
Immunotherapeutic Targeting of NG2/CSPG4 in Solid Organ Cancers

Abstract
:1. Brief Introduction to NG2/CSPG4 in Solid Organ Cancers
2. Initial Discovery of NG2/CSPG4 as a Precursor Marker in the CNS and Its Association with CNS Cancers
3. NG2/CSPG4 as a Marker of Precursor or Progenitor Cells outside the CNS and Its Association with Solid Organ Cancers
4. Selection of NG2/CSPG4 as a Target Antigen for the Treatment of Solid Organ Cancers
5. Conclusions and Perspective
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| bFGF | basic fibroblast growth factor |
| CNS | the central nervous system |
| CSCs | cancer stem cells |
| ECs | endothelial cells |
| ECM | extracellular matrix |
| HCC | hepatocellular carcinoma |
| HMW-MAA | human molecular weight melanoma-associated antigen |
| MCSP | melanoma chondroitin sulfate proteoglycan |
| MMPs | metalloproteinases |
| MSCs | mesenchymal stem cells |
| NG2/CSPG4 | neuro-glia antigen 2/chondroitin sulfate proteoglycan 4 |
| OPCs | oligodendrocyte precursor cells |
| PCs | pericytes |
| PDAC | pancreatic ductal adenocarcinoma |
| PDGFR-á | platelet-derived growth factor receptor á |
| PG | proteoglycan |
| SMAs | smooth muscle cells |
| SSCs | solid stem cells |
| SSEA-1 | stage-specific embryonic antigen |
References
- Polito, A.; Reynolds, R. NG2-expressing cells as oligodendrocyte progenitors in the normal and demyelinated adult central nervous system. J. Anat. 2005, 207, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, D.; Mellai, M.; Boldorini, R.; Bisogno, I.; Grifoni, S.; Corona, C.; Bertero, L.; Cassoni, P.; Casalone, C.; Annovazzi, L. The Significance of Chondroitin Sulfate Proteoglycan 4 (CSPG4) in Human Gliomas. Int. J. Mol. Sci. 2018, 19, 2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-González, R.; Bribián, A.; López-Mascaraque, L. Cell Fate Potential of NG2 Progenitors. Sci. Rep. 2020, 10, 9876. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; You, X.; Yang, W.; Jiang, S.; Lai, J.; Zhang, H.; Bai, L. Survival of endogenous hepatic stem/progenitor cells in liver tissues during liver cirrhosis. Life Sci. 2020, 241, 117121. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.L.; Sun, J.; Lai, J.J.; Jiang, Y.; Bai, L.H.; Zhang, L.D. Neuron-glial antigen 2 overexpression in hepatocellular carcinoma predicts poor prognosis. World J. Gastroenterol. 2015, 21, 6649–6659. [Google Scholar] [CrossRef]
- Kutluk Cenik, B.; Ostapoff, K.T.; Gerber, D.E.; Brekken, R.A. BIBF 1120 (nintedanib), a triple angiokinase inhibitor, induces hypoxia but not EMT and blocks progression of preclinical models of lung and pancreatic cancer. Mol. Cancer Ther. 2013, 12, 992–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.D.; Stallcup, W.; Lefkove, B.; Govindarajan, B.; Au, K.S.; Northrup, H.; Lang, D.; Fisher, D.E.; Patel, A.; Amin, M.B.; et al. Expression of the neural stem cell markers NG2 and L1 in human angiomyolipoma: Are angiomyolipomas neoplasms of stem cells? Mol. Med. 2007, 13, 160–165. [Google Scholar] [CrossRef]
- Stallcup, W.B. The NG2 antigen, a putative lineage marker: Immunofluorescent localization in primary cultures of rat brain. Dev. Biol. 1981, 83, 154–165. [Google Scholar] [CrossRef]
