Sequences. Mol. Biol. Evol. 2018, 35, 2572–2581. [Google Scholar] [CrossRef]
- Codagenix, Inc. Codagenix Announces the Synthesis and Preliminary Safety of Scalable Live-Attenuated Vaccine Candidate Against COVID-19. Available online: https://www.prnewswire.com/news-releases/codagenix-announces-the-synthesis-and-preliminary-safety-of-scalable-live-attenuated-vaccine-candidate-against-covid-19-301079306.html (accessed on 18 June 2020).
- Pulendran, B.; Arunachalam, P.S.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef]
- Zhao, T.; Cai, Y.; Jiang, Y.; He, X.; Wei, Y.; Yu, Y.; Tian, X. Vaccine adjuvants: Mechanisms and platforms. Signal Transduct. Target. Ther. 2023, 8, 283. [Google Scholar] [CrossRef] [PubMed]
- Cantisani, R.; Pezzicoli, A.; Cioncada, R.; Malzone, C.; De Gregorio, E.; D’Oro, U.; Piccioli, D. Vaccine adjuvant MF59 promotes retention of unprocessed antigen in lymph node macrophage compartments and follicular dendritic cells. J. Immunol. 2015, 194, 1717–1725. [Google Scholar] [CrossRef]
- O’Hagan, D.T.; Ott, G.S.; De Gregorio, E.; Seubert, A. The mechanism of action of MF59—An innately attractive adjuvant formulation. Vaccine 2012, 30, 4341–4348. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Omer, S.B. Why and How Vaccines Work. Cell 2020, 183, 290–295. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; Valiante, N.M. Recent advances in the discovery and delivery of vaccine adjuvants. Nat. Rev. Drug Discov. 2003, 2, 727–735. [Google Scholar] [CrossRef]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Schmidt, C.; Schnierle, B.S. Self-Amplifying RNA Vaccine Candidates: Alternative Platforms for mRNA Vaccine Development. Pathogens 2023, 12, 138. [Google Scholar] [CrossRef]
- Li, Y.; Wang, F.; Yang, J.; Han, Z.; Chen, L.; Jiang, W.; Zhou, H.; Li, T.; Tang, Z.; Deng, J.; et al. Deep Generative Optimization of mRNA Codon Sequences for Enhanced Protein Production and Therapeutic Efficacy. bioRxiv 2024. [Google Scholar] [CrossRef]
- Francis, M.J. Recent Advances in Vaccine Technologies. Vet. Clin. N. Am. Small Anim. Pract. 2018, 48, 231–241. [Google Scholar] [CrossRef]
- Andrade, V.M.; Christensen-Quick, A.; Agnes, J.; Tur, J.; Reed, C.; Kalia, R.; Marrero, I.; Elwood, D.; Schultheis, K.; Purwar, M.; et al. INO-4800 DNA vaccine induces neutralizing antibodies and T cell activity against global SARS-CoV-2 variants. NPJ Vaccines 2021, 6, 121. [Google Scholar] [CrossRef] [PubMed]
- Lindblad, E.B.; Duroux, L. Mineral Adjuvants. In Immunopotentiators in Modern Vaccines; Academic Press: New York, NY, USA, 2017; pp. 347–375. [Google Scholar] [CrossRef]
- Hershberg, R.; Petrov, D.A. Selection on codon bias. Annu. Rev. Genet. 2008, 42, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Tews, B.A.; Meyers, G. Self-Replicating RNA. Methods Mol. Biol. 2017, 1499, 15–35. [Google Scholar] [CrossRef] [PubMed]
- Aljabali, A.A.; Obeid, M.A.; Bashatwah, R.M.; Serrano-Aroca, Á.; Mishra, V.; Mishra, Y.; El-Tanani, M.; Hromić-Jahjefendić, A.; Kapoor, D.N.; Goyal, R.; et al. Nanomaterials and Their Impact on the Immune System. Int. J. Mol. Sci. 2023, 24, 2008. [Google Scholar] [CrossRef]
- Farooq, M.A.; Johnston, A.P.; Trevaskis, N.L. Impact of Nanoparticle Properties on Immune Cell Interactions in the Lymph Node. Acta Biomater. 2024, 193, 65–82. [Google Scholar] [CrossRef]
- Perez-Potti, A.; Rodríguez-Pérez, M.; Polo, E.; Pelaz, B.; Del Pino, P. Nanoparticle-based immunotherapeutics: From the properties of nanocores to the differential effects of administration routes. Adv. Drug Deliv. Rev. 2023, 197, 114829. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, Y.; Chen, Y.; Huang, K. Nano-based approaches in the development of antiviral agents and vaccines. Life Sci. 2021, 265, 118761. [Google Scholar] [CrossRef]
- Curley, S.M.; Putnam, D. Biological Nanoparticles in Vaccine Development. Front. Bioeng. Biotechnol. 2022, 10, 867119. [Google Scholar] [CrossRef]
- Sun, C.; Kang, Y.-F.; Fang, X.-Y.; Liu, Y.-N.; Bu, G.-L.; Wang, A.-J.; Li, Y.; Zhu, Q.-Y.; Zhang, H.; Xie, C.; et al. A gB nanoparticle vaccine elicits a protective neutralizing antibody response against EBV. Cell Host Microbe 2023, 31, 1882–1897.E10. [Google Scholar] [CrossRef]
- Lyu, J.H.; Liou, G.G.; Wang, M.; Kan, M.C. The genetic, biophysical and immunological studies of a self-adjuvanted protein nanoparticle. Vaccine 2025, 56, 127087. [Google Scholar] [CrossRef]
