
Increased Vaccination Diversity Leads to Higher and Less-Variable Neutralization of TBE Viruses of the European Subtype

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Sera
2.2. Virus Strains
2.3. Virus Cultivation Quantification and Microserum Neutralization Tests
2.4. Phylogenetic Analysis of the TBEV-EU Genome Sequences
2.5. Statistical Analysis
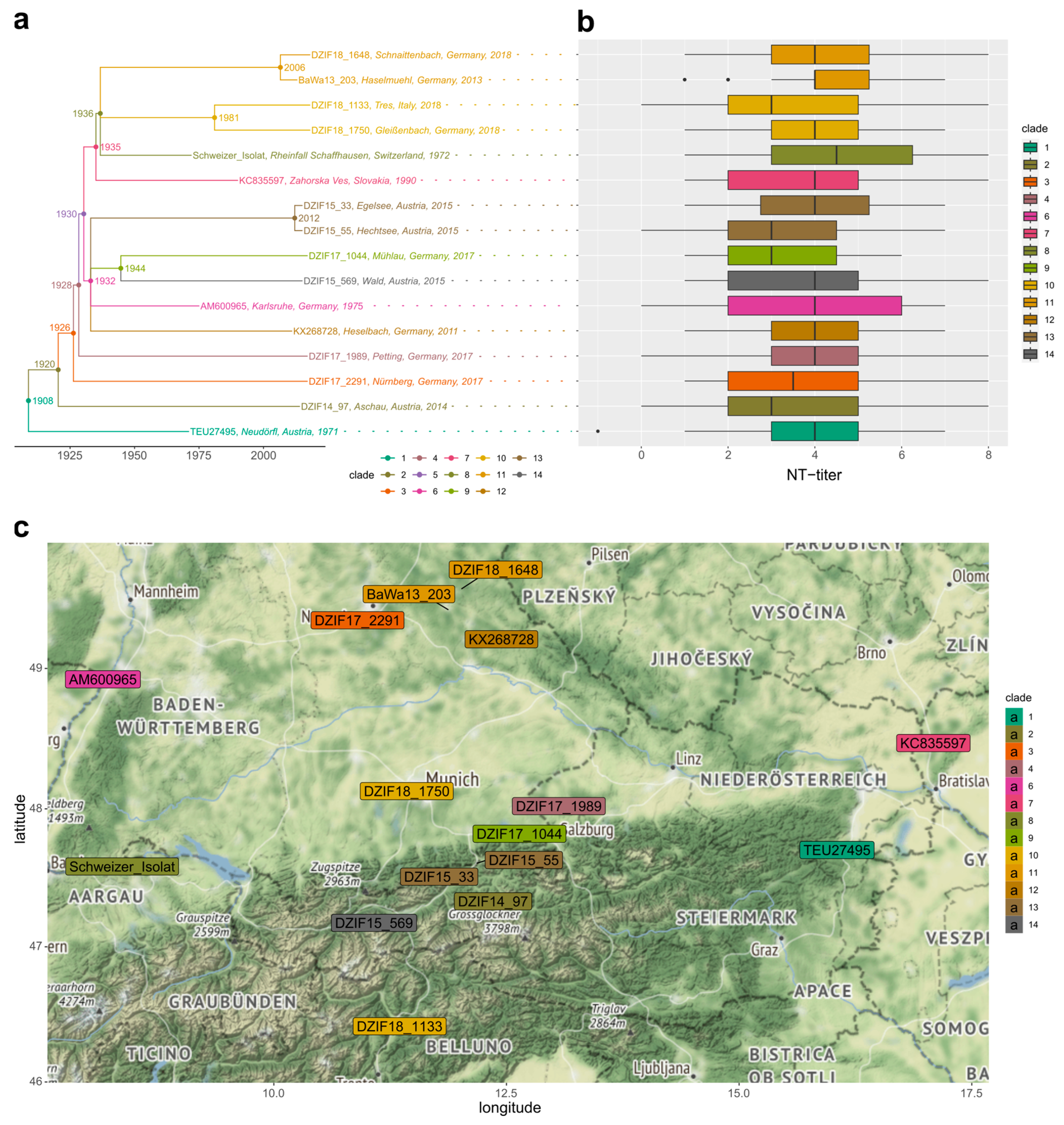
3. Results
3.1. High Inter-Serum Variation and Low Sampling Bias
3.2. Genotypes of the Diverse and Highly Divergent TBEV-EU Panel Were Neutralized Independent of Their Phylogenetic Position
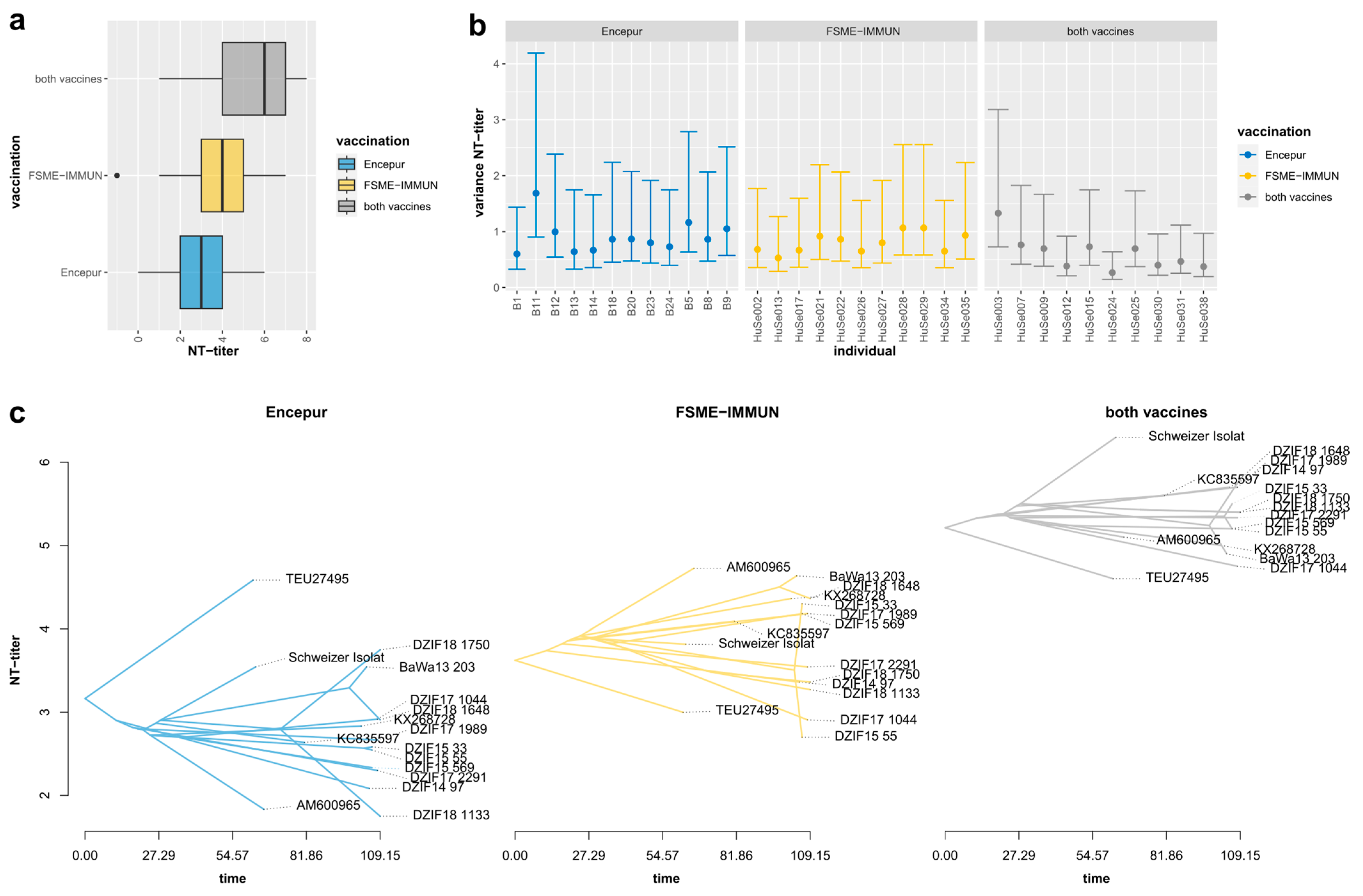
3.3. Neutralization Titers Differed Significantly, Depending on Serum Immunization Profiles
3.4. Sequence Analysis of Possible Genetic Determinants for the NT-Titer Phenoytpes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobler, G. Update Zur Frühsommer-Meningoenzephalitis. MMW-Fortschritte Med. 2020, 162, 40–43. [Google Scholar] [CrossRef]
- Boelke, M.; Bestehorn, M.; Marchwald, B.; Kubinski, M.; Liebig, K.; Glanz, J.; Schulz, C.; Dobler, G.; Monazahian, M.; Becker, S.C. First Isolation and Phylogenetic Analyses of Tick-Borne Encephalitis Virus in Lower Saxony, Germany. Viruses 2019, 11, 462. [Google Scholar] [CrossRef]
- Holding, M.; Dowall, S.D.; Medlock, J.M.; Carter, D.P.; Pullan, S.T.; Lewis, J.; Vipond, R.; Rocchi, M.S.; Baylis, M.; Hewson, R. Tick-Borne Encephalitis Virus, United Kingdom. Emerg. Infect. Dis. 2020, 26, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Camino, E.; Schmid, S.; Weber, F.; Pozo, P.; de Juan, L.; König, M.; Cruz-Lopez, F. Detection of Antibodies against Tick-Borne Encephalitis Flaviviruses in Breeding and Sport Horses from Spain. Ticks Tick-Borne Dis. 2020, 11, 101487. [Google Scholar] [CrossRef] [PubMed]