- Nishiyama, A.; Lin, X.H.; Stallcup, W.B. Generation of truncated forms of the NG2 proteoglycan by cell surface proteolysis. Mol. Biol. Cell 1995, 6, 1819–1832. [Google Scholar] [CrossRef] [Green Version]
- Goretzki, L.; Burg, M.A.; Grako, K.A.; Stallcup, W.B. High-affinity binding of basic fibroblast growth factor and platelet-derived growth factor-AA to the core protein of the NG2 proteoglycan. J. Biol. Chem. 1999, 274, 16831–16837. [Google Scholar] [CrossRef] [Green Version]
- Melrose, J.; Hayes, A.J.; Bix, G. The CNS/PNS Extracellular Matrix Provides Instructive Guidance Cues to Neural Cells and Neuroregulatory Proteins in Neural Development and Repair. Int. J. Mol. Sci. 2021, 22, 5583. [Google Scholar] [CrossRef] [PubMed]
- Alex, L.; Tuleta, I.; Harikrishnan, V.; Frangogiannis, N.G. Validation of Specific and Reliable Genetic Tools to Identify, Label, and Target Cardiac Pericytes in Mice. J. Am. Heart Assoc. 2022, 11, e023171. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.J.; You, W.K.; Bonaldo, P.; Seyfried, T.N.; Pasquale, E.B.; Stallcup, W.B. Pericyte deficiencies lead to aberrant tumor vascularizaton in the brain of the NG2 null mouse. Dev. Biol. 2010, 344, 1035–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelyshev, Y.A.; Kabdesh, I.M.; Mukhamedshina, Y.O. Extracellular Matrix in Neural Plasticity and Regeneration. Cell Mol. Neurobiol. 2022, 42, 647–664. [Google Scholar]
- Bluemel, C.; Hausmann, S.; Fluhr, P.; Sriskandarajah, M.; Stallcup, W.B.; Baeuerle, P.A.; Kufer, P. Epitope distance to the target cell membrane and antigen size determine the potency of T cell-mediated lysis by BiTE antibodies specific for a large melanoma surface antigen. Cancer Immunol. Immunother. 2010, 59, 1197–1209. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Bosse, F.; D’Urso, D.; Muller, H.; Sereda, M.W.; Nave, K.; Niehaus, A.; Kempf, T.; Schnölzer, M.; Trotter, J. The AN2 protein is a novel marker for the Schwann cell lineage expressed by immature and nonmyelinating Schwann cells. J. Neurosci. 2001, 21, 920–933. [Google Scholar] [CrossRef] [Green Version]
- Dahl, J.A.; Jung, I.; Aanes, H.; Greggains, G.D.; Manaf, A.; Lerdrup, M.; Li, G.; Kuan, S.; Li, B.; Lee, A.Y.; et al. Broad histone H3K4me3 domains in mouse oocytes modulate maternal-to-zygotic transition. Nature 2016, 537, 548–552. [Google Scholar] [CrossRef]
- Trost, A.; Bruckner, D.; Rivera, F.J.; Reitsamer, H.A. Pericytes in the Retina. Adv. Exp. Med. Biol. 2019, 1122, 1–26. [Google Scholar] [PubMed]
- Reynolds, R.; Dawson, M.; Papadopoulos, D.; Polito, A.; Di Bello, I.C.; Pham-Dinh, D.; Levine, J. The response of NG2-expressing oligodendrocyte progenitors to demyelination in MOG-EAE and, M.S. J. Neurocytol. 2002, 31, 523–536. [Google Scholar] [CrossRef]
- Bai, L.; Hecker, J.; Kerstetter, A.; Miller, R.H. Myelin repair and functional recovery mediated by neural cell transplantation in a mouse model of multiple sclerosis. Neurosci. Bull. 2013, 29, 239–250. [Google Scholar] [CrossRef]
- Busch, S.A.; Horn, K.P.; Cuascut, F.X.; Hawthorne, A.L.; Bai, L.; Miller, R.H.; Silver, J. Adult NG2+ cells are permissive to neurite outgrowth and stabilize sensory axons during macrophage-induced axonal dieback after spinal cord injury. J. Neurosci. 2010, 30, 255–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, X.; Bie, P.; Miller, R.H.; Bai, L. Adult NG2-Expressing Cells in Multiple Organs: A Novel Progenitor in Regenerative Medicine. J. Genet. Syndr. Gene Ther. 2013, S3, 1. [Google Scholar]
