
Heterologous Prime-Boost Vaccination with Inactivated Influenza Viruses Induces More Effective Cross-Protection than Homologous Repeat Vaccination

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Reagents, and Viruses
2.2. Immunization and Virus Challenge of Mice
2.3. Antibody Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Hemagglutination Inhibition (HAI) Assay
2.5. Lung Viral Titration
2.6. In Vitro IgG Antibody Detection and Cytokine ELISA
2.7. In Vivo Protection Efficacy Test of Immune Sera
2.8. In Vivo Depletion of T Cells
2.9. Intracellular Cytokine Staining and Flow Cytometry Analysis
2.10. Statistical Analyses
3. Results
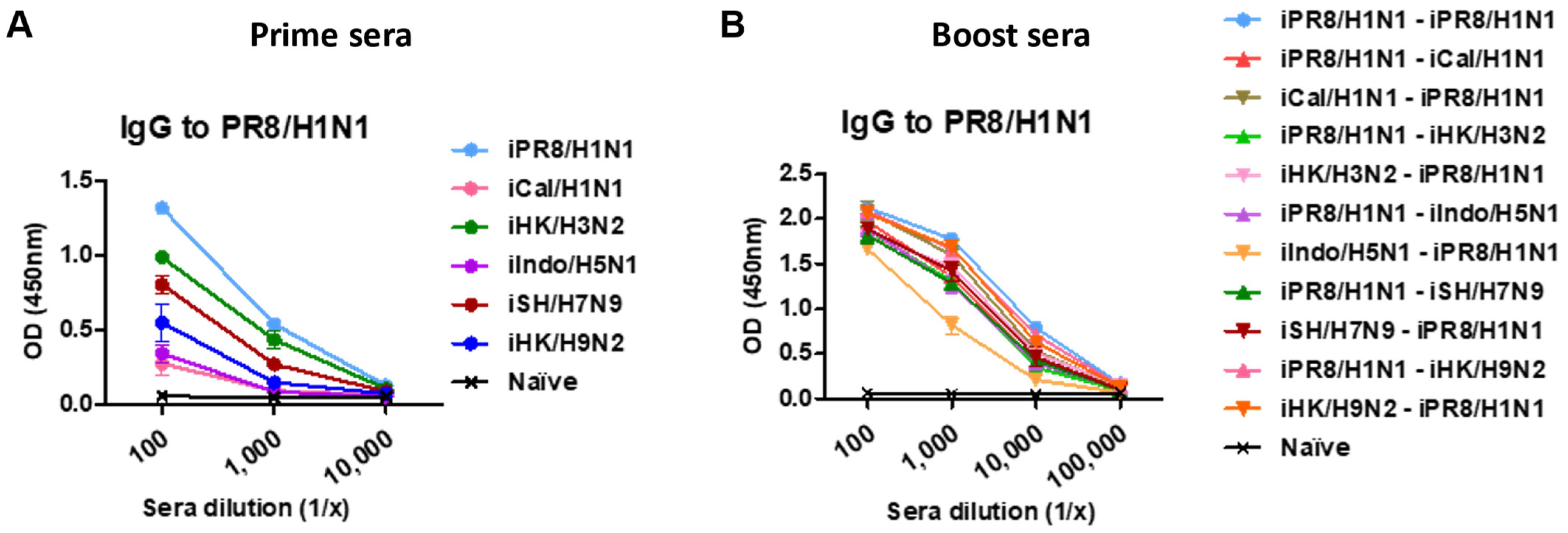
3.1. Heterologous and Heterosubtypic Prime-Boost Vaccinations Induce Cross-Reactive Virus-Specific IgG and Group-Specific HA Stalk Antibodies