- Widge, A.T.; Hofstetter, A.R.; Houser, K.V.; Awan, S.F.; Chen, G.L.; Burgos Florez, M.C.; Berkowitz, N.M.; Mendoza, F.; Hendel, C.S.; Holman, L.A.; et al. An influenza hemagglutinin stem nanoparticle vaccine induces cross-group 1 neutralizing antibodies in healthy adults. Sci. Transl. Med. 2023, 15, eade4790. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, A.; Felgner, J.; Jain, A.; Jan, S.; Albin, T.J.; Badten, A.J.; Gregory, A.E.; Nakajima, R.; Jasinskas, A.; Felgner, P.L.; et al. Engineering Protein Nanoparticles Functionalized with an Immunodominant Coxiella burnetii Antigen to Generate a Q Fever Vaccine. Bioconjug. Chem. 2023, 34, 1653–1666. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Shen, C.; Li, M.; Ma, S.; Liu, C.; Huang, J.; Ren, Z.; Yang, Y.; Zhao, M.; Xie, Q. Programmable Macrophage Vesicle Based Bionic Self-Adjuvanting Vaccine for Immunization against Monkeypox Virus. Adv. Sci. 2025, 12, 2408608. [Google Scholar] [CrossRef]
- Martínez-Pérez, A.; Igea, A.; Estévez, O.; Ferreira, C.M.; Torrado, E.; Castro, A.G.; Fernández, C.; Spetz, A.L.; Adam, L.; López González, M.; et al. Changes in the Immune Phenotype and Gene Expression Profile Driven by a Novel Tuberculosis Nanovaccine: Short and Long-Term Post-immunization. Front. Immunol. 2020, 11, 589863. [Google Scholar] [CrossRef] [PubMed]
- Essink, B.J.; Shapiro, C.; Isidro, M.G.D.; Bradley, P.; Pragalos, A.; Bloch, M.; Santiaguel, J.; Frias, M.V.; Miyakis, S.; Alves de Mesquita, M.; et al. Safety and immunogenicity of a modified mRNA-lipid nanoparticle vaccine candidate against COVID-19: Results from a phase 1, dose-escalation study. Hum. Vaccin. Immunother. 2024, 20, 2408863. [Google Scholar] [CrossRef]
- Mayer, R.L.; Verbeke, R.; Asselman, C.; Aernout, I.; Gul, A.; Eggermont, D.; Boucher, K.; Thery, F.; Maia, T.M.; Demol, H.; et al. Immunopeptidomics-based design of mRNA vaccine formulations against Listeria monocytogenes. Nat. Commun. 2022, 13, 6075. [Google Scholar] [CrossRef]
- Wan, J.; Wang, C.; Wang, Z.; Wang, L.; Wang, H.; Zhou, M.; Fu, Z.F.; Zhao, L. CXCL13 promotes broad immune responses induced by circular RNA vaccines. Proc. Natl. Acad. Sci. USA 2024, 121, e2406434121. [Google Scholar] [CrossRef]
- Marques Neto, L.M.; Kipnis, A.; Junqueira-Kipnis, A.P. Role of Metallic Nanoparticles in Vaccinology: Implications for Infectious Disease Vaccine Development. Front. Immunol. 2017, 8, 239. [Google Scholar] [CrossRef]
- Hoseini, Z.S.; Hajizade, A.; Easton, A.J.; Ahmadian, G.; Ramezani, F. A meta-analysis of the efficiency of metal nanoparticles in vaccine delivery against infectious disease. Nanomedicine 2021, 16, 481–495. [Google Scholar] [CrossRef]
- Friedman-Klabanoff, D.J.; Berry, A.A.; Travassos, M.A.; Shriver, M.; Cox, C.; Butts, J.; Lundeen, J.S.; Strauss, K.A.; Joshi, S.; Shrestha, B.; et al. Recombinant Full-length Plasmodium falciparum Circumsporozoite Protein-Based Vaccine Adjuvanted With Glucopyranosyl Lipid A-Liposome Quillaja saponaria 21: Results of Phase 1 Testing with Malaria Challenge. J. Infect. Dis. 2024, 229, 1883–1893. [Google Scholar] [CrossRef]
- Pollock, K.M.; Borges, Á.H.; Cheeseman, H.M.; Rosenkrands, I.; Schmidt, K.L.; Søndergaard, R.E.; Day, S.; Evans, A.; McFarlane, L.R.; Joypooranachandran, J.; et al. An investigation of trachoma vaccine regimens by the chlamydia vaccine CTH522 administered with cationic liposomes in healthy adults (CHLM-02): A phase 1, double-blind trial. Lancet Infect. Dis. 2024, 24, 829–844. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, A.; Rastogi, A.; Ayyavoo, V.; Srivastava, S. Nanotechnology-Based Approaches for the Development of Diagnostics, Therapeutics, and Vaccines. Monoclon. Antibodies Immunodiagn. Immunother. 2014, 33, 186–191. [Google Scholar] [CrossRef]
- Ward, B.J.; Gobeil, P.; Séguin, A.; Atkins, J.; Boulay, I.; Charbonneau, P.Y.; Couture, M.; D’Aoust, M.A.; Dhaliwall, J.; Finkle, C.; et al. Phase 1 randomized trial of a plant-derived virus-like particle vaccine for COVID-19. Nat. Med. 2021, 27, 1071–1078. [Google Scholar] [CrossRef]