- Wallenhammar, A.; Lindqvist, R.; Asghar, N.; Gunaltay, S.; Fredlund, H.; Davidsson, Å.; Andersson, S.; Överby, A.K.; Johansson, M. Revealing New Tick-Borne Encephalitis Virus Foci by Screening Antibodies in Sheep Milk. Parasites Vectors 2020, 13, 185. [Google Scholar] [CrossRef] [PubMed]
- Brockmann, S.; Oehme, R.; Buckenmaier, T.; Beer, M.; Jeffery-Smith, A.; Spannenkrebs, M.; Haag-Milz, S.; Wagner-Wiening, C.; Schlegel, C.; Fritz, J.; et al. A Cluster of Two Human Cases of Tick-Borne Encephalitis (TBE) Transmitted by Unpasteurised Goat Milk and Cheese in Germany, May 2016. Eurosurveillance 2018, 23, 17-00336. [Google Scholar] [CrossRef]
- Holzmann, H.; Aberle, S.W.; Stiasny, K.; Werner, P.; Mischak, A.; Zainer, B.; Netzer, M.; Koppi, S.; Bechter, E.; Heinz, F.X. Tick-Borne Encephalitis from Eating Goat Cheese in a Mountain Region of Austria. Emerg. Infect. Dis. 2009, 15, 1671–1673. [Google Scholar] [CrossRef]
- Tick-Borne Encephalitis Outbreak Linked to Raw Milk Cheese in France. Available online: https://www.foodsafetynews.com/2020/06/tick-borne-encephalitis-outbreak-linked-to-raw-milk-cheese-in-france/ (accessed on 9 July 2020).
- Dai, X.; Shang, G.; Lu, S.; Yang, J.; Xu, J. A New Subtype of Eastern Tick-Borne Encephalitis Virus Discovered in Qinghai-Tibet Plateau, China. Emerg. Microbes Infect. 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Ecker, M.; Allison, S.L.; Meixner, T.; Heinz, F.X. Sequence Analysis and Genetic Classification of Tick-Borne Encephalitis Viruses from Europe and Asia. J. Gen. Virol. 1999, 80 Pt 1, 179–185. [Google Scholar] [CrossRef]
- Kovalev, S.; Mukhacheva, T.A. Reconsidering the Classification of Tick-Borne Encephalitis Virus within the Siberian Subtype Gives New Insights into Its Evolutionary History. Infect. Genet. Evol. 2017, 55, 159–165. [Google Scholar] [CrossRef]