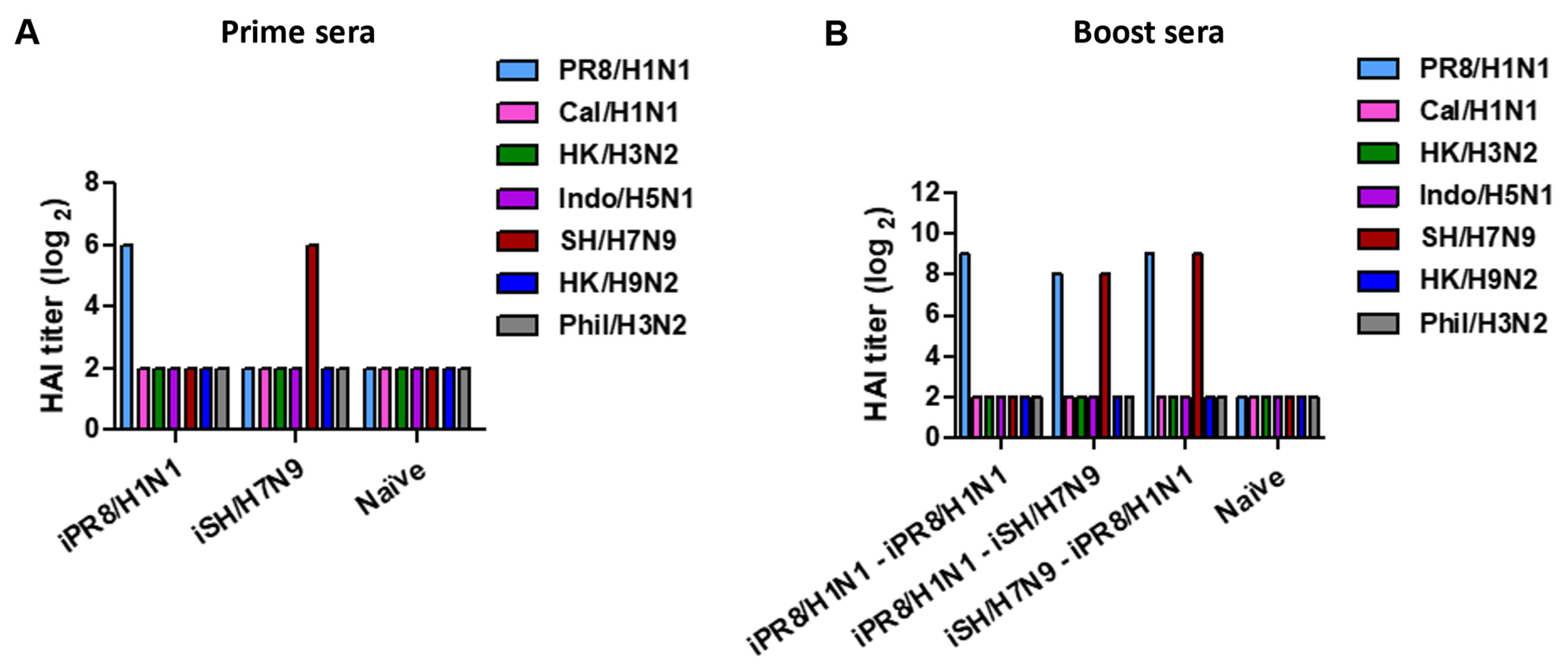
3.2. Heterologous Boost Enhances the Induction of HAI Titers against Prior Prime and Current Boost Virus Strains
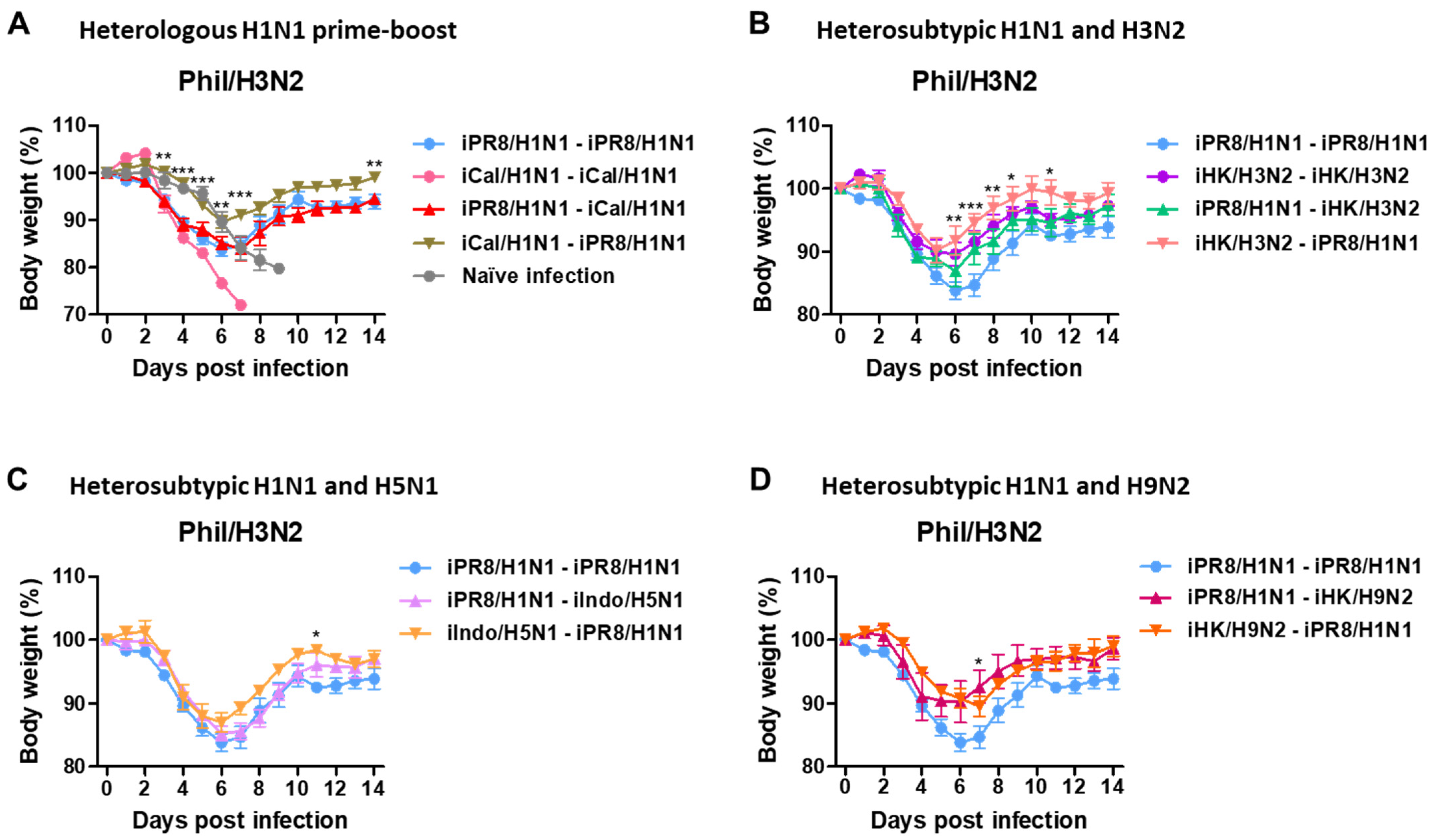
3.3. Heterologous and Heterosubtypic Prime-Boost Vaccinations Induce Cross-Protection
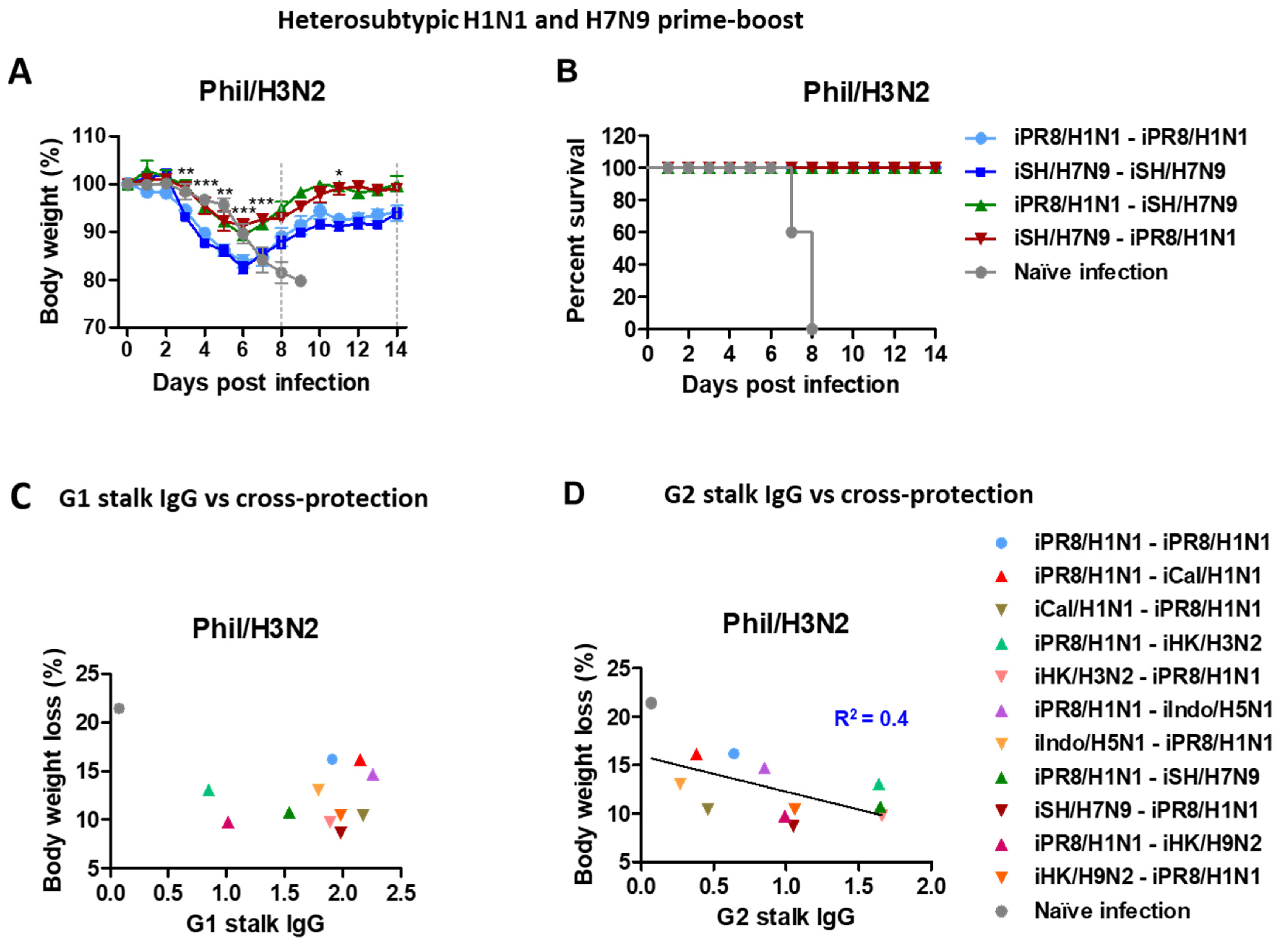
3.4. H1N1 and H7N9 Prime-Boost Vaccination Provides Enhanced Heterosubtypic Protection against H3N2 Virus, Supporting a Correlation with Stalk Antibodies
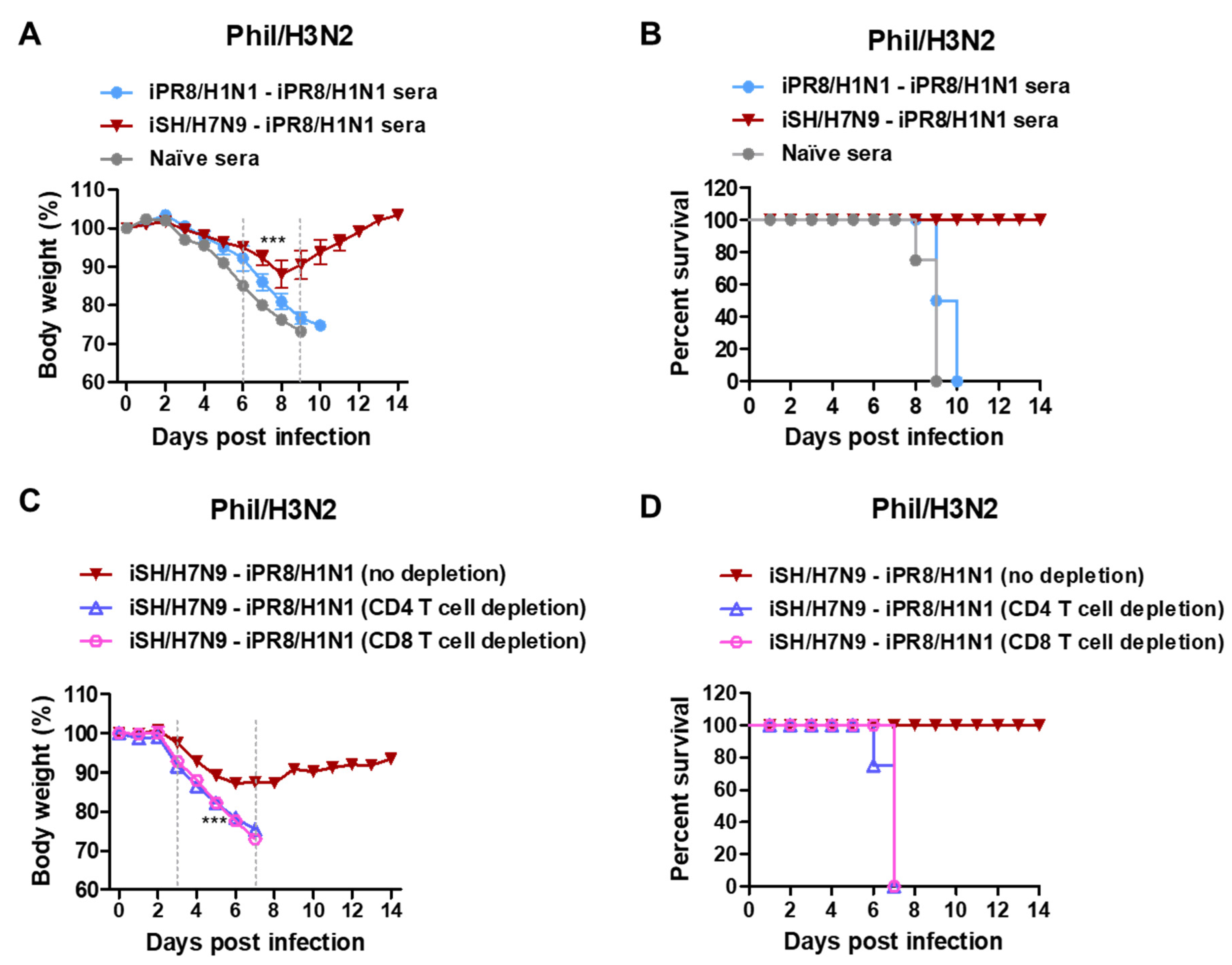
3.5. Immune Sera and T Cells Contribute to Protection against Phil/H3N2 Virus after H7N9-H1N1 Prime-Boost Vaccination
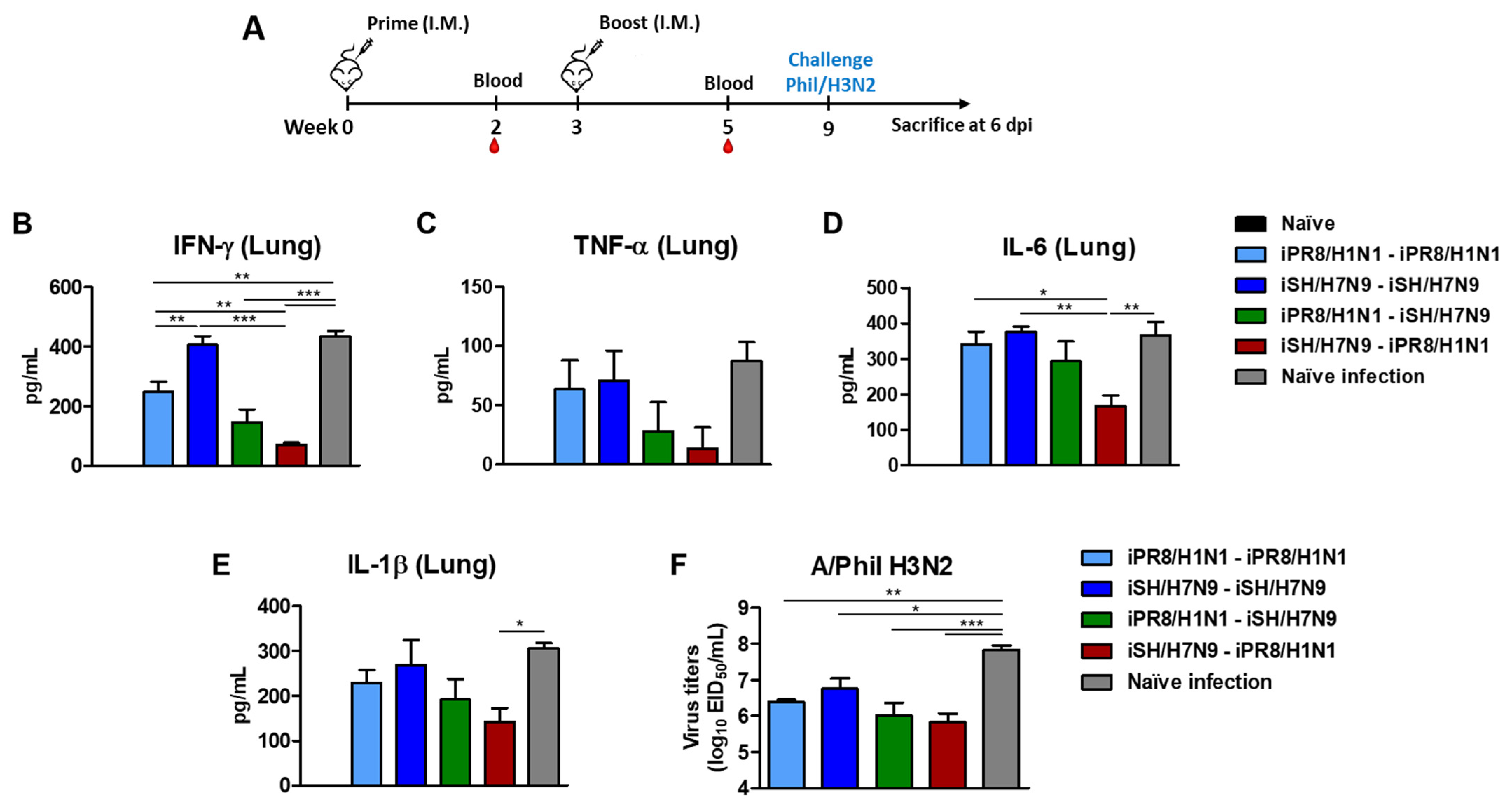