- Zhang, H.; Siegel, C.T.; Shuai, L.; Lai, J.; Zeng, L.; Zhang, Y.; Lai, X.; Bie, P.; Bai, L. Repair of liver mediated by adult mouse liver neuro-glia antigen 2-positive progenitor cell transplantation in a mouse model of cirrhosis. Sci. Rep. 2016, 6, 21783. [Google Scholar] [CrossRef]
- Yuan, H.; Fang, C.; Deng, Y.; Huang, J.; Niu, R.; Chen, J.; Chen, T.; Zhu, Z.; Chen, L.; Xiong, L.; et al. A2B5-positive oligodendrocyte precursor cell transplantation improves neurological deficits in rats following spinal cord contusion associated with changes in expression of factors involved in the Notch signaling pathway. Neurochir. 2022, 68, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Keirstead, H.S.; Levine, J.M.; Blakemore, W.F. Response of the oligodendrocyte progenitor cell population (defined by NG2 labelling) to demyelination of the adult spinal cord. Glia 1998, 22, 161–170. [Google Scholar] [CrossRef]
- McTigue, D.M.; Wei, P.; Stokes, B.T. Proliferation of NG2-positive cells and altered oligodendrocyte numbers in the contused rat spinal cord. J. Neurosci. 2001, 21, 3392–3400. [Google Scholar] [CrossRef] [PubMed]
- Gomes, N.A.; do Valle, I.B.; Gleber-Netto, F.O.; Silva, T.A.; Oliveira, H.M.C.; de Oliveira, R.F.; Ferreira, L.D.A.Q.; Castilho, L.S.; Reis, P.H.R.G.; Prazeres, P.H.D.M.; et al. Nestin and NG2 transgenes reveal two populations of perivascular cells stimulated by photobiomodulation. J. Cell Physiol. 2022, 237, 2198–2210. [Google Scholar] [CrossRef]
- Wilms, C.; Lepka, K.; Häberlein, F.; Edwards, S.; Felsberg, J.; Pudelko, L.; Lindenberg, T.T.; Poschmann, G.; Qin, N.; Volbracht, K.; et al. Glutaredoxin 2 promotes SP-1-dependent CSPG4 transcription and migration of wound healing NG2 glia and glioma cells: Enzymatic Taoism. Redox. Biol. 2022, 49, 102221. [Google Scholar] [CrossRef] [PubMed]
- Yi, B.; Li, H.; Cai, H.; Lou, X.; Yu, M.; Li, Z. LOXL1-AS1 communicating with TIAR modulates vasculogenic mimicry in glioma via regulation of the miR-374b-5p/MMP14 axis. J. Cell Mol. Med. 2022, 26, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.S.; Dejesus, J.; Short, A.R.; Otero, J.J.; Sarkar, A.; Winter, J.O. Glioblastoma behaviors in three-dimensional collagen-hyaluronan composite hydrogels. ACS Appl. Mater. Interfaces 2013, 5, 9276–9284. [Google Scholar] [CrossRef] [PubMed]
- Le Joncour, V.; Guichet, P.O.; Dembélé, K.P.; Mutel, A.; Campisi, D.; Perzo, N.; Desrues, L.; Modzelewski, R.; Couraud, P.-O.; Honnorat, J.; et al. Targeting the Urotensin II/UT G Protein-Coupled Receptor to Counteract Angiogenesis and Mesenchymal Hypoxia/Necrosis in Glioblastoma. Front. Cell Dev. Biol. 2021, 9, 652544. [Google Scholar] [CrossRef]
- Chekenya, M.; Krakstad, C.; Svendsen, A.; Netland, I.A.; Staalesen, V.; Tysnes, B.B.; Selheim, F.; Wang, J.; Sakariassen, P.Ø.; Sandal, T.; et al. The progenitor cell marker NG2/MPG promotes chemoresistance by activation of integrin-dependent PI3K/Akt signaling. Oncogene 2008, 27, 5182–5194. [Google Scholar] [CrossRef] [Green Version]
- Long, P.M.; Tighe, S.W.; Driscoll, H.E.; Moffett, J.R.; Namboodiri, A.M.; Viapiano, M.S.; Lawler, S.E.; Jaworski, D.M. Acetate supplementation induces growth arrest of NG2/PDGFRα-positive oligodendroglioma-derived tumor-initiating cells. PLoS ONE 2013, 8, e80714. [Google Scholar]