- Bogovic, P.; Strle, F. Tick-Borne Encephalitis: A Review of Epidemiology, Clinical Characteristics, and Management. World J. Clin. Cases WJCC 2015, 3, 430–441. [Google Scholar] [CrossRef]
- Ruzek, D.; Avšič Županc, T.; Borde, J.; Chrdle, A.; Eyer, L.; Karganova, G.; Kholodilov, I.; Knap, N.; Kozlovskaya, L.; Matveev, A.; et al. Tick-Borne Encephalitis in Europe and Russia: Review of Pathogenesis, Clinical Features, Therapy, and Vaccines. Antiviral Res. 2019, 164, 23–51. [Google Scholar] [CrossRef]
- Dobler, G.; Gniel, D.; Petermann, R.; Pfeffer, M. Epidemiology and Distribution of Tick-Borne Encephalitis. Wien. Med. Wochenschr. 2012, 162, 230–238. [Google Scholar] [CrossRef]
- Kollaritsch, H.; Paulke-Korinek, M.; Holzmann, H.; Hombach, J.; Bjorvatn, B.; Barrett, A. Vaccines and Vaccination against Tick-Borne Encephalitis. Expert Rev. Vaccines 2012, 11, 1103–1119. [Google Scholar] [CrossRef]
- Gritsun, T.S.; Lashkevich, V.A.; Gould, E.A. Tick-Borne Encephalitis. Antivir. Res. 2003, 57, 129–146. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Johnson, N.; Phipps, L.P.; Stephenson, J.R.; Fooks, A.R.; Solomon, T. Tick-Borne Encephalitis Virus—A Review of an Emerging Zoonosis. J. Gen. Virol. 2009, 90, 1781–1794. [Google Scholar] [CrossRef]
- Lang, D.; Chitimia-Dobler, L.; Bestehorn-Willmann, M.; Lindau, A.; Drehmann, M.; Stroppel, G.; Hengge, H.; Mackenstedt, U.; Kaier, K.; Dobler, G.; et al. The Emergence and Dynamics of Tick-Borne Encephalitis Virus in a New Endemic Region in Southern Germany. Microorganisms 2022, 10, 2125. [Google Scholar] [CrossRef]
- Kutschera, L.S.; Wolfinger, M.T. Evolutionary Traits of Tick-Borne Encephalitis Virus: Pervasive Non-Coding RNA Structure Conservation and Molecular Epidemiology. Virus Evol. 2022, 8, veac051. [Google Scholar] [CrossRef]
- WHO|Vaccine Position Papers. Available online: http://www.who.int/immunization/documents/positionpapers/en/ (accessed on 30 July 2020).