3.6. Heterosubtypic Prime-Boost Vaccination Controls Inflammatory Cytokines and Viral Replication after H3N2 Virus Challenge
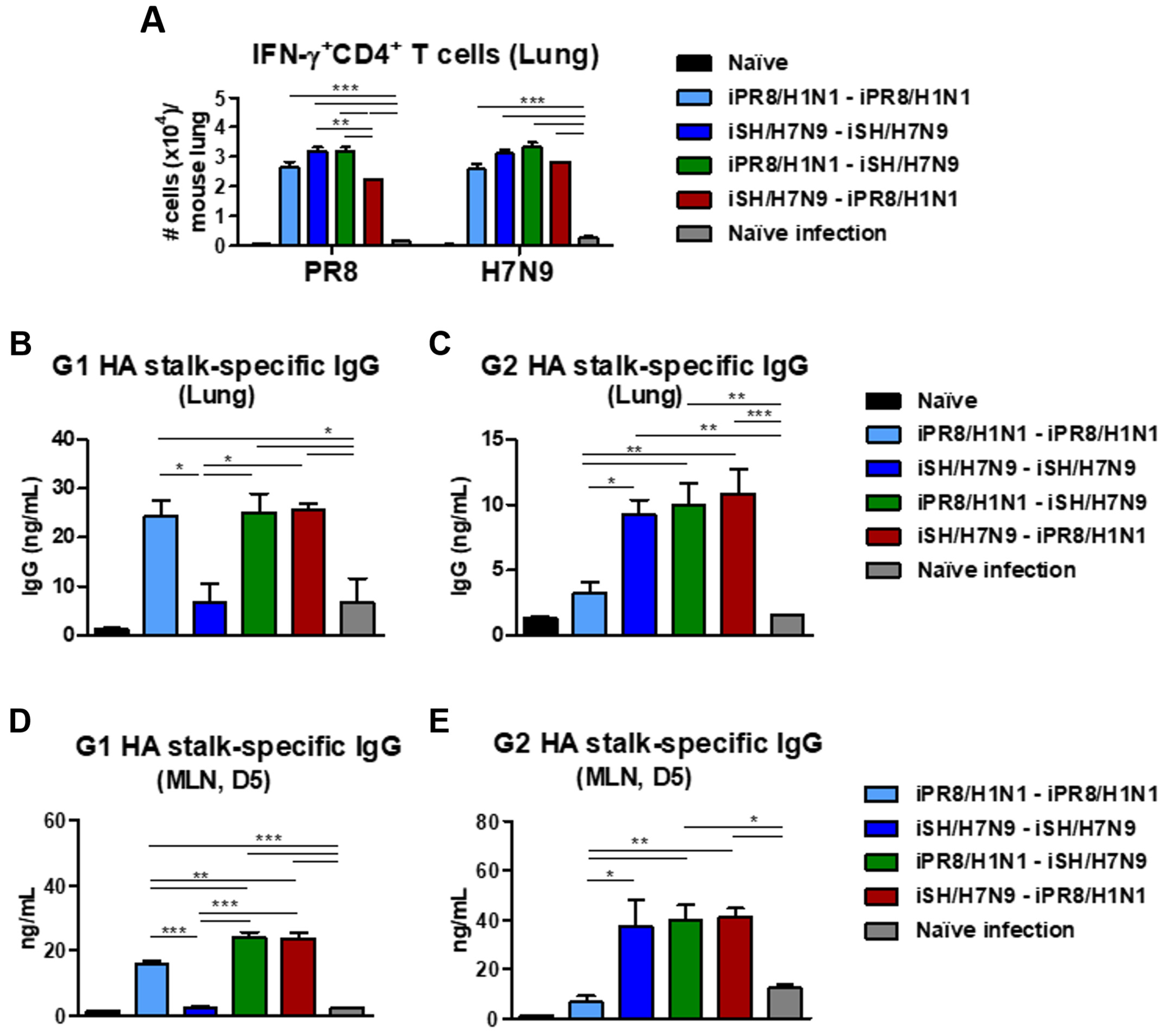
3.7. Heterosubtypic Prime-Boost Vaccination Enhances IFN-γ-Producing Cells and HA Stalk-Specific Antibody-Secreting Cellular Responses after Phil/H3N2 Virus Challenge
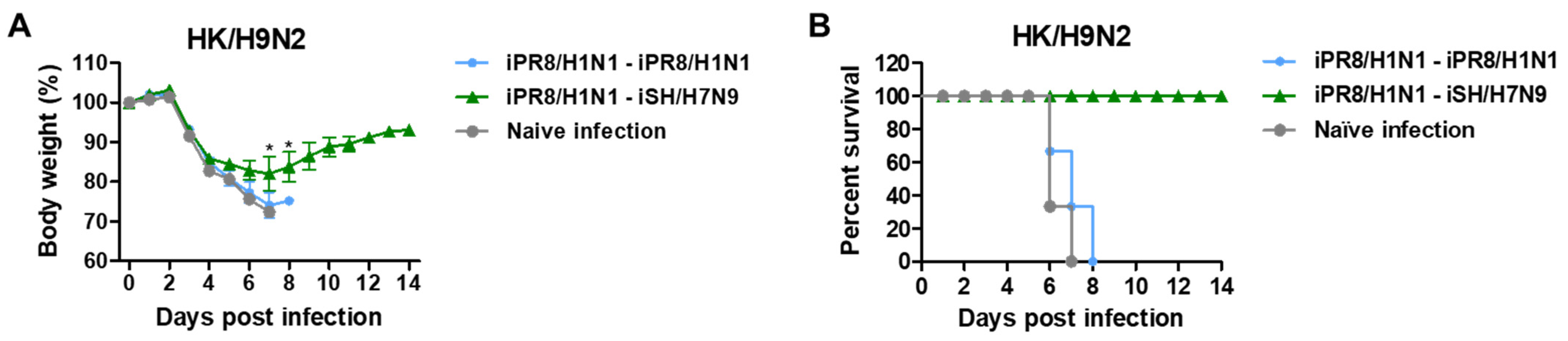
3.8. H1N1-H7N9 Heterosubtypic Prime-Boost Vaccination Confers Survival Protection against H5N1 and H9N2 Viruses
3.9. Hetero-G1 H1N1 Prime-Boost Serum Is Effective in Providing Protection against G1 H5N1 Virus Compared to H7N9-H1N1 G2-G1 HA Virus Prime-Boost
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef]