- Svendsen, A.; Verhoeff, J.J.; Immervoll, H.; Brøgger, J.C.; Kmiecik, J.; Poli, A.; Netland, I.A.; Prestegarden, L.; Planagumà, J.; Torsvik, A.; et al. Expression of the progenitor marker NG2/CSPG4 predicts poor survival and resistance to ionising radiation in glioblastoma. Acta Neuropathol. 2011, 122, 495–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, A.H.; Holland, E.C. Platelet-derived growth factor (PDGF) and glial tumorigenesis. Cancer Lett. 2006, 232, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, S.H.; Nadesan, P.; Puviindran, V.; Stallcup, W.B.; Kirsch, D.G.; Alman, B.A. Effects of chondroitin sulfate proteoglycan 4 (NG2/CSPG4) on soft-tissue sarcoma growth depend on tumor developmental stage. J. Biol. Chem. 2018, 293, 2466–2475. [Google Scholar] [CrossRef] [Green Version]
- Weng, X.; Li, J.; Guan, Q.; Zhao, H.; Wang, Z.; Gleave, M.E.; Nguan, C.Y.C.; Du, C. The functions of clusterin in renal mesenchymal stromal cells: Promotion of cell growth and regulation of macrophage activation. Exp. Cell Res. 2022, 413, 113081. [Google Scholar] [CrossRef] [PubMed]
- Mi, G.; Gao, Y.; Liu, S.; Ye, E.; Li, Y.; Jin, X.; Yang, H.; Yang, Z. Cyclin-dependent kinase inhibitor flavopiridol promotes remyelination in a cuprizone induced demyelination model. Cell Cycle 2016, 15, 2780–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.I.; Correa, D. The MSC: An injury drugstore. Cell Stem Cell 2011, 9, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, E.; Corona, C.; Eleftheriou, C.G.; Bianchimano, P.; Sagdullaev, B.T. Retina-specific targeting of pericytes reveals structural diversity and enables control of capillary blood flow. J. Comp. Neurol. 2021, 529, 1121–1134. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Bai, X.; Meyer, E.; Scheller, A. Acute brain injuries trigger microglia as an additional source of the proteoglycan NG2. Acta Neuropathol. Commun. 2020, 8, 146. [Google Scholar] [CrossRef] [PubMed]
- Teichert, M.; Milde, L.; Holm, A.; Stanicek, L.; Gengenbacher, N.; Savant, S.; Ruckdeschel, T.; Hasanov, Z.; Srivastava, K.; Hu, J.; et al. Pericyte-expressed Tie2 controls angiogenesis and vessel maturation. Nat. Commun. 2017, 8, 16106. [Google Scholar] [CrossRef]
- Smyth, L.C.D.; Rustenhoven, J.; Scotter, E.L.; Schweder, P.; Faull, R.L.M.; Park, T.I.H.; Dragunow, M. Markers for human brain pericytes and smooth muscle cells. J. Chem. Neuroanat. 2018, 92, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Mykuliak, A.; Yrjänäinen, A.; Mäki, A.J.; Gebraad, A.; Lampela, E.; Kääriäinen, M.; Pakarinen, T.-K.; Kallio, P.; Miettinen, S.; Vuorenpää, H.; et al. Vasculogenic Potency of Bone Marrow- and Adipose Tissue-Derived Mesenchymal Stem/Stromal Cells Results in Differing Vascular Network Phenotypes in a Microfluidic Chip. Front. Bioeng. Biotechnol. 2022, 10, 764237. [Google Scholar] [CrossRef]
- Tao, Y.K.; Zeng, H.; Zhang, G.Q.; Chen, S.T.; Xie, X.J.; He, X.; Wang, S.; Wen, H.; Chen, J.-X. Notch3 deficiency impairs coronary microvascular maturation and reduces cardiac recovery after myocardial ischemia. Int. J. Cardiol. 2017, 236, 413–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenoy, P.S.; Bose, B. Hepatic perivascular mesenchymal stem cells with myogenic properties. J. Tissue Eng. Regen. Med. 2018, 12, e1297–e1310. [Google Scholar] [CrossRef]