- Klockmann, U.; Krivanec, K.; Stephenson, J.; Hilfenhaus, J. Protection against European Isolates of Tick-Borne Encephalitis Virus after Vaccination with a New Tick-Borne Encephalitis Vaccine. Vaccine 1991, 9, 210–212. [Google Scholar] [CrossRef]
- Mandl, C.W.; Heinz, F.X.; Stöckl, E.; Kunz, C. Genome Sequence of Tick-Borne Encephalitis Virus (Western Subtype) and Comparative Analysis of Nonstructural Proteins with Other Flaviviruses. Virology 1989, 173, 291–301. [Google Scholar] [CrossRef]
- Wallner, G.; Mandl, C.W.; Kunz, C.; Heinz, F.X. The Flavivirus 3’-Noncoding Region: Extensive Size Heterogeneity Independent of Evolutionary Relationships among Strains of Tick-Borne Encephalitis Virus. Virology 1995, 213, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K.L.; Horton, D.L.; Johnson, N.; Li, L.; Barrett, A.D.T.; Smith, D.J.; Galbraith, S.E.; Solomon, T.; Fooks, A.R. Flavivirus-Induced Antibody Cross-Reactivity. J. Gen. Virol. 2011, 92, 2821–2829. [Google Scholar] [CrossRef]
- Saron, W.A.A.; Rathore, A.P.S.; Ting, L.; Ooi, E.E.; Low, J.; Abraham, S.N.; John, A.L.S. Flavivirus Serocomplex Cross-Reactive Immunity Is Protective by Activating Heterologous Memory CD4 T Cells. Sci. Adv. 2018, 4, eaar4297. [Google Scholar] [CrossRef]
- Antigenic Variation among Members of the Tick-Borne Encephalitis Complex|Microbiology Society. Available online: https://www.microbiologyresearch.org/content/journal/jgv/10.1099/0022-1317-65-1-81;jsessionid=s_-q09J2Y6hqMFua3VCKkV2E.mbslive-10-240-10-152 (accessed on 1 February 2021).
- McAuley, A.J.; Sawatsky, B.; Ksiazek, T.; Torres, M.; Korva, M.; Lotrič-Furlan, S.; Avšič-Županc, T.; von Messling, V.; Holbrook, M.R.; Freiberg, A.N.; et al. Cross-Neutralisation of Viruses of the Tick-Borne Encephalitis Complex Following Tick-Borne Encephalitis Vaccination and/or Infection. NPJ Vaccines 2017, 2, 5. [Google Scholar] [CrossRef]
- Mandl, C.W.; Holzmann, H.; Meixner, T.; Rauscher, S.; Stadler, P.F.; Allison, S.L.; Heinz, F.X. Spontaneous and Engineered Deletions in the 3′ Noncoding Region of Tick-Borne Encephalitis Virus: Construction of Highly Attenuated Mutants of a Flavivirus. J. Virol. 1998, 72, 2132–2140. [Google Scholar] [CrossRef]
- Beck, Y.; Fritz, R.; Orlinger, K.; Kiermayr, S.; Ilk, R.; Portsmouth, D.; Pöllabauer, E.-M.; Löw-Baselli, A.; Hessel, A.; Kölch, D.; et al. Molecular Basis of the Divergent Immunogenicity of Two Pediatric Tick-Borne Encephalitis Virus Vaccines. J. Virol. 2016, 90, 1964–1972. [Google Scholar] [CrossRef]
- Rockstroh, A.; Moges, B.; Berneck, B.S.; Sattler, T.; Revilla-Fernández, S.; Schmoll, F.; Pacenti, M.; Sinigaglia, A.; Barzon, L.; Schmidt-Chanasit, J.; et al. Specific Detection and Differentiation of Tick-borne Encephalitis and West Nile Virus Induced IgG Antibodies in Humans and Horses. Transbound. Emerg. Dis. 2019, 66, 1701–1708. [Google Scholar] [CrossRef]
- Bradt, V.; Malafa, S.; von Braun, A.; Jarmer, J.; Tsouchnikas, G.; Medits, I.; Wanke, K.; Karrer, U.; Stiasny, K.; Heinz, F.X. Pre-Existing Yellow Fever Immunity Impairs and Modulates the Antibody Response to Tick-Borne Encephalitis Vaccination. NPJ Vaccines 2019, 4, 38. [Google Scholar] [CrossRef]
- Dobler, G.; Bestehorn, M.; Antwerpen, M.; Överby-Wernstedt, A. Complete Genome Sequence of a Low-Virulence Tick-Borne Encephalitis Virus Strain. Genome Announc. 2016, 4, e01145-16. [Google Scholar] [CrossRef]