- Thompson, W.W.; Shay, D.K.; Weintraub, E.; Brammer, L.; Cox, N.; Anderson, L.J.; Fukuda, K. Mortality associated with influenza and respiratory syncytial virus in the United States. JAMA 2003, 289, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Pica, N.; Palese, P. Toward a universal influenza virus vaccine: Prospects and challenges. Annu. Rev. Med. 2013, 64, 189–202. [Google Scholar] [CrossRef]
- Sasaki, S.; Sullivan, M.; Narvaez, C.F.; Holmes, T.H.; Furman, D.; Zheng, N.Y.; Nishtala, M.; Wrammert, J.; Smith, K.; James, J.A.; et al. Limited efficacy of inactivated influenza vaccine in elderly individuals is associated with decreased production of vaccine-specific antibodies. J. Clin. Investig. 2011, 121, 3109–3119. [Google Scholar] [CrossRef]
- Petrie, J.G.; Monto, A.S. Untangling the Effects of Prior Vaccination on Subsequent Influenza Vaccine Effectiveness. J. Infect. Dis. 2017, 215, 841–843. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.J.; Forrest, S.; Ackley, D.H.; Perelson, A.S. Variable efficacy of repeated annual influenza vaccination. Proc. Natl. Acad. Sci. USA 1999, 96, 14001–14006. [Google Scholar] [CrossRef] [PubMed]
- De Bruijn, I.A.; Remarque, E.J.; Jol-van der Zijde, C.M.; van Tol, M.J.; Westendorp, R.G.; Knook, D.L. Quality and quantity of the humoral immune response in healthy elderly and young subjects after annually repeated influenza vaccination. J. Infect. Dis. 1999, 179, 31–36. [Google Scholar] [CrossRef]
- CDC. Seasonal Influenza Vaccine Effectiveness, 2004–2019. Available online: https://www.cdc.gov/flu/vaccines-work/effectiveness-studies.htm#figure (accessed on 1 January 2020).