- Carlotti, F.; Zaldumbide, A.; Loomans, C.J.; van Rossenberg, E.; Engelse, M.; de Koning, E.J.; Hoeben, R.C. Isolated human islets contain a distinct population of mesenchymal stem cells. Islets 2010, 2, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Barilani, M.; Banfi, F.; Sironi, S.; Ragni, E.; Guillaumin, S.; Polveraccio, F.; Rosso, L.; Moro, M.; Astori, G.; Pozzobon, M.; et al. Low-affinity Nerve Growth Factor Receptor (CD271) Heterogeneous Expression in Adult and Fetal Mesenchymal Stromal Cells. Sci. Rep. 2018, 8, 9321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butt, A.M.; Pugh, M.; Hubbard, P.; James, G. Functions of optic nerve glia: Axoglial signalling in physiology and pathology. Eye 2004, 18, 1110–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanishi, H.; Fujiwara, S.; Soma, T. Perivascular localization of dermal stem cells in human scalp. Exp. Dermatol. 2012, 21, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Uusitalo-Kylmälä, L.; Santo Mendes, A.C.; Polari, L.; Joensuu, K.; Heino, T.J. An In Vitro Co-Culture Model of Bone Marrow Mesenchymal Stromal Cells and Peripheral Blood Mononuclear Cells Promotes the Differentiation of Myeloid Angiogenic Cells and Pericyte-Like Cells. Stem Cells Dev. 2021, 30, 309–324. [Google Scholar] [CrossRef]
- Crisan, M.; Corselli, M.; Chen, W.C.; Péault, B. Perivascular cells for regenerative medicine. J. Cell Mol. Med. 2012, 16, 2851–2860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, J.; Jiang, S.; Shuai, L.; Zhang, Y.; Xia, R.; Chen, Q.; Bai, L. Comparison of the biological and functional characteristics of mesenchymal stem cells from intrahepatic and identical bone marrow. Stem Cell Res. 2021, 55, 102477. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Siegel, C.T.; Li, J.; Lai, J.; Shuai, L.; Lai, X.; Zhang, Y.; Jiang, Y.; Bie, P.; Bai, L. Functional liver tissue engineering by an adult mouse liver-derived neuro-glia antigen 2-expressing stem/progenitor population. J. Tissue Eng. Regen. Med. 2018, 12, e190–e202. [Google Scholar] [CrossRef] [PubMed]
- Burg, M.A.; Pasqualini, R.; Arap, W.; Ruoslahti, E.; Stallcup, W.B. NG2 proteoglycan-binding peptides target tumor neovasculature. Cancer Res. 1999, 59, 2869–2874. [Google Scholar]
- Caporarello, N.; D’Angeli, F.; Cambria, M.T.; Candido, S.; Giallongo, C.; Salmeri, M.; Lombardo, C.; Longo, A.; Giurdanella, G.; Anfuso, C.D.; et al. Pericytes in Microvessels: From “Mural” Function to Brain and Retina Regeneration. Int. J. Mol. Sci. 2019, 20, 6351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, E.V.; Figueroa, R.J.; Bussmann, J.; Lawson, N.D.; Amigo, J.D.; Siekmann, A.F. Regenerating vascular mural cells in zebrafish fin blood vessels are not derived from pre-existing mural cells and differentially require Pdgfrb signalling for their development. Development 2022, 149, dev199640. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, H.; Mundel, T.M.; Kieran, M.W.; Kalluri, R. Identification of fibroblast heterogeneity in the tumor microenvironment. Cancer Biol. Ther. 2006, 5, 1640–1646. [Google Scholar] [CrossRef] [Green Version]