- Gobeil, P.; Pillet, S.; Boulay, I.; Charland, N.; Lorin, A.; Cheng, M.P.; Vinh, D.C.; Boutet, P.; Van Der Most, R.; Roman, F.; et al. Durability and cross-reactivity of immune responses induced by a plant-based virus-like particle vaccine for COVID-19. Nat. Commun. 2022, 13, 6905. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Ding, X.; Zhao, J.; Zeng, J.; Zhou, Y.; Xiao, W.; Hua, D.; Liu, M.; Guo, H.; Zhang, Y.; et al. A novel pan-epitope based nanovaccine self-assembled with CpG enhances immune responses against flavivirus. J. Nanobiotechnol. 2024, 22, 738. [Google Scholar] [CrossRef]
- Langley, J.M.; Gantt, S.; Halperin, S.A.; Ward, B.; McNeil, S.; Ye, L.; Cai, Y.; Smith, B.; Anderson, D.E.; Mitoma, F.D. An enveloped virus-like particle alum-adjuvanted cytomegalovirus vaccine is safe and immunogenic: A first-in-humans Canadian Immunization Research Network (CIRN) study. Vaccine 2024, 42, 713–722. [Google Scholar] [CrossRef]
- Sokouti, B.; Amjad, E. Chapter 16—Systems immunology. In Systems Biology and In-Depth Applications for Unlocking Diseases; Sokouti, B., Ed.; Academic Press: New York, NY, USA, 2025; pp. 207–217. [Google Scholar] [CrossRef]
- Skerrett-Byrne Anthony, D.; Jiang Chen, C.; Nixon, B.; Hondermarck, H. Transcriptomics. In Encyclopedia of Cell Biology, 2nd ed.; Bradshaw, R.A., Hart, G.W., Stahl, P.D., Eds.; Academic Press: Oxford, UK, 2023; pp. 363–371. [Google Scholar] [CrossRef]
- Al-Amrani, S.; Al-Jabri, Z.; Al-Zaabi, A.; Alshekaili, J.; Al-Khabori, M. Proteomics: Concepts and applications in human medicine. World J. Biol. Chem. 2021, 12, 57–69. [Google Scholar] [CrossRef]
- Yue, R.; Dutta, A. Computational systems biology in disease modeling and control, review and perspectives. Npj Syst. Biol. Appl. 2022, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef]
- Rao, A.; Strauss, O.; Kokkinou, E.; Bruchard, M.; Tripathi, K.P.; Schlums, H.; Carrasco, A.; Mazzurana, L.; Konya, V.; Villablanca, E.J.; et al. Cytokines regulate the antigen-presenting characteristics of human circulating and tissue-resident intestinal ILCs. Nat. Commun. 2020, 11, 2049. [Google Scholar] [CrossRef]
- Kambayashi, T.; Laufer, T.M. Atypical MHC class II-expressing antigen-presenting cells: Can anything replace a dendritic cell? Nat. Rev. Immunol. 2014, 14, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Kealy, L.; Good-Jacobson, K.L. Advances in understanding the formation and fate of B-cell memory in response to immunization or infection. Oxf. Open Immunol. 2021, 2, iqab018. [Google Scholar] [CrossRef]
- Muñoz-Wolf, N.; Lavelle, E.C. A Guide to IL-1 family cytokines in adjuvanticity. FEBS J. 2018, 285, 2377–2401. [Google Scholar] [CrossRef]
- Netea, M.G.; van de Veerdonk, F.L.; van der Meer, J.W.; Dinarello, C.A.; Joosten, L.A. Inflammasome-independent regulation of IL-1-family cytokines. Annu. Rev. Immunol. 2015, 33, 49–77. [Google Scholar] [CrossRef]
- Xu, Y.D.; Cheng, M.; Shang, P.P.; Yang, Y.Q. Role of IL-6 in dendritic cell functions. J. Leukoc. Biol. 2022, 111, 695–709. [Google Scholar] [CrossRef]
- Raeber, M.E.; Zurbuchen, Y.; Impellizzieri, D.; Boyman, O. The role of cytokines in T-cell memory in health and disease. Immunol. Rev. 2018, 283, 176–193. [Google Scholar] [CrossRef] [PubMed]
- Shuaib, M.; Adroub, S.; Mourier, T.; Mfarrej, S.; Zhang, H.; Esau, L.; Alsomali, A.; Alofi, F.S.; Ahmad, A.N.; Shamsan, A.; et al. Impact of the SARS-CoV-2 nucleocapsid 203K/204R mutations on the inflammatory immune response in COVID-19 severity. Genome Med. 2023, 15, 54. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, B.; Valizadeh, S.; Ghaffari, H.; Vahedi, A.; Karbalaei, M.; Eslami, M. A global treatments for coronaviruses including COVID-19. J. Cell. Physiol. 2020, 235, 9133–9142. [Google Scholar] [CrossRef]
- Wang, W.; Feng, S.; Ye, Z.; Gao, H.; Lin, J.; Ouyang, D. Prediction of lipid nanoparticles for mRNA vaccines by the machine learning algorithm. Acta Pharm. Sin. B 2022, 12, 2950–2962. [Google Scholar] [CrossRef]
- Haghmorad, D.; Eslami, M.; Orooji, N.; Halabitska, I.; Kamyshna, I.; Kamyshnyi, O.; Oksenych, V. mRNA vaccine platforms: Linking infectious disease prevention and cancer immunotherapy. Front. Bioeng. Biotechnol. 2025, 13, 1547025. [Google Scholar] [CrossRef]
- Yousefi, B.; Eslami, M. Genetic and structure of novel coronavirus COVID-19 and molecular mechanisms in the pathogenicity of coronaviruses. Rev. Res. Med. Microbiol. 2022, 33, e180–e188. [Google Scholar] [CrossRef]