- Keitel, W.A.; Cate, T.R.; Couch, R.B.; Huggins, L.L.; Hess, K.R. Efficacy of repeated annual immunization with inactivated influenza virus vaccines over a five year period. Vaccine 1997, 15, 1114–1122. [Google Scholar] [CrossRef]
- Zimmerman, R.K.; Nowalk, M.P.; Chung, J.; Jackson, M.L.; Jackson, L.A.; Petrie, J.G.; Monto, A.S.; McLean, H.Q.; Belongia, E.A.; Gaglani, M.; et al. 2014–2015 Influenza Vaccine Effectiveness in the United States by Vaccine Type. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2016, 63, 1564–1573. [Google Scholar] [CrossRef]
- McLean, H.Q.; Thompson, M.G.; Sundaram, M.E.; Meece, J.K.; McClure, D.L.; Friedrich, T.C.; Belongia, E.A. Impact of repeated vaccination on vaccine effectiveness against influenza A(H3N2) and B during 8 seasons. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2014, 59, 1375–1385. [Google Scholar] [CrossRef]
- Leung, V.K.Y.; Carolan, L.A.; Worth, L.J.; Harper, S.A.; Peck, H.; Tilmanis, D.; Laurie, K.L.; Slavin, M.A.; Sullivan, S.G. Influenza vaccination responses: Evaluating impact of repeat vaccination among health care workers. Vaccine 2017, 35, 2558–2568. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, N.; Takeishi, K. Change in the efficacy of influenza vaccination after repeated inoculation under antigenic mismatch: A systematic review and meta-analysis. Vaccine 2018, 36, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Hahn, M.; Coyle, E.M.; King, L.R.; Lin, T.L.; Treanor, J.; Sant, A.; Golding, H. Repeat vaccination reduces antibody affinity maturation across different influenza vaccine platforms in humans. Nat. Commun. 2019, 10, 3338. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, M.; Holmes, T.H.; Maecker, H.T.; Albrecht, R.A.; Dekker, C.L.; He, X.S.; Greenberg, H.B. Diminished B-Cell Response After Repeat Influenza Vaccination. J. Infect. Dis. 2019, 219, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Cobey, S.; Hensley, S.E. Immune history and influenza virus susceptibility. Curr. Opin. Virol. 2017, 22, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Guthmiller, J.J.; Wilson, P.C. Harnessing immune history to combat influenza viruses. Curr. Opin. Immunol. 2018, 53, 187–195. [Google Scholar] [CrossRef]
- Bhatnagar, N.; Kim, K.H.; Subbiah, J.; Park, B.R.; Ko, E.J.; Seong, B.L.; Kang, S.M. Comparison of the effects of different potent adjuvants on enhancing the immunogenicity and cross-protection by influenza virus vaccination in young and aged mice. Antivir. Res. 2022, 197, 105229. [Google Scholar] [CrossRef]
- Park, B.R.; Kim, K.H.; Kotomina, T.; Kim, M.C.; Kwon, Y.M.; Jeeva, S.; Jung, Y.J.; Bhatnagar, N.; Isakova-Sivak, I.; Mezhenskaya, D.; et al. Broad cross protection by recombinant live attenuated influenza H3N2 seasonal virus expressing conserved M2 extracellular domain in a chimeric hemagglutinin. Sci. Rep. 2021, 11, 4151. [Google Scholar] [CrossRef]
- Deng, L.; Mohan, T.; Chang, T.Z.; Gonzalez, G.X.; Wang, Y.; Kwon, Y.M.; Kang, S.M.; Compans, R.W.; Champion, J.A.; Wang, B.Z. Double-layered protein nanoparticles induce broad protection against divergent influenza A viruses. Nat. Commun. 2018, 9, 359. [Google Scholar] [CrossRef]
- Song, J.M.; Van Rooijen, N.; Bozja, J.; Compans, R.W.; Kang, S.M. Vaccination inducing broad and improved cross protection against multiple subtypes of influenza A virus. Proc. Natl. Acad. Sci. USA 2011, 108, 757–761. [Google Scholar] [CrossRef]
- Bhatnagar, N.; Kim, K.-H.; Subbiah, J.; Park, B.R.; Wang, P.; Gill, H.S.; Wang, B.-Z.; Kang, S.-M. Adjuvant Effects of a New Saponin Analog VSA-1 on Enhancing Homologous and Heterosubtypic Protection by Influenza Virus Vaccination. Vaccines 2022, 10, 1383. [Google Scholar] [CrossRef] [PubMed]
- Chae, W.; Kim, P.; Hwang, B.J.; Seong, B.L. Universal monoclonal antibody-based influenza hemagglutinin quantitative enzyme-linked immunosorbent assay. Vaccine 2019, 37, 1457–1466. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.J.; Lee, Y.; Lee, Y.T.; Kim, Y.J.; Kim, K.H.; Kang, S.M. MPL and CpG combination adjuvants promote homologous and heterosubtypic cross protection of inactivated split influenza virus vaccine. Antivir. Res. 2018, 156, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Lee, Y.T.; Park, S.; Jung, Y.J.; Lee, Y.; Ko, E.J.; Kim, Y.J.; Li, X.; Kang, S.M. Neuraminidase expressing virus-like particle vaccine provides effective cross protection against influenza virus. Virology 2019, 535, 179–188. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Lee, Y.N.; Lee, Y.T.; Kim, M.C.; Gewirtz, A.T.; Kang, S.M. A Novel Vaccination Strategy Mediating the Induction of Lung-Resident Memory CD8 T Cells Confers Heterosubtypic Immunity against Future Pandemic Influenza Virus. J. Immunol. 2016, 196, 2637–2645. [Google Scholar] [CrossRef]