- Bagley, R.G.; Rouleau, C.; Morgenbesser, S.D.; Weber, W.; Cook, B.P.; Shankara, S.; Madden, S.L.; Teicher, B.A. Pericytes from human non-small cell lung carcinomas: An attractive target for anti-angiogenic therapy. Microvasc. Res. 2006, 71, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Virgintino, D.; Girolamo, F.; Errede, M.; Capobianco, C.; Robertson, D.; Stallcup, W.B.; Perris, R.; Roncali, L. An intimate interplay between precocious, migrating pericytes and endothelial cells governs human fetal brain angiogenesis. Angiogenesis 2007, 10, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Nasarre, P.; Thomas, M.; Kruse, K.; Helfrich, I.; Wolter, V.; Deppermann, C.; Schadendorf, D.; Thurston, G.; Fiedler, U.; Augustin, H.G. Host-derived angiopoietin-2 affects early stages of tumor development and vessel maturation but is dispensable for later stages of tumor growth. Cancer Res. 2009, 69, 1324–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Svendsen, A.; Kmiecik, J.; Immervoll, H.; Skaftnesmo, K.O.; Planaguma, J.; Reed, R.K.; Bjerkvig, R.; Miletic, H.; Enger, P.Ø.; et al. Targeting the NG2/CSPG4 Proteoglycan Retards Tumour Growth and Angiogenesis in Preclinical Models of GBM and Melanoma. PLoS ONE 2011, 6, e23062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, N.; Hahn, C.N.; Poh, A.; Dong, C.; Wilhelm, D.; Olsson, J.; Muscat, G.E.O.; Parsons, P.; Gamble, J.R.; Koopman, P. Effect of disrupted SOX18 transcription factor function on tumor growth, vascularization, and endothelial development. J. Natl. Cancer Inst. 2006, 98, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Benassi, M.S.; Pazzaglia, L.; Chiechi, A.; Alberghini, M.; Conti, A.; Cattaruzza, S.; Wassermann, B.; Picci, P.; Perris, R. NG2 expression predicts the metastasis formation in soft-tissue sarcoma patients. J. Orthop. Res. 2009, 27, 135–140. [Google Scholar] [CrossRef]
- Zimmerlin, L.; Park, T.S.; Bhutto, I.; Lutty, G.; Zambidis, E.T. Generation of Pericytic-Vascular Progenitors from Tankyrase/PARP-Inhibitor-Regulated Naïve (TIRN) Human Pluripotent Stem Cells. Methods Mol. Biol. 2022, 2416, 133–156. [Google Scholar] [PubMed]
- Cheng, L.; Huang, Z.; Zhou, W.; Wu, Q.; Donnola, S.; Liu, J.K.; Fang, X.; Sloan, A.E.; Mao, Y.; Lathia, J.D.; et al. Glioblastoma stem cells generate vascular pericytes to support vessel function and tumor growth. Cell 2013, 153, 139–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordaan, S.; Chetty, S.; Mungra, N.; Koopmans, I.; van Bommel, P.E.; Helfrich, W.; Barth, S. CSPG4: A Target for Selective Delivery of Human Cytolytic Fusion Proteins and TRAIL. Biomedicines 2017, 5, 37. [Google Scholar] [CrossRef]
- Matuszewska, K.; Ten Kortenaar, S.; Pereira, M.; Santry, L.A.; Petrik, D.; Lo, K.M.; Bridle, B.W.; Wootton, S.K.; Lawler, J.; Petrik, J. Addition of an Fc-IgG induces receptor clustering and increases the in vitro efficacy and in vivo anti-tumor properties of the thrombospondin-1 type I repeats (3TSR) in a mouse model of advanced stage ovarian cancer. Gynecol. Oncol. 2022, 164, 154–169. [Google Scholar] [CrossRef]
- Kujawski, M.; Li, L.; Li, H.; Yazaki, P.J.; Swiderski, P.; Shively, J.E. T-cell surface generation of dual bivalent, bispecific T-cell engaging, RNA duplex cross-linked antibodies (dbBiTERs) for re-directed tumor cell lysis. Biotechnol. J. 2022, 17, e2100389. [Google Scholar] [CrossRef]