- Maharjan, R.; Kim, K.H.; Lee, K.; Han, H.-K.; Jeong, S.H. Machine learning-driven optimization of mRNA-lipid nanoparticle vaccine quality with XGBoost/Bayesian method and ensemble model approaches. J. Pharm. Anal. 2024, 14, 100996. [Google Scholar] [CrossRef]
- Suyash, S.; Khan, W.H.; Maitra, P.; Jangid, V.; Punia, P.; Mishra, A. Machine Learning Optimization Approach to Design Multi-Epitope Marburg Vaccine Construct. Biotech. Res. Asia 2024, 21, 1463–1484. [Google Scholar] [CrossRef]
- Prosper, P.; Rodríguez Puertas, R.; Guérin, D.M.A.; Branda, M.M. Computational method for designing vaccines applied to virus-like particles (VLPs) as epitope carriers. Vaccine 2024, 42, 3916–3929. [Google Scholar] [CrossRef]
- Garmeh Motlagh, F.; Azimzadeh Irani, M.; Masoomi Nomandan, S.Z.; Assadizadeh, M. Computational design and investigation of the monomeric spike SARS-CoV-2-ferritin nanocage vaccine stability and interactions. Front. Mol. Biosci. 2024, 11, 1403635. [Google Scholar] [CrossRef]
- Torres-Sangiao, E.; Holban, A.M.; Gestal, M.C. Advanced Nanobiomaterials: Vaccines, Diagnosis and Treatment of Infectious Diseases. Molecules 2016, 21, 867. [Google Scholar] [CrossRef]
- Blank, F.; Stumbles, P.A.; Seydoux, E.; Holt, P.G.; Fink, A.; Rothen-Rutishauser, B.; Strickland, D.H.; von Garnier, C. Size-dependent uptake of particles by pulmonary antigen-presenting cell populations and trafficking to regional lymph nodes. Am. J. Respir. Cell Mol. Biol. 2013, 49, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, A.S.; Alonso, M.J.; de la Fuente, M. Nanoengineering of vaccines using natural polysaccharides. Biotechnol. Adv. 2015, 33, 1279–1293. [Google Scholar] [CrossRef]
- Ni, Q.; Zhang, F.; Liu, Y.; Wang, Z.; Yu, G.; Liang, B.; Niu, G.; Su, T.; Zhu, G.; Lu, G.; et al. A bi-adjuvant nanovaccine that potentiates immunogenicity of neoantigen for combination immunotherapy of colorectal cancer. Sci. Adv. 2020, 6, eaaw6071. [Google Scholar] [CrossRef]
- Lai, C.-J.; Kim, D.; Kang, S.; Li, K.; Cha, I.; Sasaki, A.; Porras, J.; Xia, T.; Jung, J.U. Viral codon optimization on SARS-CoV-2 Spike boosts immunity in the development of COVID-19 mRNA vaccines. J. Med. Virol. 2023, 95, e29183. [Google Scholar] [CrossRef]
- Surnar, B.; Kamran, M.Z.; Shah, A.S.; Basu, U.; Kolishetti, N.; Deo, S.; Jayaweera, D.T.; Daunert, S.; Dhar, S. Orally Administrable Therapeutic Synthetic Nanoparticle for Zika Virus. ACS Nano 2019, 13, 11034–11048. [Google Scholar] [CrossRef] [PubMed]
- Kremsner, P.G.; Mann, P.; Kroidl, A.; Leroux-Roels, I.; Schindler, C.; Gabor, J.J.; Schunk, M.; Leroux-Roels, G.; Bosch, J.J.; Fendel, R.; et al. Safety and immunogenicity of an mRNA-lipid nanoparticle vaccine candidate against SARS-CoV-2: A phase 1 randomized clinical trial. Wien. Klin. Wochenschr. 2021, 133, 931–941. [Google Scholar] [CrossRef]
- Cohen, C.J.; Gartner, J.J.; Horovitz-Fried, M.; Shamalov, K.; Trebska-McGowan, K.; Bliskovsky, V.V.; Parkhurst, M.R.; Ankri, C.; Prickett, T.D.; Crystal, J.S.; et al. Isolation of neoantigen-specific T cells from tumor and peripheral lymphocytes. J. Clin. Invest. 2015, 125, 3981–3991. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.J.R.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Børresen-Dale, A.-L.; et al. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef]
- Li, L.; Goedegebuure, P.; Mardis, E.R.; Ellis, M.J.; Zhang, X.; Herndon, J.M.; Fleming, T.P.; Carreno, B.M.; Hansen, T.H.; Gillanders, W.E. Cancer genome sequencing and its implications for personalized cancer vaccines. Cancers 2011, 3, 4191–4211. [Google Scholar] [CrossRef] [PubMed]
- Wolter, N.; Jassat, W.; Walaza, S.; Welch, R.; Moultrie, H.; Groome, M.; Amoako, D.G.; Everatt, J.; Bhiman, J.N.; Scheepers, C.; et al. Early assessment of the clinical severity of the SARS-CoV-2 omicron variant in South Africa: A data linkage study. Lancet 2022, 399, 437–446. [Google Scholar] [CrossRef]
- Nyberg, T.; Ferguson, N.M.; Nash, S.G.; Webster, H.H.; Flaxman, S.; Andrews, N.; Hinsley, W.; Bernal, J.L.; Kall, M.; Bhatt, S.; et al. Comparative analysis of the risks of hospitalisation and death associated with SARS-CoV-2 omicron (B.1.1.529) and delta (B.1.617.2) variants in England: A cohort study. Lancet 2022, 399, 1303–1312. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Schumacher, T.N.; Schreiber, R.D. Neoantigens in cancer immunotherapy. Science 2015, 348, 69–74. [Google Scholar] [CrossRef]