- Weidmann, M.; Frey, S.; Freire, C.C.M.; Essbauer, S.; Růžek, D.; Klempa, B.; Zubrikova, D.; Vögerl, M.; Pfeffer, M.; Hufert, F.T.; et al. Molecular Phylogeography of Tick-Borne Encephalitis Virus in Central Europe. J. Gen. Virol. 2013, 94, 2129–2139. [Google Scholar] [CrossRef]
- Baer, A.; Kehn-Hall, K. Viral Concentration Determination Through Plaque Assays: Using Traditional and Novel Overlay Systems. J. Vis. Exp. JoVE 2014, 93, e52065. [Google Scholar] [CrossRef]
- Haut, M.; Girl, P.; Oswald, B.; Romig, T.; Obiegala, A.; Dobler, G.; Pfeffer, M. The Red Fox (Vulpes vulpes) as Sentinel for Tick-Borne Encephalitis Virus in Endemic and Non-Endemic Areas. Microorganisms 2020, 8, 1817. [Google Scholar] [CrossRef] [PubMed]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.A.; Korobeynikov, A.; Lapidus, A.; Prjibelski, A.D.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling Single-Cell Genomes and Mini-Metagenomes From Chimeric MDA Products. J. Comput. Biol. 2013, 20, 714–737. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Yamada, K.D.; Tomii, K.; Katoh, K. Parallelization of MAFFT for Large-Scale Multiple Sequence Alignments. Bioinformatics 2018, 34, 2490–2492. [Google Scholar] [CrossRef]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A Fast, Scalable and User-Friendly Tool for Maximum Likelihood Phylogenetic Inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef]
- Sagulenko, P.; Puller, V.; Neher, R.A. TreeTime: Maximum-Likelihood Phylodynamic Analysis. Virus Evol. 2018, 4, vex042. [Google Scholar] [CrossRef]
- Revell, L.J. Phytools: An R Package for Phylogenetic Comparative Biology (and Other Things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. Ape 5.0: An Environment for Modern Phylogenetics and Evolutionary Analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Yu, G. Data Integration, Manipulation and Visualization of Phylogenetic Treess, 1st ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2022. [Google Scholar]
- Yu, G. Using Ggtree to Visualize Data on Tree-Like Structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef]
- Wang, L.-G.; Lam, T.T.-Y.; Xu, S.; Dai, Z.; Zhou, L.; Feng, T.; Guo, P.; Dunn, C.W.; Jones, B.R.; Bradley, T.; et al. Treeio: An R Package for Phylogenetic Tree Input and Output with Richly Annotated and Associated Data. Mol. Biol. Evol. 2020, 37, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Kahle, D.; Wickham, H. Ggmap: Spatial Visualization with Ggplot2. R J. 2013, 5, 144–161. [Google Scholar] [CrossRef]
- Padgham, M.; Rudis, B.; Lovelace, R.; Salmon, M. Osmdata. J. Open Source Softw. 2017, 2, 1. [Google Scholar] [CrossRef]
- Blomberg, S.P.; Garland, T.; Ives, A.R. Testing for Phylogenetic Signal in Comparative Data: Behavioral Traits Are More Labile. Evol. Int. J. Org. Evol. 2003, 57, 717–745. [Google Scholar] [CrossRef]
- Pennell, M.W.; Eastman, J.M.; Slater, G.J.; Brown, J.W.; Uyeda, J.C.; FitzJohn, R.G.; Alfaro, M.E.; Harmon, L.J. Geiger v2.0: An Expanded Suite of Methods for Fitting Macroevolutionary Models to Phylogenetic Trees. Bioinformatics 2014, 30, 2216–2218. [Google Scholar] [CrossRef]
- Kayser, M.; Klein, H.; Paasch, I.; Pilaski, J.; Blenk, H.; Heeg, K. Human Antibody Response to Immunization with 17D Yellow Fever and Inactivated TBE Vaccine. J. Med. Virol. 1985, 17, 35–45. [Google Scholar] [CrossRef]