- Kim, M.C.; Lee, J.S.; Kwon, Y.M.; Eunju, O.; Lee, Y.J.; Choi, J.G.; Wang, B.Z.; Compans, R.W.; Kang, S.M. Multiple heterologous M2 extracellular domains presented on virus-like particles confer broader and stronger M2 immunity than live influenza A virus infection. Antivir. Res. 2013, 99, 328–335. [Google Scholar] [CrossRef]
- Thompson, M.G.; Naleway, A.; Fry, A.M.; Ball, S.; Spencer, S.M.; Reynolds, S.; Bozeman, S.; Levine, M.; Katz, J.M.; Gaglani, M. Effects of Repeated Annual Inactivated Influenza Vaccination among Healthcare Personnel on Serum Hemagglutinin Inhibition Antibody Response to A/Perth/16/2009 (H3N2)-like virus during 2010–2011. Vaccine 2016, 34, 981–988. [Google Scholar] [CrossRef]
- Song, J.Y.; Noh, J.Y.; Lee, J.S.; Wie, S.H.; Kim, Y.K.; Lee, J.; Jeong, H.W.; Kim, S.W.; Lee, S.H.; Park, K.H.; et al. Effectiveness of repeated influenza vaccination among the elderly population with high annual vaccine uptake rates during the three consecutive A/H3N2 epidemics. Vaccine 2020, 38, 318–322. [Google Scholar] [CrossRef]
- Belongia, E.A.; Skowronski, D.M.; McLean, H.Q.; Chambers, C.; Sundaram, M.E.; De Serres, G. Repeated annual influenza vaccination and vaccine effectiveness: Review of evidence. Expert Rev. Vaccines 2017, 16, 723–736. [Google Scholar] [CrossRef]
- Levine, M.Z.; Holiday, C.; Jefferson, S.; Gross, F.L.; Liu, F.; Li, S.; Friel, D.; Boutet, P.; Innis, B.L.; Mallett, C.P.; et al. Heterologous prime-boost with A(H5N1) pandemic influenza vaccines induces broader cross-clade antibody responses than homologous prime-boost. NPJ Vaccines 2019, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Van Reeth, K.; Gracia, J.C.M.; Trus, I.; Sys, L.; Claes, G.; Versnaeyen, H.; Cox, E.; Krammer, F.; Qiu, Y. Heterologous prime-boost vaccination with H3N2 influenza viruses of swine favors cross-clade antibody responses and protection. NPJ Vaccines 2017, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Stacey, H.D.; Mullarkey, C.E.; Miller, M.S. Original Antigenic Sin: How First Exposure Shapes Lifelong Anti-Influenza Virus Immune Responses. J. Immunol. 2019, 202, 335–340. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, C.D.; Wright, A.; Vogel, L.; Boonnak, K.; Treanor, J.J.; Subbarao, K. Humans and ferrets with prior H1N1 influenza virus infections do not exhibit evidence of original antigenic sin after infection or vaccination with the 2009 pandemic H1N1 influenza virus. Clin. Vaccine Immunol. CVI 2014, 21, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Nunzi, E.; Iorio, A.M.; Camilloni, B. A 21-winter seasons retrospective study of antibody response after influenza vaccination in elderly (60-85 years old) and very elderly (>85 years old) institutionalized subjects. Hum. Vaccin. Immunother. 2017, 13, 2659–2668. [Google Scholar] [CrossRef]
- Ng, S.; Nachbagauer, R.; Balmaseda, A.; Stadlbauer, D.; Ojeda, S.; Patel, M.; Rajabhathor, A.; Lopez, R.; Guglia, A.F.; Sanchez, N.; et al. Novel correlates of protection against pandemic H1N1 influenza A virus infection. Nat. Med. 2019, 25, 962–967. [Google Scholar] [CrossRef]
- Corbett, K.S.; Moin, S.M.; Yassine, H.M.; Cagigi, A.; Kanekiyo, M.; Boyoglu-Barnum, S.; Myers, S.I.; Tsybovsky, Y.; Wheatley, A.K.; Schramm, C.A.; et al. Design of Nanoparticulate Group 2 Influenza Virus Hemagglutinin Stem Antigens That Activate Unmutated Ancestor B Cell Receptors of Broadly Neutralizing Antibody Lineages. mBio 2019, 10, e02810-18. [Google Scholar] [CrossRef]
- Sutton, T.C.; Chakraborty, S.; Mallajosyula, V.V.A.; Lamirande, E.W.; Ganti, K.; Bock, K.W.; Moore, I.N.; Varadarajan, R.; Subbarao, K. Protective efficacy of influenza group 2 hemagglutinin stem-fragment immunogen vaccines. NPJ Vaccines 2017, 2, 35. [Google Scholar] [CrossRef]
- Impagliazzo, A.; Milder, F.; Kuipers, H.; Wagner, M.V.; Zhu, X.; Hoffman, R.M.; van Meersbergen, R.; Huizingh, J.; Wanningen, P.; Verspuij, J.; et al. A stable trimeric influenza hemagglutinin stem as a broadly protective immunogen. Science 2015, 349, 1301–1306. [Google Scholar] [CrossRef]
- Hai, R.; Krammer, F.; Tan, G.S.; Pica, N.; Eggink, D.; Maamary, J.; Margine, I.; Albrecht, R.A.; Palese, P. Influenza viruses expressing chimeric hemagglutinins: Globular head and stalk domains derived from different subtypes. J. Virol. 2012, 86, 5774–5781. [Google Scholar] [CrossRef]
- Krammer, F.; Margine, I.; Hai, R.; Flood, A.; Hirsh, A.; Tsvetnitsky, V.; Chen, D.; Palese, P. H3 stalk-based chimeric hemagglutinin influenza virus constructs protect mice from H7N9 challenge. J. Virol. 2014, 88, 2340–2343. [Google Scholar] [CrossRef] [PubMed]
- Nachbagauer, R.; Liu, W.C.; Choi, A.; Wohlbold, T.J.; Atlas, T.; Rajendran, M.; Solorzano, A.; Berlanda-Scorza, F.; Garcia-Sastre, A.; Palese, P.; et al. A universal influenza virus vaccine candidate confers protection against pandemic H1N1 infection in preclinical ferret studies. NPJ Vaccines 2017, 2, 26. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine Groups | Virus-Specific IgG (µg/mL) | HA Stalk-Specific IgG (µg/mL) | ||||||