- Hacohen, N.; Fritsch, E.F.; Carter, T.A.; Lander, E.S.; Wu, C.J. Getting personal with neoantigen-based therapeutic cancer vaccines. Cancer Immunol. Res. 2013, 1, 11–15. [Google Scholar] [CrossRef]
- Ott, P.A.; Hu, Z.; Keskin, D.B.; Shukla, S.A.; Sun, J.; Bozym, D.J.; Zhang, W.; Luoma, A.; Giobbie-Hurder, A.; Peter, L.; et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature 2017, 547, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Nasrullah, A.; Nizamullah, F.N.U.; Shah, Z. Ai in healthcare: Using cutting-edge technologies to revolutionize vaccine development and distribution. JURIHUM J. Inov. Dan Hum. 2023, 1, 17–29. [Google Scholar]
- Zhang, L.; Huang, J.; Chen, X.; Pan, C.; He, Y.; Su, R.; Guo, D.; Yin, S.; Wang, S.; Zhou, L.; et al. Self-assembly nanovaccine containing TLR7/8 agonist and STAT3 inhibitor enhances tumor immunotherapy by augmenting tumor-specific immune response. J. Immunother. Cancer 2021, 9, e003132. [Google Scholar] [CrossRef]
- Guimaraes, L.C.; Costa, P.A.C.; Scalzo Júnior, S.R.A.; Ferreira, H.A.S.; Braga, A.C.S.; de Oliveira, L.C.; Figueiredo, M.M.; Shepherd, S.; Hamilton, A.; Queiroz-Junior, C.M.; et al. Nanoparticle-based DNA vaccine protects against SARS-CoV-2 variants in female preclinical models. Nat. Commun. 2024, 15, 590. [Google Scholar] [CrossRef] [PubMed]
- Hoces, D.; Miguens Blanco, J.; Hernández-López, R.A. A synthetic biology approach to engineering circuits in immune cells. Immunol. Rev. 2023, 320, 120–137. [Google Scholar] [CrossRef]
- Zhu, B.; Yin, H.; Zhang, D.; Zhang, M.; Chao, X.; Scimeca, L.; Wu, M.-R. Synthetic biology approaches for improving the specificity and efficacy of cancer immunotherapy. Cell. Mol. Immunol. 2024, 21, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Irvine, D.J.; Hanson, M.C.; Rakhra, K.; Tokatlian, T. Synthetic nanoparticles for vaccines and immunotherapy. Chem. Rev. 2015, 115, 11109–11146. [Google Scholar] [CrossRef]
- Haleem, A.; Javaid, M.; Singh, R.P.; Rab, S.; Suman, R. Applications of nanotechnology in medical field: A brief review. Glob. Health J. 2023, 7, 70–77. [Google Scholar] [CrossRef]
- Halamoda-Kenzaoui, B.; Holzwarth, U.; Roebben, G.; Bogni, A.; Bremer-Hoffmann, S. Mapping of the available standards against the regulatory needs for nanomedicines. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 11, e1531. [Google Scholar] [CrossRef]
- Pautler, M.; Brenner, S. Nanomedicine: Promises and challenges for the future of public health. Int. J. Nanomed. 2010, 5, 803–809. [Google Scholar]
- Berube, D.M. The public acceptance of nanomedicine: A personal perspective. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2009, 1, 2–5. [Google Scholar] [CrossRef]
- Sharifi, S.; Mahmoud, N.N.; Voke, E.; Landry, M.P.; Mahmoudi, M. Importance of Standardizing Analytical Characterization Methodology for Improved Reliability of the Nanomedicine Literature. Nanomicro Lett. 2022, 14, 172. [Google Scholar] [CrossRef]
- De Jong, W.H.; Geertsma, R.E.; Borchard, G. Regulatory safety evaluation of nanomedical products: Key issues to refine. Drug Deliv. Transl. Res. 2022, 12, 2042–2047. [Google Scholar] [CrossRef]
- Gupta, R.; Arora, K.; Roy, S.S.; Joseph, A.; Rastogi, R.; Arora, N.M.; Kundu, P.K. Platforms, advances, and technical challenges in virus-like particles-based vaccines. Front. Immunol. 2023, 14, 1123805. [Google Scholar] [CrossRef] [PubMed]
- Vallet, T.; Vignuzzi, M. Self-Amplifying RNA: Advantages and Challenges of a Versatile Platform for Vaccine Development. Viruses 2025, 17, 566. [Google Scholar] [CrossRef]
- Verbeke, R.; Lentacker, I.; De Smedt, S.C.; Dewitte, H. Three decades of messenger RNA vaccine development. Nano Today 2019, 28, 100766. [Google Scholar] [CrossRef]
- Wouters, O.J.; Shadlen, K.C.; Salcher-Konrad, M.; Pollard, A.J.; Larson, H.J.; Teerawattananon, Y.; Jit, M. Challenges in ensuring global access to COVID-19 vaccines: Production, affordability, allocation, and deployment. Lancet 2021, 397, 1023–1034. [Google Scholar] [CrossRef]
- Dolgin, E. The tangled history of mRNA vaccines. Nature 2021, 597, 318–324. [Google Scholar] [CrossRef]
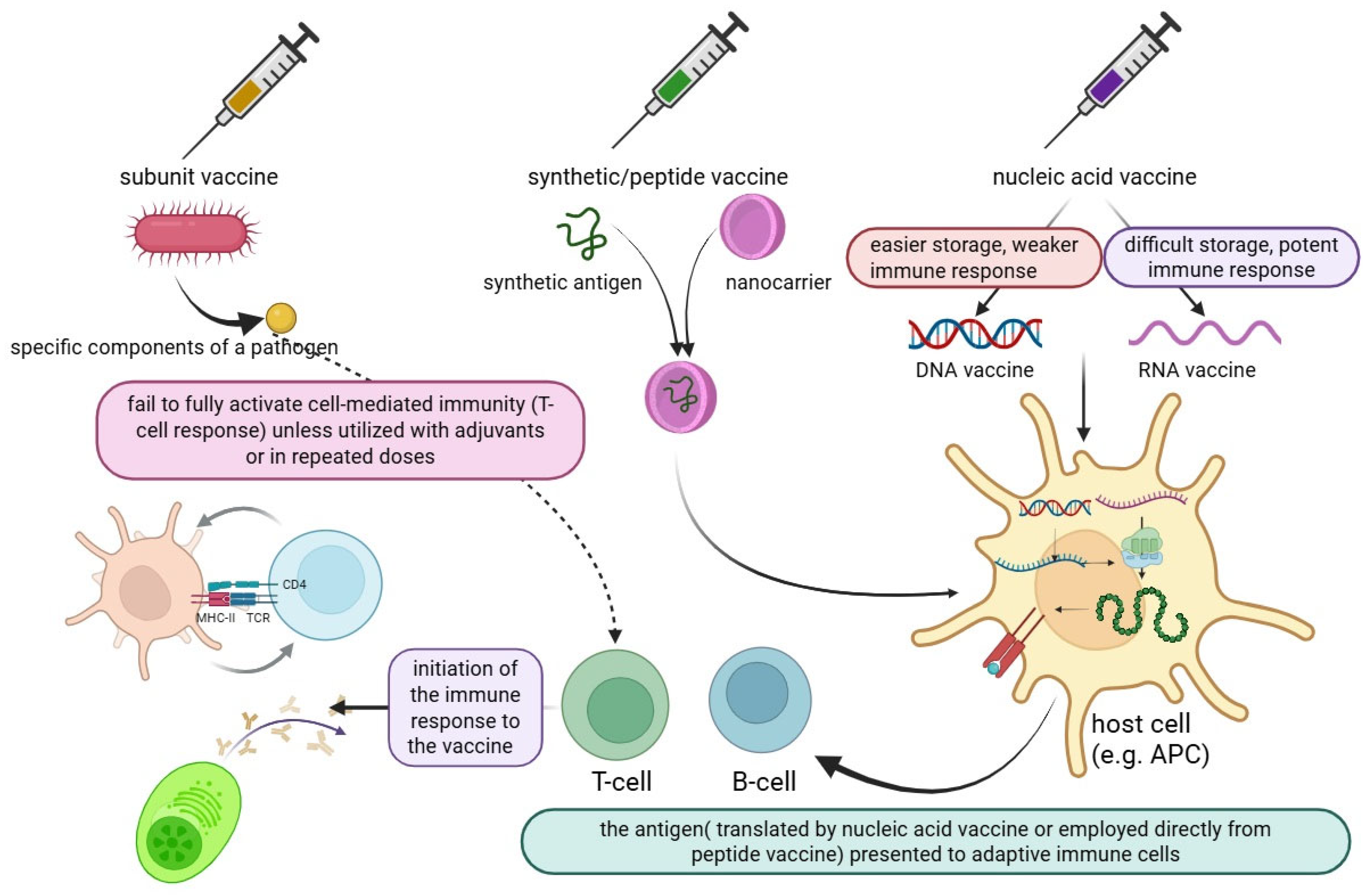
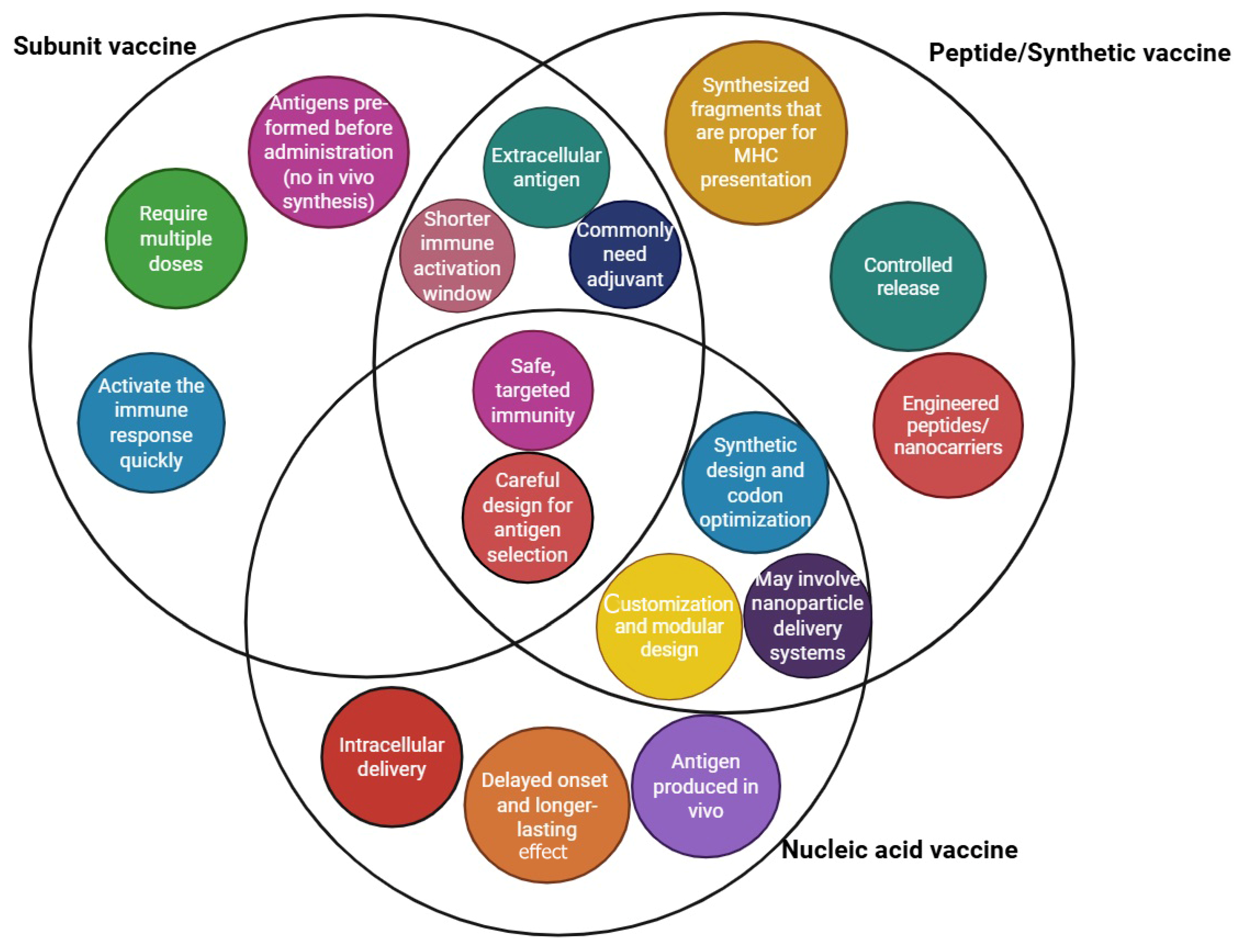
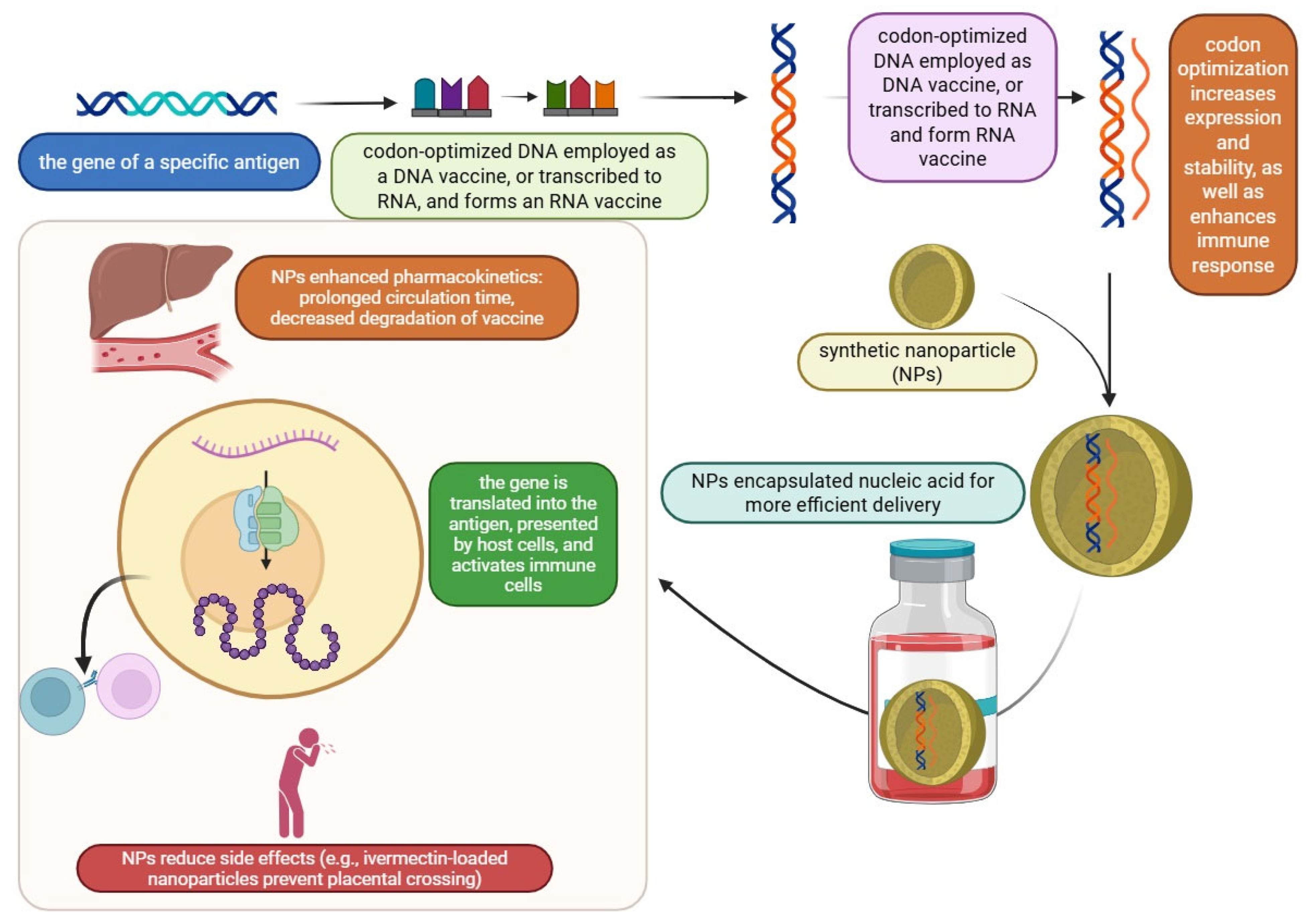
| Adjuvant | Composition/Type | Mechanism of Action | Approved Vaccines |
