
Development of an Inactivated Avian Influenza Virus Vaccine against Circulating H9N2 in Chickens and Ducks

Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses and Animals
2.2. Preparation of Vaccine Candidate
2.3. Antibody Responses after Immunization
2.4. Protective Efficacy
2.5. Determination of Minimum Protective Dose
2.6. Cross-Protective Efficacy of Inactivated Vaccine against H9N2 Viruses
2.7. Field Application
2.8. Detection of Egg Yolk Antibody and Maternal Antibody
2.9. Statistical Analysis
3. Results
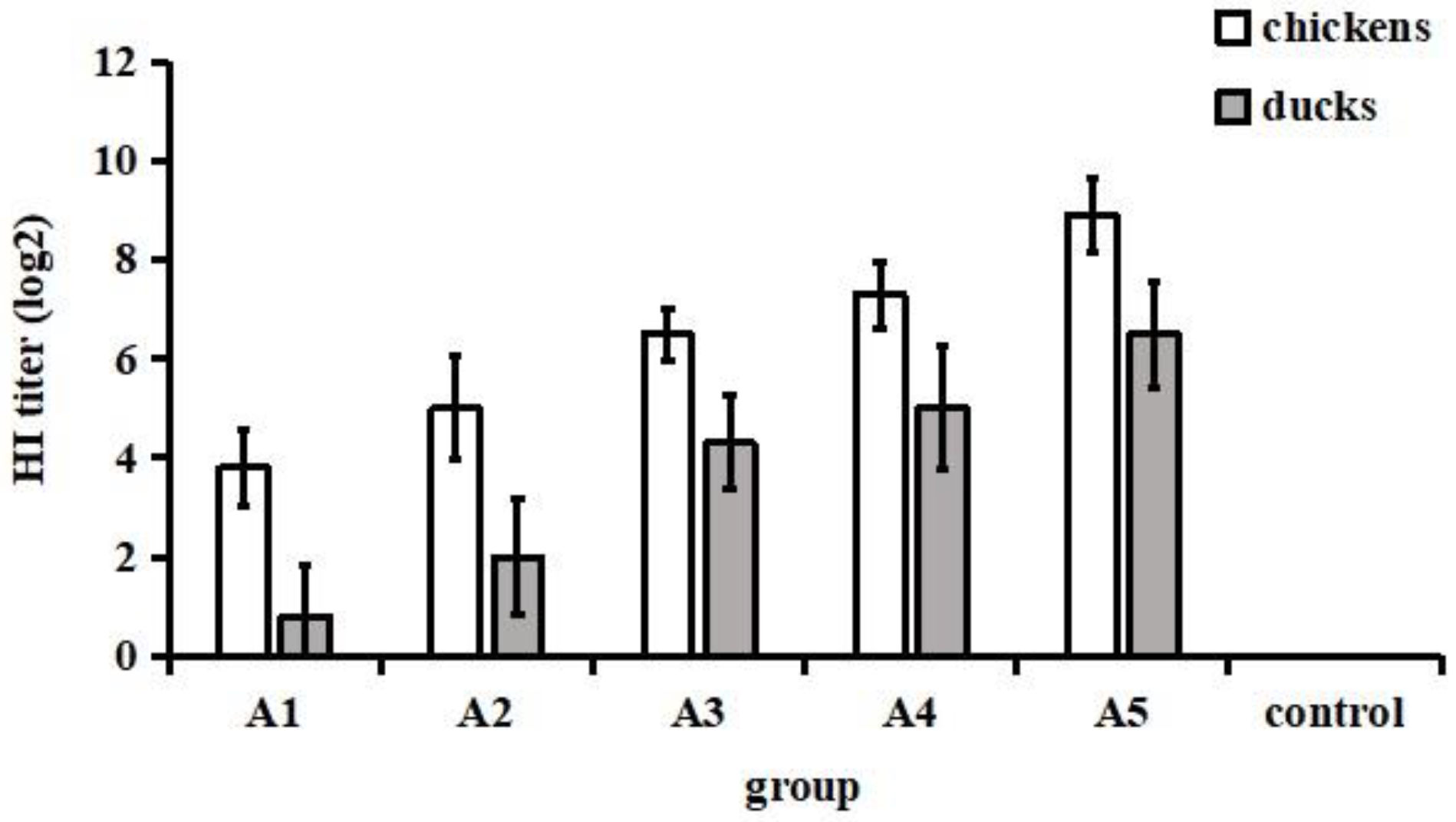
3.1. Antibody Responses after Immunization
3.2. Protective Efficacy after Viral Challenge
3.3. Determination of Minimum Protective Dose
3.4. Cross-Protective Efficacy of Inactivated Vaccine against Heterologous H9N2 Viruses
3.5. Field Application
3.6. Determination of Egg Yolk Antibody and Maternal Antibody
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang:, Y.; Zou, S.M.; Li, X.D.; Dong, L.B.; Bo, H.; Gao, R.B.; Wang, D.Y.; Shu, Y.L. Detection of reassortant avian influenza A (H11N9) virus in environmental samples from live poultry markets in China. Infect. Dis. Poverty 2016, 5, 59. [Google Scholar] [CrossRef][Green Version]
- Kajihara, M.; Sakoda, Y.; Soda, K.; Minari, K.; Okamatsu, M.; Takada, A.; Kida, H. The PB2, PA, HA, NP, and NS genes of a highly pathogenic avian influenza virus A/whooper swan/Mongolia/3/2005 (H5N1) are responsible for pathogenicity in ducks. Virol. J. 2013, 10, 45. [Google Scholar] [CrossRef]
- Mitrasinovic, P.M. Advances in the structure-based design of the influenza A neuraminidase inhibitors. Curr. Drug Targets 2010, 11, 315–326. [Google Scholar] [CrossRef]
- Sultankulova, K.T.; Kozhabergenov, N.S.; Strochkov, V.M.; Burashev, Y.D.; Shorayeva, K.A.; Chervyakova, O.V.; Rametov, N.M.; Sandybayev, N.T.; Sansyzbay, A.R.; Orynbayev, M.B. New oligonucleotide microarray for rapid diagnosis of avian viral diseases. Virol. J. 2017, 14, 69. [Google Scholar] [CrossRef]
- Rebel, J.M.; Peeters, B.; Fijten, H.; Post, J.; Cornelissen, J.; Vervelde, L. Highly pathogenic or low pathogenic avian influenza virus subtype H7N1 infection in chicken lungs: Small differences in general acute responses. Vet. Res. 2011, 42, 10. [Google Scholar] [CrossRef]
- Dupré, G.; Hoede, C.; Figueroa, T.; Bessière, P.; Bertagnoli, S.; Ducatez, M.; Gaspin, C.; Volmer, R. Phylodynamic study of the conserved RNA structure encompassing the hemagglutinin cleavage site encoding region of H5 and H7 low pathogenic avian influenza viruses. Virus Evol. 2021, 7, veab093. [Google Scholar] [CrossRef]