- Garner-Spitzer, E.; Poellabauer, E.-M.; Wagner, A.; Guzek, A.; Zwazl, I.; Seidl-Friedrich, C.; Binder, C.J.; Stiasny, K.; Kundi, M.; Wiedermann, U. Obesity and Sex Affect the Immune Responses to Tick-Borne Encephalitis Booster Vaccination. Front. Immunol. 2020, 11, 860. [Google Scholar] [CrossRef]
- Janik, M.; Płaczkowska, S.; Woźniak, M.; Bil-Lula, I. Analysis of Multiple Risk Factors for Seronegative Rate of Anti-Tick-Borne Encephalitis Virus Immunization in Human Serum. Medicina 2020, 56, 244. [Google Scholar] [CrossRef]
- Baldovin, T.; Mel, R.; Bertoncello, C.; Carpenè, G.; Soppelsa, F.; Giliberti, A.; Baldo, V. Persistence of Immunity to Tick-Borne Encephalitis after Vaccination and Natural Infection. J. Med. Virol. 2012, 84, 1274–1278. [Google Scholar] [CrossRef]
- Hansson, K.E.; Rosdahl, A.; Insulander, M.; Vene, S.; Lindquist, L.; Gredmark-Russ, S.; Askling, H.H. Tick-Borne Encephalitis Vaccine Failures: A 10-Year Retrospective Study Supporting the Rationale for Adding an Extra Priming Dose in Individuals Starting at Age 50 Years. Clin. Infect. Dis. 2020, 70, 245–251. [Google Scholar] [CrossRef]
- Stiasny, K.; Aberle, J.; Keller, M.; Grubeck-Loebenstein, B.; Heinz, F.X. Age Affects Quantity but Not Quality of Antibody Responses after Vaccination with an Inactivated Flavivirus Vaccine against Tick-Borne Encephalitis. PLoS ONE 2012, 7, e34145. [Google Scholar] [CrossRef] [PubMed]
- Nygren, T.M.; Pilic, A.; Böhmer, M.M.; Wagner-Wiening, C.; Wichmann, O.; Harder, T.; Hellenbrand, W. Tick-Borne Encephalitis Vaccine Effectiveness and Barriers to Vaccination in Germany. Sci. Rep. 2022, 12, 11706. [Google Scholar] [CrossRef] [PubMed]
- Heinz, F.X.; Holzmann, H.; Essl, A.; Kundi, M. Field Effectiveness of Vaccination against Tick-Borne Encephalitis. Vaccine 2007, 25, 7559–7567. [Google Scholar] [CrossRef]
- Erber, W.; Khan, F.; Zavadska, D.; Freimane, Z.; Dobler, G.; Böhmer, M.M.; Jodar, L.; Schmitt, H.-J. Effectiveness of TBE Vaccination in Southern Germany and Latvia. Vaccine 2022, 40, 819–825. [Google Scholar] [CrossRef]
- Askling, H.H.; Vene, S.; Rombo, L.; Lindquist, L. Immunogenicity of Delayed TBE-Vaccine Booster. Vaccine 2012, 30, 499–502. [Google Scholar] [CrossRef]
- Steffen, R. Tick-Borne Encephalitis (TBE) in Children in Europe: Epidemiology, Clinical Outcome and Comparison of Vaccination Recommendations. Ticks Tick-Borne Dis. 2019, 10, 100–110. [Google Scholar] [CrossRef]
- Salat, J.; Mikulasek, K.; Larralde, O.; Pokorna Formanova, P.; Chrdle, A.; Haviernik, J.; Elsterova, J.; Teislerova, D.; Palus, M.; Eyer, L.; et al. Tick-Borne Encephalitis Virus Vaccines Contain Non-Structural Protein 1 Antigen and May Elicit NS1-Specific Antibody Responses in Vaccinated Individuals. Vaccines 2020, 8, 81. [Google Scholar] [CrossRef]
- Loew-Baselli, A.; Konior, R.; Pavlova, B.G.; Fritsch, S.; Poellabauer, E.; Maritsch, F.; Harmacek, P.; Krammer, M.; Barrett, P.N.; Ehrlich, H.J. Safety and Immunogenicity of the Modified Adult Tick-Borne Encephalitis Vaccine FSME-IMMUN®: Results of Two Large Phase 3 Clinical Studies. Vaccine 2006, 24, 5256–5263. [Google Scholar] [CrossRef]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological Memory to SARS-CoV-2 Assessed for up to 8 Months after Infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef]