|---|---|---|---|
| Aluminum salts (Alum) | Mineral salts (e.g., aluminum hydroxide) | Creates a depot effect and activates inflammasomes (e.g., NLRP3), enhancing APC uptake and Th2 responses | Hepatitis A/B, DTaP, HPV |
| MF59 | Squalene-based oil-in-water emulsion | Enhances antigen uptake and recruitment of APCs; promotes cytokine release (IL-6, MCP-1) | Influenza (e.g., Fluad) |
| AS03 | α-tocopherol + squalene emulsion | Stimulates cytokine production (e.g., IL-6) and promotes antigen presentation | H1N1 (e.g., Pandemrix) |
| AS01 | Liposome-based with MPL + QS-21 | Activates TLR4 and promotes Th1-biased immunity through APC maturation | Shingrix (herpes zoster), Malaria (RTS,S) |
| CpG 1018 | Synthetic TLR9 agonist (DNA oligo) | Directly activates plasmacytoid dendritic cells and B cells, promoting Th1 responses | HEPLISAV-B (hepatitis B) |
| Poly I:C | Synthetic dsRNA (TLR3 agonist) | Mimics viral RNA, stimulating IFN production and cytotoxic T cell responses | (Experimental; not yet widely approved) |
| Platform | Mechanism | Examples | Advantages | Limitations | Ref. |
|---|---|---|---|---|---|
| mRNA vaccines | Encodes antigens via synthetic RNA expressed in host cells | Pfizer-BioNTech, Moderna | Rapid production, customizable, strong immune response | Requires cold chain, limited stability | [22,23,24,25,26,43,44] |
| DNA vaccines | Synthetic plasmid DNA encoding antigens delivered into host cells | INO-4800 | High stability, easy storage | Lower efficacy in humans, requires special devices | [26,27,28,42,45] |
| Mineral salts | ↑ the immunogenicity of some vaccines, such as pertussis, diphtheria, poliomyelitis | aluminum adjuvants | ↑ Immune response, improved antigen delivery, and development of new vaccines | Potential side effects, limited human use, and restricted approval | [34,38,46] |
| Emulsions | ↑ the immunogenicity of some vaccines such as influenza. | MF59 AS03 | ↑ Immune response, improved antigen delivery, and the development of new vaccines. | Potential side effects, limited human use, and restricted approval | [34,36,37] |
| Codon optimization | Process of modifying the codons in a gene to enhance protein expression in a specific organism | CodaVax-H1N1 CodaVax-RSV CDX-005 | ↑ Protein production, no in-depth viral function knowledge needed, faster response to a reduced risk of viral reversion | Codon usage bias variability, time-dependent process | [29,30,31,32,33,47] |
| Self-amplifying RNA vaccines | Synthetic RNA replicates inside cells producing more antigens | ARCT-154 | Lower dose needed, and stronger immune response | Novel platform, with safety still under evaluation | [41,42,48] |
| Studies (Year) | Nano Material | Properties | Application | Advantages | Challenges | Ref. |
|---|---|---|---|---|---|---|
| Sun (2023) | Protein-based polymeric nanoparticle | Glycoprotein | EBV | Durable humoral immunity (Nab, IgG, IgA) | No cellular immunity—autoimmunity | [54] |
| Widge (2023) | Protein-based nanoparticle | Hemagglutinin head | Influenza | Durable humoral immunity (Nab, IgG)—cross-reactivity among group 1 influenza | Limited response against group 2 influenza | [56] |
| Ramirez (2023) | Protein-based nanoparticle | Outer membrane protein antigen | Coxiella burnetii | Strong cellular and humoral immune response | Not evaluated in humans—low durability of responses | [57] |
| Martínez-Pérez (2021) | Lipid-protein nanoparticles | Protein antigen | Mycobacterium tuberculosis | Memory T cell induction | Low durability of responses | [59] |
| Essink (2024) | Lipid-based nanoparticle | mRNA | SARS-CoV-2 | Cross-reactivity | Small sample size | [60] |
| Mayer (2022) | Lipid-based nanoparticle | mRNA | Listeria monocytogenes | Strong cellular response | Not evaluated in humans | [61] |
| Wan (2024) | Lipid-based nanoparticle | circRNA | Influenza virus and SARS-CoV-2 | Robust cross reactivity | Not evaluated in humans—allergic-type reaction | [62] |
| Friedman Klabanoff (2024) | Liposomes | full-length recombinant Circumsporo-zoite | Plasmodium falciparum | Favorable safety and tolerability—high avidity of full-length and C-terminal region antibodies | Non-protective immunity | [65] |
| Pollock (2024) | Liposomes | Outer membrane molecule | Chlamydia trachomatis | Humoral (IgG and IgA) and cellular immune response | Small sample size | [66] |