- Wang, J.; Li, C.C.; Diao, Y.X.; Sun, X.Y.; Hao, D.M.; Liu, X.; Ge, P.P. Different outcomes of infection of chickens and ducks with a duck-origin H9N2 influenza A virus. Acta Virol. 2014, 58, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Mettenleiter, T.C.; Abdelwhab, E.M. A brief summary of the epidemiology and genetic relatedness of avian influenza H9N2 virus in birds and mammals in the Middle East and North Africa. Epidemiol. Infect. 2017, 145, 3320–3333. [Google Scholar] [CrossRef] [PubMed]
- Perdue, M.L.; Swayne, D.E. Public health risk from avian influenza viruses. Avian Dis. 2005, 49, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Lai, H.Z.; Li, L.; Liu, Y.P.; Zhang, W.Y.; Gao, R.; Huang, W.K.; Luo, Q.F.; Gao, Y.; Luo, Q.; et al. Endemic variation of H9N2 avian influenza virus in China. Avian Dis. 2016, 60, 817–825. [Google Scholar] [CrossRef]
- Dong, J.; Zhou, Y.; Pu, J.; Liu, L. Status and challenges for vaccination against avian H9N2 influenza virus in China. Life 2022, 12, 1326. [Google Scholar] [CrossRef]
- Song, W.; Qin, K. Human-infecting influenza A (H9N2) virus: A forgotten potential pandemic strain? Zoonoses Public Health 2020, 67, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Shi, W.; Shi, Y.; Wang, D.; Xiao, H.; Li, W.; Bi, Y.; Wu, Y.; Li, X.; Yan, J.; et al. Origin and diversity of novel avian influenza A H7N9 viruses causing human infection: Phylogenetic, structural, and coalescent analyses. Lancet 2013, 381, 1926–1932. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Shortridge, K.F.; Krauss, S.; Webster, R.G. Molecular characterization of H9N2 influenza viruses: Were they the donors of the “internal” genes of H5N1 viruses in Hong Kong? Proc. Natl. Acad. Sci. USA 1999, 96, 9363–9367. [Google Scholar] [CrossRef]
- Roche, B.; Lebarbenchon, C.; Gauthier-Clerc, M.; Chang, C.M.; Thomas, F.; Renaud, F.; van der Werf, S.; Guégan, J.F. Water-borne transmission drives avian influenza dynamics in wild birds: The case of the 2005–2006 epidemics in the Camargue area. Infect. Genet. Evol. 2009, 9, 800–805. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Lai, H.; Xu, J.; Huang, W.; Liu, Y.; Zhao, D.; Chen, R. Evaluation of the protective efficacy of poly I:C as an adjuvant for H9N2 subtype avian influenza inactivated vaccine and its mechanism of action in ducks. PLoS ONE 2017, 12, e0170681. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, C.; Li, J.; Li, L.; Cao, M.; Li, Q.; Li, H. Construction of a recombinant duck enteritis virus vaccine expressing hemagglutinin of H9N2 avian influenza virus and evaluation of its efficacy in ducks. Arch. Virol. 2017, 162, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Zhao, J.; Yang, H.; Xu, H.; Ma, Z.; Zhang, G. Efficacy of an inactivated bivalent vaccine against the prevalent strains of Newcastle disease and H9N2 avian influenza. Virol. J. 2017, 14, 56. [Google Scholar] [CrossRef]