- Bröker, M.; Schöndorf, I. Are Tick-Borne Encephalitis Vaccines Interchangeable? Expert Rev. Vaccines 2006, 5, 461–466. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sera | Vaccination | Count | Age | Sex | YF Vaccination |
|---|---|---|---|---|---|
| HuSe002 | FSME-IMMUN® | 5 | 30–40 | Male | Vaccinated |
| HuSe013 | FSME-IMMUN® | 3 | 20–30 | Male | No vaccination |
| HuSe017 | FSME-IMMUN® | 3 | 40–60 | Female | No vaccination |
| HuSe021 | FSME-IMMUN® | 3 | 20–30 | Female | No vaccination |
| HuSe022 | FSME-IMMUN® | 4 | 40–60 | Female | Vaccinated |
| HuSe026 | FSME-IMMUN® | 5 | 30–40 | Male | Vaccinated |
| HuSe027 | FSME-IMMUN® | 6 | 30–40 | Female | No vaccination |
| HuSe028 | FSME-IMMUN® | 6 | 30–40 | Female | No vaccination |
| HuSe029 | FSME-IMMUN® | 5 | 20–30 | Male | Vaccinated |
| HuSe035 | FSME-IMMUN® | 3 | 20–30 | Female | No vaccination |
| HuSe034 | FSME-IMMUN® | 6 | 40–60 | Female | Vaccinated |
| HuSe003 | MIX | 5 | 40–60 | Male | Vaccinated |
| HuSe007 | MIX | 5 | 30–40 | Female | Vaccinated |
| HuSe009 | MIX | 6 | 30–40 | Female | Vaccinated |
| HuSe012 | MIX | 5 | 30–40 | Female | No vaccination |
| HuSe015 | MIX | 5 | 20–30 | Male | No vaccination |
| HuSe024 | MIX | 5 | 20–30 | Female | No vaccination |
| HuSe025 | MIX | 5 | 40–60 | Female | No vaccination |
| HuSe030 | MIX | 7 | 30–40 | Male | No vaccination |
| HuSe031 | MIX | 3 | 30–40 | Female | Vaccinated |
| HuSe038 | MIX | 4 | 20–30 | Male | No vaccination |
| B1 | Encepur® | 4 | 20–30 | Female | No vaccination |
| B5 | Encepur® | 3 | 20–30 | Female | No vaccination |
| B8 | Encepur® | 5 | 20–30 | Male | No vaccination |
| B9 | Encepur® | 4 | 20–30 | Female | No vaccination |
| B11 | Encepur® | 5 | 20–30 | Female | No vaccination |
| B12 | Encepur® | 4 | 20–30 | Female | No vaccination |
| B13 | Encepur® | 6 | 20–30 | Female | No vaccination |
| B14 | Encepur® | 4 | 20–30 | Male | No vaccination |
| B18 | Encepur® | 6 | 20–30 | Female | No vaccination |
| B20 | Encepur® | 4 | 20–30 | Male | No vaccination |
| B23 | Encepur® | 4 | 20–30 | Female | No vaccination |
| B24 | Encepur® | 5 | 40–60 | Female | No vaccination |
| Strain ID | Acc. | Country | Region/City | Year of Isolation | Isolation Source | Passage History | Reference | Sequencing Method |
|---|---|---|---|---|---|---|---|---|
| K2 (Karlsruhe); K23 | AM600965 | Germany | Karlsruhe | 1975 | Ixodes ricinus | 4. BMB 1. A549 | [21] provided by FX Heinz, Vienna, Austria | Amplicon sequencing with Illumina TruSeq |
| Neudörfl | TEU27495 | Austria | Neudoerfl | 1971 | Ixodes ricinus | Unknown | [22,23] | Amplicon sequencing with Illumina TruSeq |
| DZIF14 97 | Austria | Aschau/Zillertal | 2014 | Ixodes ricinus | A549 Passage 01 | Isolate of IMB | Amplicon sequencing with Illumina TruSeq | |
| DZIF15 33 | Germany/Austria | Eglsee/Kufstein | 2015 | Ixodes ricinus | A549 Passage 01 | Isolate of IMB | Amplicon sequencing with Illumina TruSeq | |
| DZIF15 569 | Austria | Wald/Pitztal | 2015 | Ixodes ricinus | A549 Passage 01 | Isolate of IMB | Amplicon sequencing with Illumina TruSeq | |
| DZIF17 1044 | Germany | Mühlau | 2017 | Ixodes ricinus | A549 Passage 01 | Isolate of IMB | Amplicon sequencing with Illumina TruSeq | |