| Gobeil (2021–22) | Virus-like particle | Full-length Spike protein | SARS-CoV-2 | Robust, durable, and cross-reactive humoral immunity—durable Th1 and Th2 response induction | Allergic-type reaction | [68,69] |
| He (2024) | Virus-like particle | Pan-epitope peptide TBT | Flavivirus | High uptake rate | Not evaluated in humans | [70] |
| Langley (2024) | Enveloped virus-like particle | Glycoprotein | CMV | Sustained humoral responses | Low vaccine dose | [71] |
| NAb = neutralizing antibody | ||||||
| Studies (Year) | Tool | Description | Contribution | Challenges | Examples | Ref. |
|---|---|---|---|---|---|---|
| Shuaib (2023) | Transcriptomics and Proteomics in Mutation Analysis | Gene expression profiling and protein-level immune analysis of viral mutations | Reveals how KR mutations in SARS-CoV-2 nucleocapsid drive a stronger inflammatory response | Requires further exploration to link findings to vaccine development | SARS-CoV2 | [84] |
| Wang (2022) | ML-Driven LNP Optimization | Predictive modeling of LNP formulations for mRNA delivery | Identifies optimal LNP structures for enhanced vaccine efficiency | Integration with molecular modeling required further refinement | - | [86] |
| Maharjan (2024) | Microfluidics and Ensemble Models | Fine-tuning microfluidic conditions and lipid mix ratios for mRNA-LNP formulation | Achieves high prediction accuracy in particle properties | Immunogenicity not assessed | - | [89] |
| Suyash (2024) | Computational Epitope Design | Identification of immunogenic proteins for vaccine development | Enables rational design of multi-epitope vaccines | No in vitro/in vivo experimental validation performed | Marburg virus | [90] |
| Prosper (2024) | Molecular Dynamics in VLP Design | Structural analysis of VLP proteins | Assesses the antigenicity and allergenicity | Predictive models require in vitro/in vivo validation | SARS-CoV2 | [91] |
| Garmeh Motlagh (2024) | RBD-Nanoparticle Analysis | Structural and immunogenicity assessment of SARS-CoV-2 RBD-ferritin nanoparticles | Evaluates the stability, flexibility, and immune response | Limited immune response durability; requires further optimization and in vivo validation | SARS-CoV2 | [92] |
| Mayer (2022) | Immunopeptidomics for mRNA Vaccine Targeting Bacteria | Peptide profiling of infected cells to guide vaccine design | Identifies the antigenic targets for intracellular bacterial vaccines | Translating findings to human applications remains a challenge | Listeria monocytogenes | [61] |
| Vaccine Platform | Synthetic Biology Contribution | Nanotechnology Contribution | Systems Immunology Contribution | Example | Outcome | Ref. |
|---|---|---|---|---|---|---|
| mRNA-LNP Vaccines | HSV-1 gB codon_optimized mRNA sequences for SARS-CoV-2 spike protein expression | Encapsulation of optimized mRNA in lipid nanoparticles to protect mRNA, enhance delivery, ensure endosomal escape, and increase intracellular expression of the antigen. | Induction of strong humoral and cellular immune responses, including high titers of neutralizing antibodies and robust T cell activation against SARS-CoV-2 and its variants. | HSVgB codon-optimized Delta Spike mRNA vaccine co-expressed with HSV-1 ICP27. | Spike-specific IgG, neutralizing antibodies were markedly increasing, with greater immunological response at reduced dosages of mRNA. Decreased likelihood of adverse effects. | [97] |
| Self-Assembling Nanovaccines | Combination of TLR7/8 Agonists and STAT3 Inhibitors | Self-assembling vehicle-free multicomponent antitumor nanovaccine (SVMAV) | A multi-layered approach to activate and balance various parts of the immune system to improve the immune response against tumors. | HPV vaccine (Gardasil), experimental influenza vaccines | Enhanced tumor control, a stronger immune response against malignancies, and the possibility of better results from cancer immunotherapy. | [111] |
| DNA-Nanoparticle Vaccines | recombinant HexaPro spike plasmid DNA against SARS-CoV-2 Gamma lineage | Nanoparticles to boost cellular delivery and uptake | Modeling immune response to improve efficiency of dosing and delivery | DNA nanoparticle vaccine against SARS- CoV-2 ZyCoV-D | Ameliorate transfection efficiency and stronger immune activation | [112] |