- Pu, J.; Wang, S.; Yin, Y.; Zhang, G.; Carter, R.A.; Wang, J.; Xu, G.; Sun, H.; Wang, M.; Wen, C.; et al. Evolution of the H9N2 influenza genotype that facilitated the genesis of the novel H7N9 virus. Proc. Natl. Acad. Sci. USA 2015, 112, 548–553. [Google Scholar] [CrossRef]
- Bi, Y.; Li, J.; Li, S.; Fu, G.; Jin, T.; Zhang, C.; Yang, Y.; Ma, Z.; Tian, W.; Li, J.; et al. Dominant subtype switch in avian influenza viruses during 2016-2019 in China. Nat. Commun. 2020, 11, 5909. [Google Scholar] [CrossRef]
- Shao, G.; Xie, Z.; Liang, M.; Liu, Y.; Song, C.; Feng, K.; Zhang, X.; Lin, W.; Fu, J.; Xie, Q. Efficacy of recombinant Newcastle disease virus expressing HA protein of H9N2 Avian influenza virus in respiratory and intestinal tract. Poult. Sci. 2022, 101, 102078. [Google Scholar] [CrossRef]
- Liu, D.; Shi, W.; Gao, G.F. Poultry carrying H9N2 act as incubators for novel human avian influenza viruses. Lancet. 2014, 383, 869. [Google Scholar] [CrossRef]
- Zhang, A.; Li, D.; Song, C.; Jing, H.; Li, H.; Mi, J.; Zhang, G.; Jin, S.; Ren, X.; Huangfu, H.; et al. Evaluation of different combination of pam2CSK4, poly (I:C) and imiquimod enhance immune responses to H9N2 avian influenza antigen in dendritic cells and duck. PLoS ONE 2022, 17, e0271746. [Google Scholar] [CrossRef] [PubMed]
- Teng, Q.; Shen, W.; Liu, Q.; Rong, G.; Chen, L.; Li, X.; Chen, H.; Yang, J.; Li, Z. Protective efficacy of an inactivated vaccine against H9N2 avian influenza virus in ducks. Virol. J. 2015, 12, 143. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Z.; Zhang, Q.; Sun, M.; Li, S.; Su, W.; Hu, X.; He, W.; Su, J. Efficacy assessment of an inactivated Tembusu virus vaccine candidate in ducks. Res Vet. Sci. 2017, 110, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.G.; Liu, M.; Liu, F.; Liu, D.F.; Zhang, Y.; Pan, W.Q.; Chen, H.; Wan, C.H.; Sun, E.C.; Li, H.T.; et al. Evaluation of several adjuvants in avian influenza vaccine to chickens and ducks. Virol. J. 2011, 8, 321. [Google Scholar] [CrossRef]
- Sabbaghi, A.; Miri, S.M.; Keshavarz, M.; Zargar, M.; Ghaemi, A. Inactivation methods for whole influenza vaccine production. Rev. Med. Virol. 2019, 29, e2074. [Google Scholar] [CrossRef] [PubMed]
- Magor, K.E. Immunoglobulin genetics and antibody responses to influenza in ducks. Dev. Comp. Immunol. 2011, 35, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, J.B.; Post, J.; Peeters, B.; Vervelde, L.; Rebel, J.M. Differential innate responses of chickens and ducks to low-pathogenic avian influenza. Avian Pathol. 2012, 41, 519–529. [Google Scholar] [CrossRef]