|---|---|---|---|---|---|---|---|---|
| Prime | iPR8/H1N1 | iCal/H1N1 | iHK/H3N2 | iIndo/H5N1 | iSH/H7N9 | iHK/H9N2 | G1 Stalk | G2 Stalk |
| iPR8/H1N1 | 10.56 | 1.24 | 8.38 | 1.43 | 5.20 | 2.82 | 1.81 | 0.10 |
| iCal/H1N1 | 1.43 | 7.98 | 2.43 | 0.84 | 1.43 | 1.04 | 1.06 | 0.10 |
| iHK/H3N2 | 6.00 | 1.24 | 28.82 | 2.03 | 8.38 | 3.82 | 0.14 | 0.48 |
| iIndo/H5N1 | 6.00 | 1.04 | 9.97 | 15.13 | 7.98 | 4.21 | 0.36 | 0.12 |
| iSH/H7N9 | 4.41 | 1.04 | 8.78 | 1.83 | 10.36 | 3.02 | 0.14 | 0.64 |
| iHK/H9N2 | 3.02 | 1.43 | 5.40 | 0.84 | 1.83 | 8.38 | 0.36 | 0.18 |
| Naïve | 0.08 | 0.08 | 0.10 | 0.08 | 0.08 | 0.08 | 0.14 | 0.18 |
| Vaccine Groups | Virus-Specific IgG (µg/mL) | HA Stalk-Specific IgG (µg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Prime | Boost | iPR8/ H1N1 | iCal/ H1N1 | iHK/ H3N2 | iIndo/ H5N1 | iSH/ H7N9 | iHK/ H9N2 | G1 Stalk | G2 Stalk |
| iPR8/H1N1 | iPR8/H1N1 | 155.2 | 28.2 | 149.3 | 113.6 | 91.7 | 79.8 | 14.1 | 4.0 |
| iPR8/H1N1 | iCal/H1N1 | 79.8 | 34.2 | 103.6 | 85.8 | 67.9 | 50.1 | 20.9 | 3.6 |
| iCal/H1N1 | iPR8/H1N1 | 40.1 | 44.1 | 42.1 | 40.1 | 26.3 | 22.3 | 22.9 | 3.8 |
| iPR8/H1N1 | iHK/H3N2 | 105.6 | 30.2 | 250.5 | 161.2 | 121.5 | 73.9 | 3.8 | 10.4 |
| iHK/H3N2 | iPR8/H1N1 | 83.8 | 24.3 | 171.1 | 99.7 | 73.9 | 48.1 | 13.1 | 12.1 |
| iPR8/H1N1 | iIndo/H5N1 | 67.9 | 18.3 | 97.7 | 125.5 | 73.9 | 44.1 | 25.8 | 4.6 |
| iIndo/H5N1 | iPR8/H1N1 | 89.7 | 14.3 | 105.6 | 115.5 | 73.9 | 28.2 | 18.7 | 2.0 |
| iPR8/H1N1 | iSH/H7N9 | 99.7 | 30.2 | 210.8 | 177.0 | 131.4 | 44.1 | 8.4 | 9.2 |
| iSH/H7N9 | iPR8/H1N1 | 139.3 | 22.3 | 177.0 | 139.3 | 127.4 | 38.2 | 18.5 | 5.4 |
| iPR8/H1N1 | iHK/H9N2 | 77.8 | 24.3 | 196.9 | 121.5 | 87.8 | 58.0 | 3.8 | 4.0 |
| iHK/H9N2 | iPR8/H1N1 | 123.5 | 16.3 | 155.2 | 137.4 | 111.6 | 52.0 | 22.1 | 7.2 |
| Naïve | 0.05 | 0.10 | 0.08 | 0.10 | 0.10 | 0.08 | 0.08 | 0.14 | |
| Vaccine Groups | HAI Titers against Different Virus Strains | ||||||
|---|---|---|---|---|---|---|---|
| Prime | PR8/H1N1 | Cal/H1N1 | HK/H3N2 | Indo/H5N1 | SH/H7N9 | HK/H9N2 | Phil/H3N2 |
| iPR8/H1N1 | 1:64 | - | - | - | - | - | - |
| iCal/H1N1 | - | 1:64 | - | - | - | - | - |
| iHK/H3N2 | - | - | 1:256 | - | - | - | - |
| iIndo/H5N1 | - | - | - | 1:32 | - | - | - |
| iSH/H7N9 | - | - | - | - | 1:64 | - | - |
| iHK/H9N2 | - | - | - | - | - | 1:256 | - |
| Naïve | - | - | - | - | - | - | - |
| Vaccine Groups | HAI Titers against Different Virus Strains | |||||||
|---|---|---|---|---|---|---|---|---|
| Prime | Boost | PR8/ H1N1 | Cal/ H1N1 | HK/ H3N2 | Indo/ H5N1 | SH/ H7N9 | HK/ H9N2 | Phil/ H3N2 |
| iPR8/H1N1 | iPR8/H1N1 | 1:512 | - | - | - | - | - | - |
| iPR8/H1N1 | iCal/H1N1 | 1:256 | 1.64 | - | - | - | - | - |
| iCal/H1N1 | iPR8/H1N1 | 1:256 | 1:256 | - | - | - | - | - |
| iPR8/H1N1 | iHK/H3N2 | 1:256 | - | 1:1024 | - | - | - | - |
| iHK/H3N2 | iPR8/H1N1 | 1:256 | - | 1:512 | - | - | - | - |
| iPR8/H1N1 | iIndo/H5N1 | 1:256 | - | - | 1:64 | - | - | - |
| iIndo/H5N1 | iPR8/H1N1 | 1:512 | - | - | 1:64 | - | - | - |
| iPR8/H1N1 | iSH/H7N9 | 1:256 | - | - | - | 1:256 | - | - |
| iSH/H7N9 | iPR8/H1N1 | 1:512 | - | - | - | 1:512 | - | - |
| iPR8/H1N1 | iHK/H9N2 | 1:256 | - | - | - | - | 1:256 | - |
| iHK/H9N2 | iPR8/H1N1 | 1:512 | - | - | - | - | 1:512 | - |
| Naïve | - | - | - | - | - | - | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhatnagar, N.; Kim, K.-H.; Subbiah, J.; Muhammad-Worsham, S.; Park, B.R.; Liu, R.; Grovenstein, P.; Wang, B.-Z.; Kang, S.-M. Heterologous Prime-Boost Vaccination with Inactivated Influenza Viruses Induces More Effective Cross-Protection than Homologous Repeat Vaccination. Vaccines 2023, 11, 1209. https://doi.org/10.3390/vaccines11071209
Bhatnagar N, Kim K-H, Subbiah J, Muhammad-Worsham S, Park BR, Liu R, Grovenstein P, Wang B-Z, Kang S-M. Heterologous Prime-Boost Vaccination with Inactivated Influenza Viruses Induces More Effective Cross-Protection than Homologous Repeat Vaccination. Vaccines. 2023; 11(7):1209. https://doi.org/10.3390/vaccines11071209
Chicago/Turabian StyleBhatnagar, Noopur, Ki-Hye Kim, Jeeva Subbiah, Sakinah Muhammad-Worsham, Bo Ryoung Park, Rong Liu, Phillip Grovenstein, Bao-Zhong Wang, and Sang-Moo Kang. 2023. "Heterologous Prime-Boost Vaccination with Inactivated Influenza Viruses Induces More Effective Cross-Protection than Homologous Repeat Vaccination" Vaccines 11, no. 7: 1209. https://doi.org/10.3390/vaccines11071209
APA StyleBhatnagar, N., Kim, K.-H., Subbiah, J., Muhammad-Worsham, S., Park, B. R., Liu, R., Grovenstein, P., Wang, B.-Z., & Kang, S.-M. (2023). Heterologous Prime-Boost Vaccination with Inactivated Influenza Viruses Induces More Effective Cross-Protection than Homologous Repeat Vaccination. Vaccines, 11(7), 1209. https://doi.org/10.3390/vaccines11071209