- Deisting, W.; Raum, T.; Kufer, P.; Baeuerle, P.A.; Münz, M. Impact of Diverse Immune Evasion Mechanisms of Cancer Cells on T Cells Engaged by EpCAM/CD3-Bispecific Antibody Construct AMG 110. PLoS ONE 2015, 10, e0141669. [Google Scholar] [CrossRef] [PubMed]
- Chocarro, L.; Arasanz, H.; Fernández-Rubio, L.; Blanco, E.; Echaide, M.; Bocanegra, A.; Teijeira, L.; Garnica, M.; Morilla, I.; Martínez-Aguillo, M.; et al. CAR-T Cells for the Treatment of Lung Cancer. Life 2022, 12, 561. [Google Scholar] [CrossRef]
- Vahidian, F.; Khosroshahi, L.M.; Akbarzadeh, M.; Jahanban-Esfahlan, A.; Baghbanzadeh, A.; Ali-Hassanzadeh, M.; Safarzadeh, E. The tricks for fighting against cancer using CAR NK cells: A review. Mol. Cell Probes 2022, 63, 101817. [Google Scholar] [CrossRef]
- Ebrahimiyan, H.; Tamimi, A.; Shokoohian, B.; Minaei, N.; Memarnejadian, A.; Hossein-Khannazer, N.; Hassan, M.; Vosough, M. Novel insights in CAR-NK cells beyond CAR-T cell technology; promising advantages. Int. Immunopharmacol. 2022, 106, 108587. [Google Scholar] [CrossRef]
- Rodriguez, A.; Brown, C.; Badie, B. Chimeric antigen receptor T-cell therapy for glioblastoma. Transl. Res. 2017, 187, 93–102. [Google Scholar] [CrossRef]
- Wang, C.K.; Nelepcu, I.; Hui, D.; Oo, H.Z.; Truong, S.; Zhao, S.; Tahiry, Z.; Esfandnia, S.; Ghaidi, F.; Adomat, H.; et al. Internalization and trafficking of CSPG-bound recombinant VAR2CSA lectins in cancer cells. Sci. Rep. 2022, 12, 3075. [Google Scholar] [CrossRef]
- Basu, A.; Patel, N.G.; Nicholson, E.D.; Weiss, R.J. Spatiotemporal diversity and regulation of glycosaminoglycans in cell homeostasis and human disease. Am. J. Physiol. Cell Physiol. 2022, 322, C849–C864. [Google Scholar] [CrossRef] [PubMed]
- Makaremi, S.; Asadzadeh, Z.; Hemmat, N.; Baghbanzadeh, A.; Sgambato, A.; Ghorbaninezhad, F.; Safapour, H.; Argentiero, A.; Brunetti, O.; Bernardini, R.; et al. Immune Checkpoint Inhibitors in Colorectal Cancer: Challenges and Future Prospects. Biomedicines 2021, 9, 1075. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, V.; Shemesh, C.S.; Rotte, A. Pharmacology-based ranking of anti-cancer drugs to guide clinical development of cancer immunotherapy combinations. J. Exp. Clin. Cancer Res. 2021, 40, 311. [Google Scholar] [PubMed]
- Pan, K.; Farrukh, H.; Chittepu, V.C.; Xu, H.; Pan, C.; Zhu, Z. CAR race to cancer immunotherapy: From CAR T, CAR NK to CAR macrophage therapy. J. Exp. Clin. Cancer Res. 2022, 41, 119. [Google Scholar]
- Yu, X.; Li, W.; Young, K.H.; Li, Y. Posttranslational Modifications in PD-L1 Turnover and Function: From Cradle to Grave. Biomedicines 2021, 9, 1702. [Google Scholar] [CrossRef]
- Keleg, S.; Titov, A.; Heller, A.; Giese, T.; Tjaden, C.; Ahmad, S.S.; Gaida, M.M.; Bauer, A.S.; Werner, J.; Giese, N.A. Chondroitin sulfate proteoglycan CSPG4 as a novel hypoxia-sensitive marker in pancreatic tumors. PLoS ONE 2014, 9, e100178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warta, R.; Herold-Mende, C.; Chaisaingmongkol, J.; Popanda, O.; Mock, A.; Mogler, C.; Osswald, F.; Herpel, E.; Küstner, S.; Eckstein, V.; et al. Reduced promoter methylation and increased expression of CSPG4 negatively influences survival of HNSCC patients. Int. J. Cancer 2014, 135, 2727–2734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.Y.; Lee, Y.K.; Koo, J.S. Expression of cancer-associated fibroblast-related proteins in adipose stroma of breast cancer. Tumour Biol. 2015, 36, 8685–8695. [Google Scholar] [CrossRef]