- Qi, Y.; Guo, W.; Liu, C.; Li, S.; Chen, X. Maternal transfer of antibodies specific for avian influenza viruses in captive whooper swans (Cygnus cygnus). Comp. Immunol. Microbiol. Infect. Dis. 2021, 76, 101644. [Google Scholar] [CrossRef]
- Rahimi, S.; Salehifar, E.; Ghorashi, S.; Grimes, J.; Mohammad, A.; Torshizi, K. The effect of egg- derived antibody on prevention of avian influenza subtype H9N2 in layer chicken. Int. J. Poult. Sci. 2007, 6, 207–210. [Google Scholar] [CrossRef][Green Version]
- Zhang, M.; Zhang, L.; Yang, J.; Zhao, D.; Han, K.; Huang, X.; Liu, Q.; Xiao, Y.; Gu, Y.; Li, Y. An IgY effectively prevents goslings from virulent GAstV infection. Vaccines 2022, 10, 2090. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, O.B.; Estevez, C.; Yu, Q.; Suarez, D.L. Passive antibody transfer in chickens to model maternal antibody after avian influenza vaccination. Vet. Immunol. Immunopathol. 2013, 152, 341–347. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| HI Antibody Titres (log2) | Virus Isolation/Total | Protection Rate (%) |
|---|---|---|
| 3 | 4/5 | 20 |
| 4 | 4/6 | 33 |
| 5 | 1/6 | 83 |
| 6 | 0/10 | 100 |
| 7 | 0/10 | 100 |
| 8 | 0/8 | 100 |
| 9 | 0/5 | 100 |
| 10 | 0/2 | 100 |
| Control | 10/10 | 0 |
| HI Antibody Titres (log2) | Virus Isolation/Total | Protection Rate (%) |
|---|---|---|
| 0 | 4/6 | 33 |
| 1 | 2/5 | 60 |
| 2 | 1/5 | 80 |
| 3 | 0/6 | 100 |
| 4 | 1/8 | 87.5 |
| 5 | 0/7 | 100 |
| 6 | 0/7 | 100 |
| 7 | 0/4 | 100 |
| 8 | 0/2 | 100 |
| Control | 8/10 | 20 |
| Group | Vaccination Volume (mL) | Dose (EID50) | Chickens | Ducks |
|---|---|---|---|---|
| The Ratio of HI TITRE ≥6 log2 (%) | The Ratio of HI Titre ≥4 log2 (%) | |||
| B1 B2 | 0.025 0.05 | 107.1 | 0/10 (0) | 0/10 (0) |
| 107.4 | 4/10 (40%) | 1/10 (10%) | ||
| B3 | 0.1 | 107.7 | 10/10 (100%) | 8/10 (80%) |
| B4 | 0.2 | 108.0 | 10/10 (100%) | 9/10 (90%) |
| B5 | 0.3 | 108.2 | 10/10 (100%) | 10/10 (100%) |
| Control | - | - | 0/10 (0) | 0/10 (0) |
| Group | Immunization | Field Strain | Chickens | Ducks | ||
|---|---|---|---|---|---|---|
| Mean HI Titre (log2) | Virus Isolation /Total (%) | Mean HI Titre (log2) | Virus Isolation /Total (%) | |||
| C1 C2 | Vaccine Vaccine | AH01 | 8.9 ± 0.74 | 0/10 (0) | 5.1 ± 0.94 | 0/10 (0) |
| HN03 | 9.0 ± 0.67 | 0/10 (0) | 4.9 ± 0.83 | 1/10 (10%) | ||
| C3 | - | AH01 | 0 | 10/10 (100%) | 0 | 10/10 (100%) |
| C4 | - | HN03 | 0 | 10/10 (100%) | 0 | 10/10 (100%) |