| Synthetic Immune Circuits | Logic-based receptor design (e.g., SynNotch), express gene control | Nanocarriers for targeted transfer of genetic circuits | Modeling and analysis of immune responses to optimize design | SynNotch T cells activated only in the presence of the tumor antigen | Increased specificity, decreased side effects, and better control of immune functions | [113] |
| Personalized Cancer Vaccines | Neoantigen identification and mRNA synthesis | Lipid nanoparticles for personalized delivery | Patient-specific immune profiling to tailor vaccines | BioNTech mRNA-based personalized cancer vaccines | Individualized immune responses, improved tumor targeting | [109] |
| Platform Type | Immunogenicity (H/C) * | Manufacturing Complexity | Stability | Cost | Notable Advantages | Key Limitations |
|---|---|---|---|---|---|---|
| mRNA (LNP-based) | High/Moderate | Moderate to High | Low (unless lyophilized) | High | Rapid development, strong cellular response | Cold chain needed; limited long-term data |
| DNA vaccine | Moderate/Low | Low | High | Low | Stable, easy to store | Weaker human efficacy; delivery device required |
| Peptide (adjuvanted) | Low/Moderate | Low | High | Low–Moderate | Safe, defined antigens | Needs adjuvant, often weak CD8+ T-cell activation |
| Protein subunit (e.g., recombinant) | Moderate/High | Moderate | Moderate | Moderate | Proven platforms (e.g., HBV, HPV) | Requires adjuvants; slower production |
| VLP (Virus-like particle) | High/High | High | Moderate | High | Multivalent display; strong B and T cell responses | Complex manufacturing; scalability challenges |
| saRNA (Self-amplifying) | High/High | High | Low | Moderate–High | Lower dose needed; enhanced expression | Still under evaluation; safety and scale-up ongoing |
| Nano-based (liposome, polymer) | High/Variable | High | Moderate | Moderate–High | Tunable targeting; co-delivery of antigen + adjuvant | Complex formulation; regulatory challenges |
| Synthetic biology + NP + systems immunology | Very High/High | Very High | Moderate | High | Precision-targeted, personalized, and adaptable platforms | Requires infrastructure and advanced data analytics |
| Pathogen/Disease Type | Target | Vaccine Platform | Key Outcome |
|---|---|---|---|
| Viral | SARS-CoV-2 (Omicron) | mRNA in lipid nanoparticles (CV0501) | Strong neutralizing antibody and Th1-skewed T cell responses |
| Bacterial | Listeria monocytogenes (LMON_0149 antigen) | mRNA in lipid nanoparticles | Robust CD8⁺ T cell activation and antigen-specific IFN-γ production |
| Parasitic | Plasmodium falciparum (Circumsporozoite protein) | Protein subunit + GLA-LSQ nanoliposome | Elevated IgG titers with a favorable safety profile |
| Cancer (Non-Infectious) | Tumor neoantigens | Personalized mRNA vaccine + LNP | Induction of tumor-specific T cell responses and tumor regression |
| Autoimmune Conditions | Experimental autoimmune encephalomyelitis (EAE) | Tolerogenic nanoparticle-based vaccine | Reduction in disease severity via antigen-specific tolerance |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eslami, M.; Fadaee Dowlat, B.; Yaghmayee, S.; Habibian, A.; Keshavarzi, S.; Oksenych, V.; Naderian, R. Next-Generation Vaccine Platforms: Integrating Synthetic Biology, Nanotechnology, and Systems Immunology for Improved Immunogenicity. Vaccines 2025, 13, 588. https://doi.org/10.3390/vaccines13060588
Eslami M, Fadaee Dowlat B, Yaghmayee S, Habibian A, Keshavarzi S, Oksenych V, Naderian R. Next-Generation Vaccine Platforms: Integrating Synthetic Biology, Nanotechnology, and Systems Immunology for Improved Immunogenicity. Vaccines. 2025; 13(6):588. https://doi.org/10.3390/vaccines13060588
Chicago/Turabian StyleEslami, Majid, Bahram Fadaee Dowlat, Shayan Yaghmayee, Anoosha Habibian, Saeedeh Keshavarzi, Valentyn Oksenych, and Ramtin Naderian. 2025. "Next-Generation Vaccine Platforms: Integrating Synthetic Biology, Nanotechnology, and Systems Immunology for Improved Immunogenicity" Vaccines 13, no. 6: 588. https://doi.org/10.3390/vaccines13060588
APA StyleEslami, M., Fadaee Dowlat, B., Yaghmayee, S., Habibian, A., Keshavarzi, S., Oksenych, V., & Naderian, R. (2025). Next-Generation Vaccine Platforms: Integrating Synthetic Biology, Nanotechnology, and Systems Immunology for Improved Immunogenicity. Vaccines, 13(6), 588. https://doi.org/10.3390/vaccines13060588