| DZIF17 1989 | Germany | Petting | 2017 | Ixodes ricinus | A549 Passage 01 | Isolate of IMB | Amplicon sequencing with Illumina TruSeq | |
| DZIF18 1133 | Italy | Tres | 2018 | Ixodes ricinus | A549 Passage 01 | Isolate of IMB | Amplicon sequencing with Illumina TruSeq | |
| BaWa11 171 | KX268728 | Germany | Heselbach | 2011 | Ixodes ricinus | A549 Passage 01 | [32] | Sanger sequencing |
| Schweizer_Isolat (40) | Switzerland | Schaffhausen | 1972 | Ixodes ricinus | 1. BMB 1. A549 | provided by Franz X. Heinz | Amplicon sequencing with Illumina TruSeq | |
| BaWa13 203 | Germany | Haselmühl | 2013 | Ixodes ricinus | A549 Passage 01 | Isolate of IMB | Amplicon sequencing with Illumina TruSeq | |
| DZIF17 2291 | Germany | Nürnberg | 2017 | Ixodes ricinus | A549 Passage 01 | Isolate of IMB | Amplicon sequencing with Illumina TruSeq | |
| DZIF18 1750 | Germany | Gleissenbach | 2018 | Ixodes ricinus | A549 Passage 01 | Isolate of IMB | Amplicon sequencing with Illumina TruSeq | |
| DZIF18 1648 | Germany | Schnaittenbach | 2018 | Ixodes ricinus | A549 Passage 01 | Isolate of IMB | Amplicon sequencing with Illumina TruSeq | |
| CG/223_1990 | KC835597 | Slovakia | Zahorska Ves | 1990 | Myodes glareolus | 5. BMB 3. Vero 1. A549 | [33] | Sanger sequencing |
| DZIF15 55 | Germany/Austria | Hechtsee | 2015 | Ixodes ricinus | A549 Passage 01 | Isolate of IMB | Amplicon sequencing with Illumina TruSeq |
| Neutralization Titers (Mean ± SD)/10 LOG2 | |
|---|---|
| Sex of the sample donors | |
| Female | n = 22 (66%) 3.79 ± 1.85 |
| Male | n = 11 (33%) 4.17 ± 1.99 |
| Wilcoxon Test | p = 0.063 |
| Yellow fever vaccination | |
| YF+ | n = 9 (27%) 3.8 ± 1.41 |
| YF− | n = 24 (73%) 3.96 ± 2.06 |
| Wilcoxon Test | p = 0.79 |
| Number of Vaccinations (nV) | |
| 3 | n = 6 (18%) 3.83 ± 1.68 |
| 4 | n = 8 (24%) 3.37 ± 1.93 |
| 5 | n = 12 (36%) 4.19 ± 1.98 |
| 6 | n = 6 (18%) 3.81 ± 1.73 |
| 7 | n = 1 (4%) 6 |
| Kendall’s rank correlation test | R = 0.109, p = 0.0018 ** |
| Generalized linear model | nV-coefficient = 0.2393 p = 0.00212 ** |
| Immunization profile | |
| Encepur | n = 12 (36.3%) 2.8 ± 1.43 |
| FSME-IMMUN | n = 11 (33.3%) 3.8 ± 1.62 |
| MIX | n = 10 (30.3%) 5.35 ± 1.74 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bestehorn-Willmann, M.; Girl, P.; Greiner, F.; Mackenstedt, U.; Dobler, G.; Lang, D. Increased Vaccination Diversity Leads to Higher and Less-Variable Neutralization of TBE Viruses of the European Subtype. Vaccines 2023, 11, 1044. https://doi.org/10.3390/vaccines11061044
Bestehorn-Willmann M, Girl P, Greiner F, Mackenstedt U, Dobler G, Lang D. Increased Vaccination Diversity Leads to Higher and Less-Variable Neutralization of TBE Viruses of the European Subtype. Vaccines. 2023; 11(6):1044. https://doi.org/10.3390/vaccines11061044
Chicago/Turabian StyleBestehorn-Willmann, Malena, Philipp Girl, Franziska Greiner, Ute Mackenstedt, Gerhard Dobler, and Daniel Lang. 2023. "Increased Vaccination Diversity Leads to Higher and Less-Variable Neutralization of TBE Viruses of the European Subtype" Vaccines 11, no. 6: 1044. https://doi.org/10.3390/vaccines11061044
APA StyleBestehorn-Willmann, M., Girl, P., Greiner, F., Mackenstedt, U., Dobler, G., & Lang, D. (2023). Increased Vaccination Diversity Leads to Higher and Less-Variable Neutralization of TBE Viruses of the European Subtype. Vaccines, 11(6), 1044. https://doi.org/10.3390/vaccines11061044