- Hu, Z.Y.; Zheng, C.; Yang, J.; Ding, S.; Tian, C.; Xie, N.; Xue, L.; Wu, M.; Fu, S.; Rao, Z.; et al. Co-Expression and Combined Prognostic Value of CSPG4 and PDL1 in TP53-Aberrant Triple-Negative Breast Cancer. Front Oncol. 2022, 12, 804466. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Lee, Y.K.; Koo, J.S. Expression of CAF-Related Proteins Is Associated with Histologic Grade of Breast Phyllodes Tumor. Dis. Markers. 2016, 2016, 4218989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenfeld, A.J.; Wang, X.; Wang, Y.; Hornicek, F.J.; Nielsen, G.P.; Duan, Z.; Ferrone, S.; Schwab, J.H. CSPG4 as a prognostic biomarker in chordoma. Spine J. 2016, 16, 722–727. [Google Scholar] [CrossRef] [PubMed]
- Al-Mayhani, M.; Grenfell, R.; Narita, M.; Piccirillo, S.; Kenney-Herbert, E.; Fawcett, J.W.; Collins, V.P.; Ichimura, K.; Watts, C. NG2 expression in glioblastoma identifies an actively proliferating population with an aggressive molecular signature. Neuro-oncology 2011, 13, 830–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, H.; Sun, T.; Chen, J.; Guo, L.; Shen, H.; Du, Z.; Zhou, Y. Biological characteristics of a new human glioma cell line transformed into A2B5(+) stem cells. Mol. Cancer. 2015, 14, 75. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Liu, Y.; Xie, X.; Zhu, T.; Soules, M.; DiMeco, F.; Vescovi, A.L.; Fan, X.; Lubman, D.M. Identification of cell surface glycoprotein markers for glioblastoma-derived stem-like cells using a lectin microarray and LC-MS/MS approach. J. Proteome Res. 2010, 9, 2565–2572. [Google Scholar] [CrossRef] [Green Version]
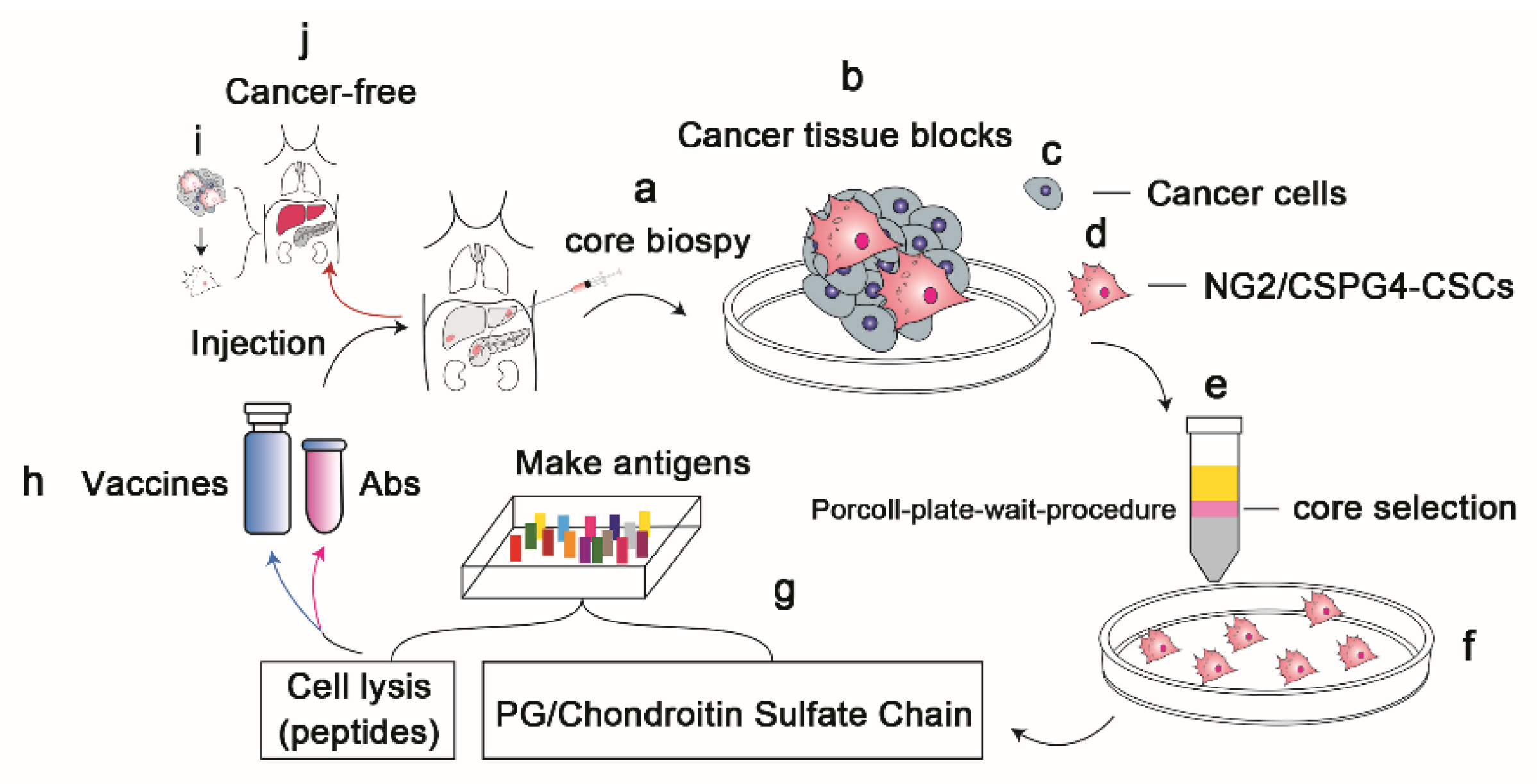
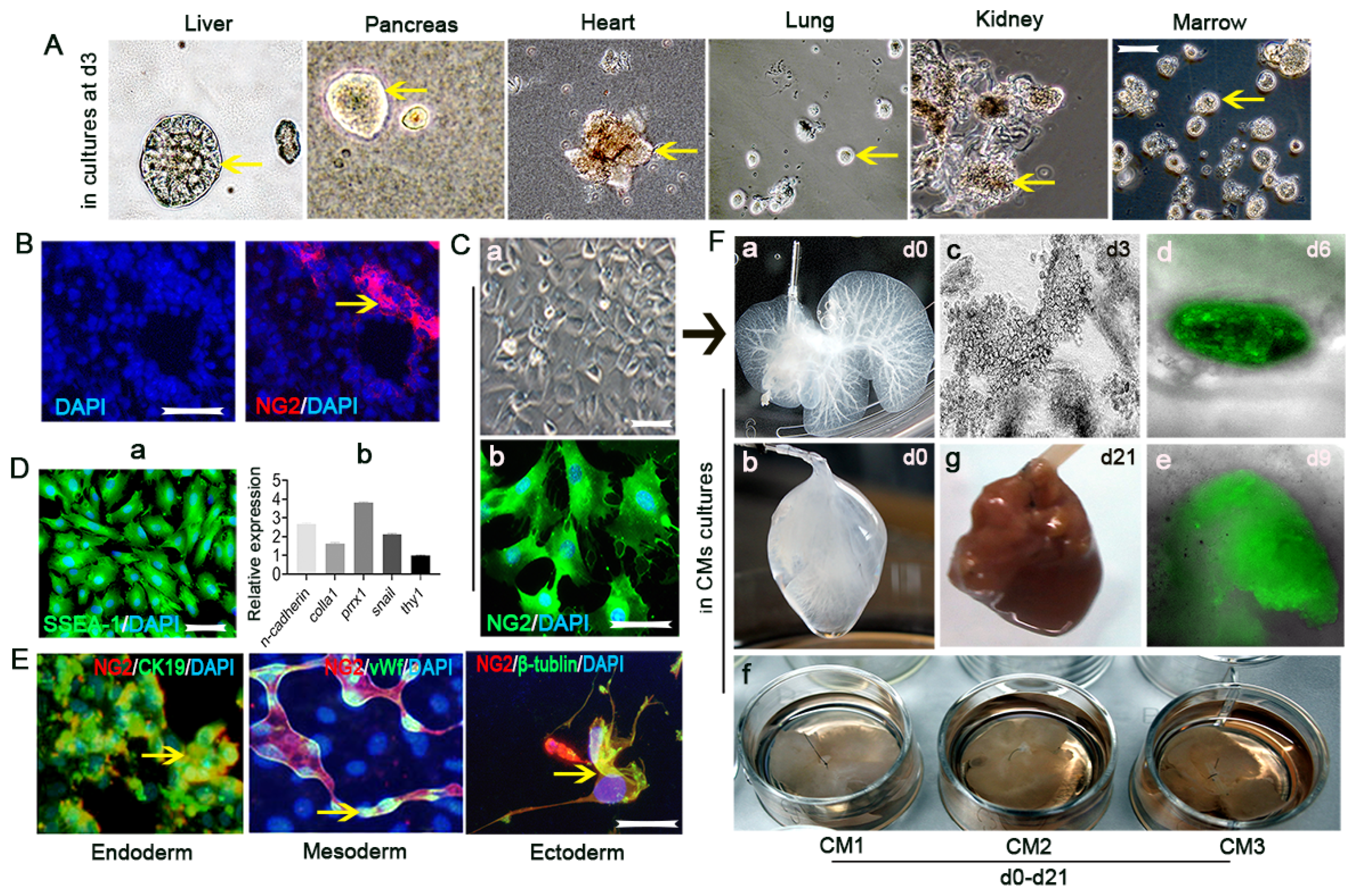
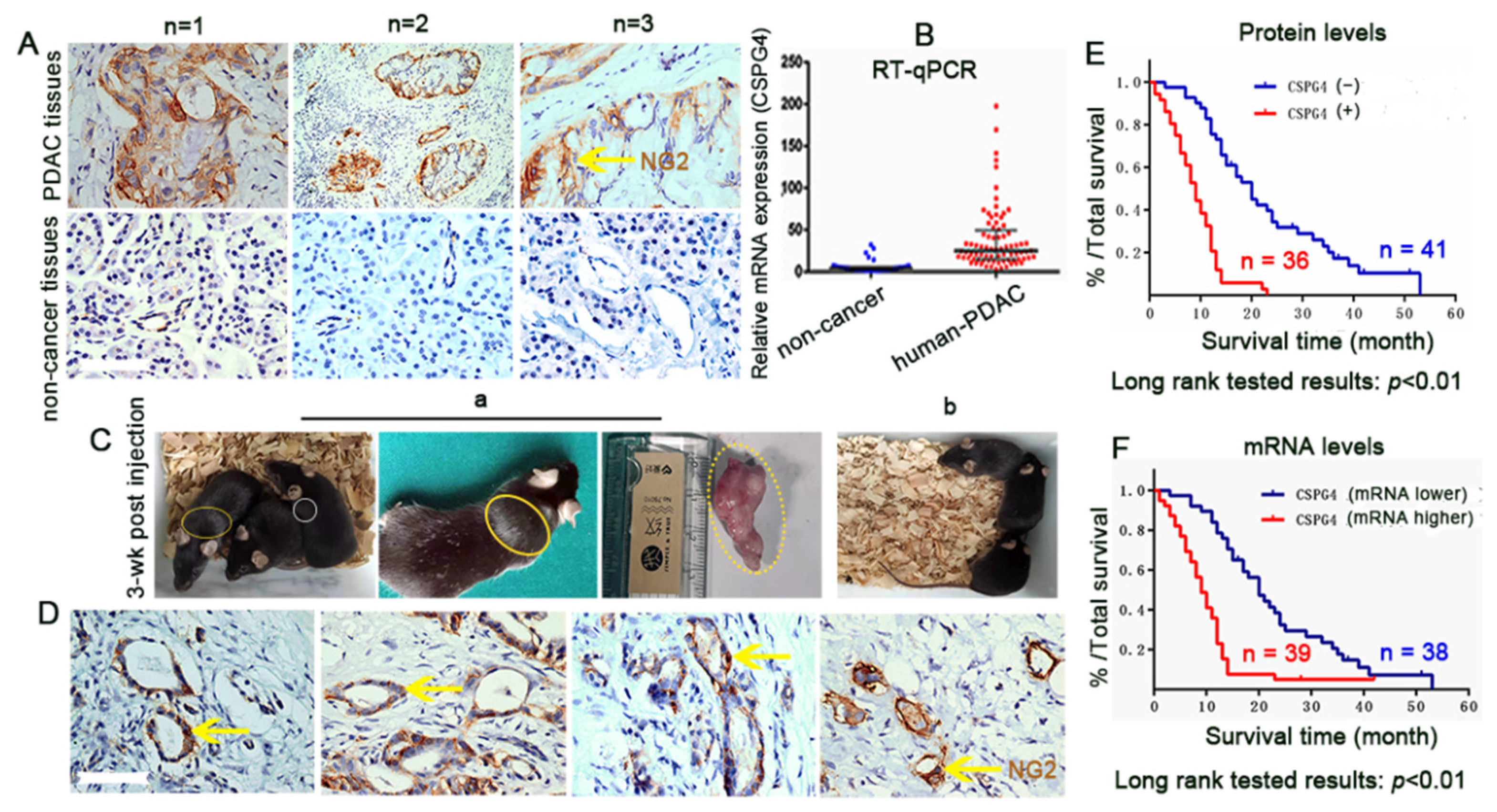

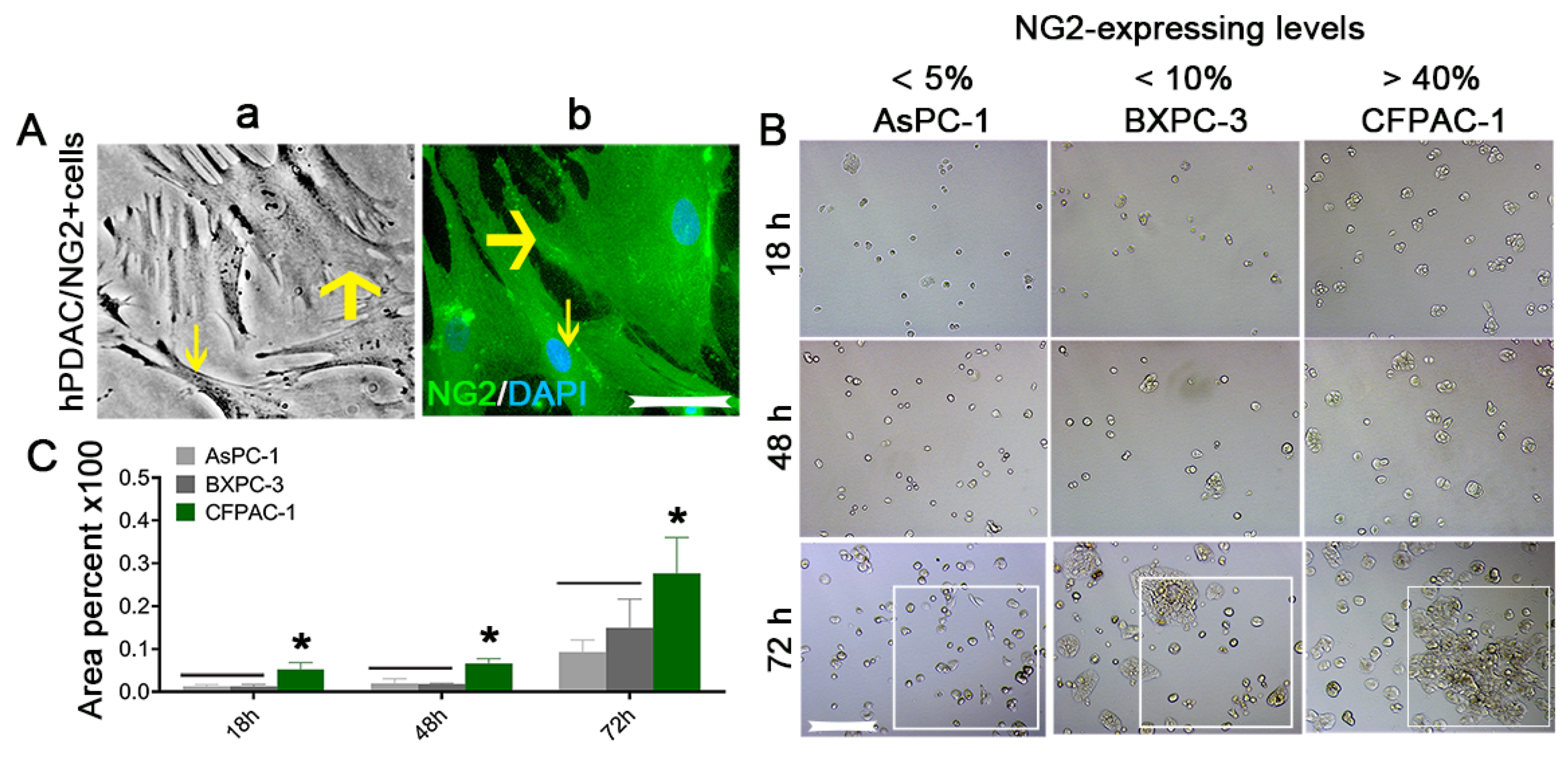

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Types | Patients | NG2/CSPG4 | High NG2/CSPG4 | References | |
|---|---|---|---|---|---|
| (Case) | mRNA | Protein | Outcomes | ||
| Pancreas | 256 | qPCR | IHC * | poor | [82] |
| Breast | 240 | qPCR | IHC | poor | [83,84,85] |
| Breast Phyllodes | 194 | IHC | poor | [86] | |
| Nead neck squcemeus carcema | unknow | qPCR | IHC | poor | [83] |
| Chordoma | 86 | IHC | poor | [87] | |
| Glioblastema (GBM) | unknow | qPCR | IHC | poor | [88,89,90] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wu, Z.; Hu, D.; Yan, M.; Sun, J.; Lai, J.; Bai, L. Immunotherapeutic Targeting of NG2/CSPG4 in Solid Organ Cancers. Vaccines 2022, 10, 1023. https://doi.org/10.3390/vaccines10071023
Zhang H, Wu Z, Hu D, Yan M, Sun J, Lai J, Bai L. Immunotherapeutic Targeting of NG2/CSPG4 in Solid Organ Cancers. Vaccines. 2022; 10(7):1023. https://doi.org/10.3390/vaccines10071023
Chicago/Turabian StyleZhang, Hongyu, Zhenyu Wu, Deyu Hu, Min Yan, Jing Sun, Jiejuan Lai, and Lianhua Bai. 2022. "Immunotherapeutic Targeting of NG2/CSPG4 in Solid Organ Cancers" Vaccines 10, no. 7: 1023. https://doi.org/10.3390/vaccines10071023
APA StyleZhang, H., Wu, Z., Hu, D., Yan, M., Sun, J., Lai, J., & Bai, L. (2022). Immunotherapeutic Targeting of NG2/CSPG4 in Solid Organ Cancers. Vaccines, 10(7), 1023. https://doi.org/10.3390/vaccines10071023