| Farm | Immunization | Chickens | Ducks | ||
|---|---|---|---|---|---|
| The Ratio of HI Titre ≥ 6 log2 (%) | Mean HI Titre (log2) | The Ratio of HI Titre ≥ 4 log2 (%) | Mean HI Titre (log2) | ||
| No. 1 | Vaccine | 100 | 7.9 ± 0.54 | 100 | 5.1 ± 1.01 |
| - | 0 | 0 | 0 | 0 | |
| No. 2 | Vaccine | 100 | 8.5 ± 0.86 | 90 | 4.3 ± 1.44 |
| - | 0 | 0 | 0 | 0 | |
| No. 3 | Vaccine | 100 | 7.8 ± 0.97 | 100 | 5.6 ± 1.05 |
| - | 0 | 0 | 0 | 0 | |
| Immunization | Serum | Yolk | ||
|---|---|---|---|---|
| Chickens | Vaccine | The ratio of HI titre ≥ 6 log2 (%) | 100 | 100 |
| Mean HI titre (log2) | 9.75 ± 0.71 | 9.2 ± 0.75 | ||
| - | The ratio of HI titre ≥ 6 log2 (%) | 0 | 0 | |
| Mean HI titre (log2) | 0 | 0 | ||
| Ducks | Vaccine | The ratio of HI titre ≥ 4 log2 (%) | 100 | 100 |
| Mean HI titre (log2) | 8.7 ± 1.02 | 8.4 ± 0.99 | ||
| - | The ratio of HI titre ≥ 4 log2 (%) | 0 | 0 | |
| Mean HI titre (log2) | 0 | 0 |
| Age (Day) | Immunization | Chicks | Ducklings | ||
|---|---|---|---|---|---|
| The Ratio of HI Titre ≥ 6 log2 (%) | Mean HI Titre (log2) | The Ratio of HI Titre ≥ 4 log2 (%) | Mean HI Titre (log2) | ||
| 1 | Vaccine | 100 | 8.6 ± 0.87 | 100 | 7.3 ± 0.82 |
| - | 0 | 0 | 0 | 0 | |
| 3 | Vaccine | 100 | 7.6 ± 0.82 | 100 | 5.9 ± 0.85 |
| - | 0 | 0 | 0 | 0 | |
| 6 | Vaccine | 90 | 6.9 ± 0.71 | 80 | 4.4 ± 0.97 |
| - | 0 | 0 | 0 | 0 | |
| 9 | Vaccine | 90 | 6.1 ± 0.60 | 50 | 3.1 ± 0.82 |
| - | 0 | 0 | 0 | 0 | |
| 12 | Vaccine | 40 | 5.3 ± 0.67 | 10 | 1.1 ± 0.74 |
| - | 0 | 0 | 0 | 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Zhao, D.; Zhang, J.; Huang, X.; Han, K.; Liu, Q.; Yang, J.; Zhang, L.; Li, Y. Development of an Inactivated Avian Influenza Virus Vaccine against Circulating H9N2 in Chickens and Ducks. Vaccines 2023, 11, 596. https://doi.org/10.3390/vaccines11030596
Liu Y, Zhao D, Zhang J, Huang X, Han K, Liu Q, Yang J, Zhang L, Li Y. Development of an Inactivated Avian Influenza Virus Vaccine against Circulating H9N2 in Chickens and Ducks. Vaccines. 2023; 11(3):596. https://doi.org/10.3390/vaccines11030596
Chicago/Turabian StyleLiu, Yuzhuo, Dongmin Zhao, Jingfeng Zhang, Xinmei Huang, Kaikai Han, Qingtao Liu, Jing Yang, Lijiao Zhang, and Yin Li. 2023. "Development of an Inactivated Avian Influenza Virus Vaccine against Circulating H9N2 in Chickens and Ducks" Vaccines 11, no. 3: 596. https://doi.org/10.3390/vaccines11030596
APA StyleLiu, Y., Zhao, D., Zhang, J., Huang, X., Han, K., Liu, Q., Yang, J., Zhang, L., & Li, Y. (2023). Development of an Inactivated Avian Influenza Virus Vaccine against Circulating H9N2 in Chickens and Ducks. Vaccines, 11(3), 596. https://doi.org/10.3390/vaccines11030596