

NMDA Receptors Regulate Oxidative Damage in Keratinocytes during Complex Regional Pain Syndrome in HaCaT Cells and Male Rats


Abstract

1. Introduction
2. Materials and Methods
2.1. Cell Culture and Establishment of an OGD/R HaCaT Cell Model
2.2. In Vitro Experimental Design and Grouping
2.3. Animals and the Rat Model of CPIP
2.4. Animal Study Design
2.5. Behavioral Tests
2.6. Cell Viability Assay and Tunnel Staining
2.7. Immunofluorescence
2.8. Measurement of Mitochondrial Membrane Potential
2.9. Transmission Electron Microscopy
2.10. Real-Time Quantitative Polymerase Chain Reaction
2.11. RNA-Seq Analysis
2.12. Statistical Analysis
3. Results
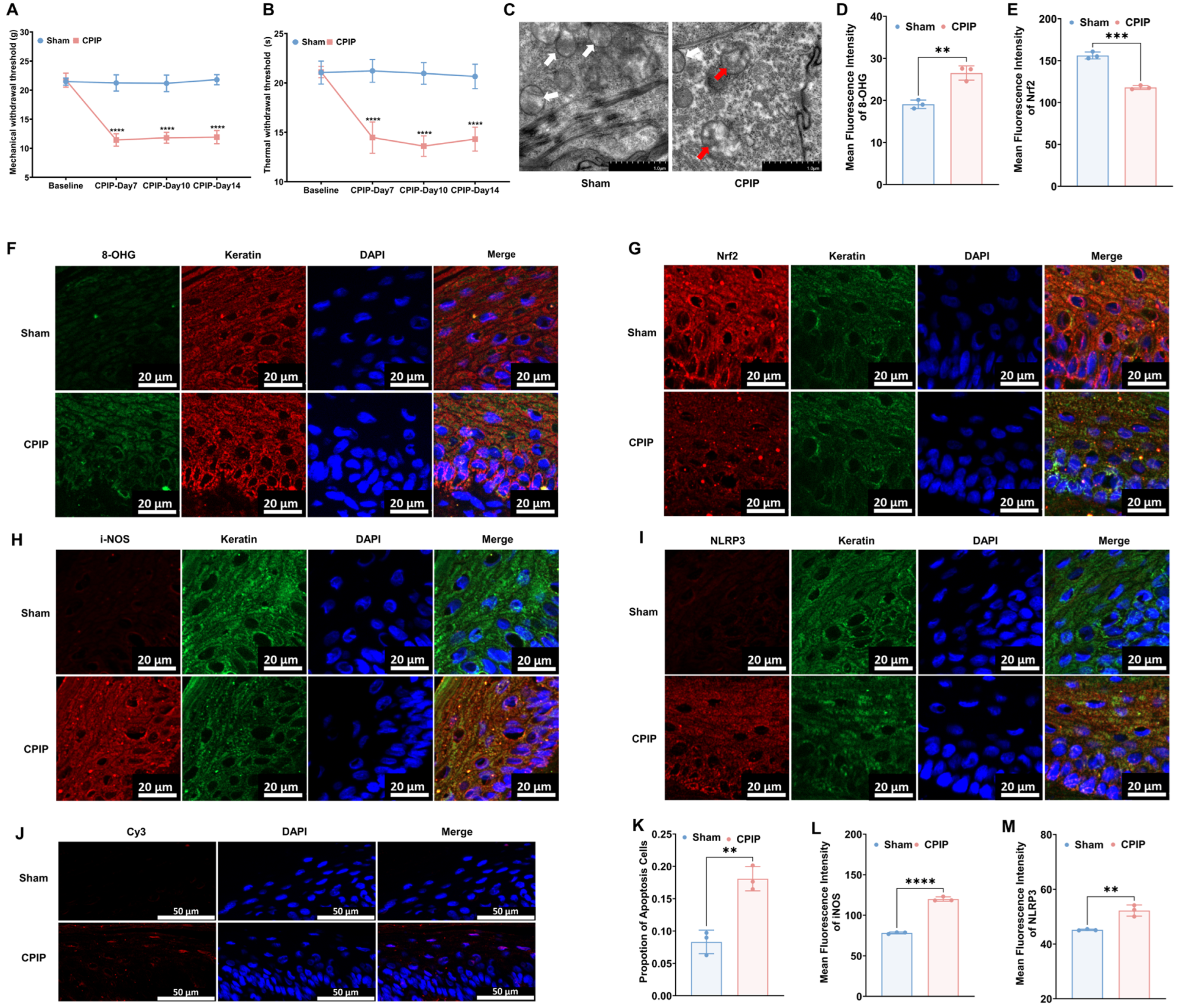
3.1. Keratinocytes in Male Rats with CPIP Show Oxidative Damage
3.2. OGD/R Induces Oxidative Damage in HaCaT Cells
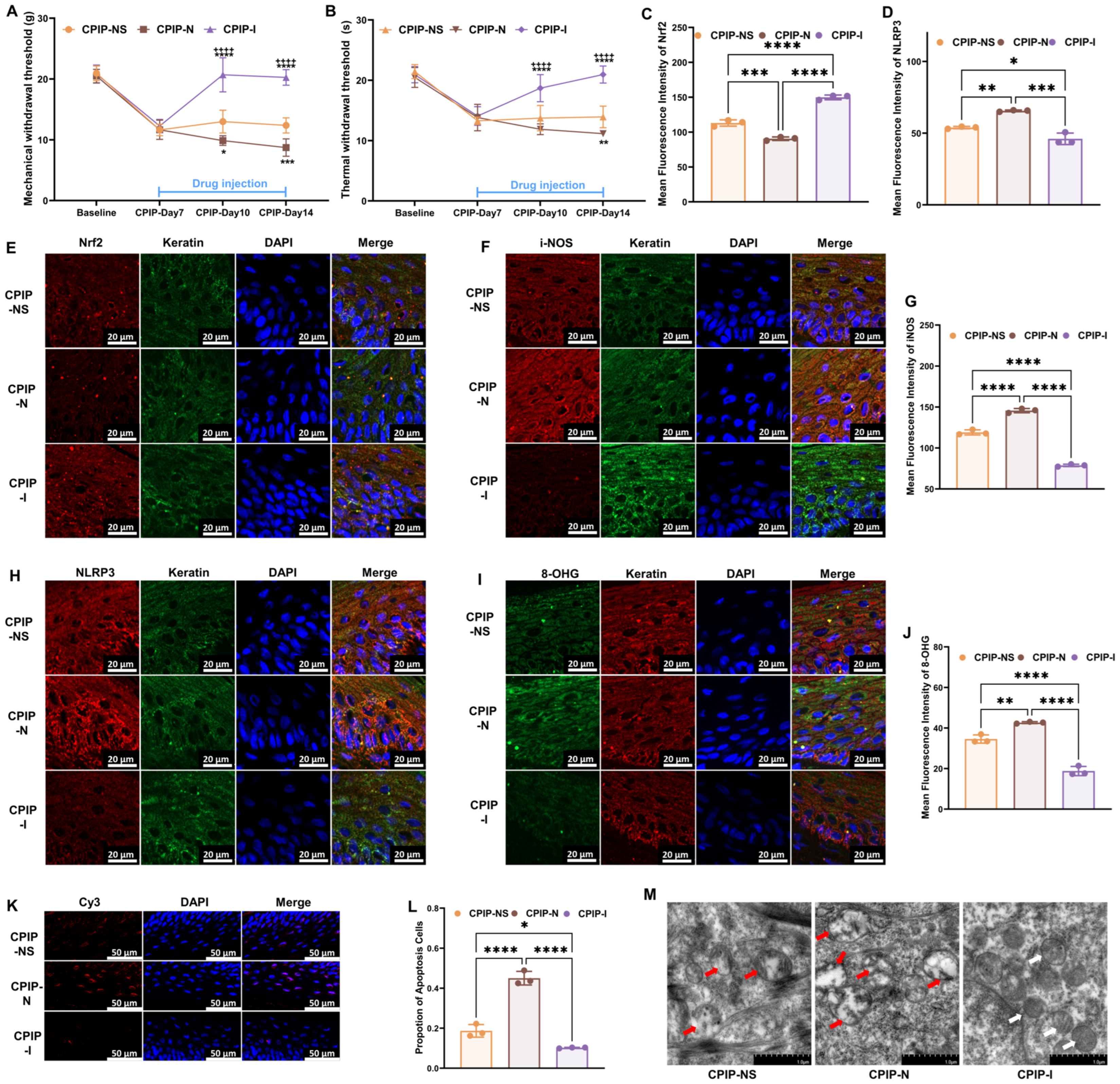
3.3. NMDA Receptors Modulate Oxidative Damage in Keratinocytes of CRPS
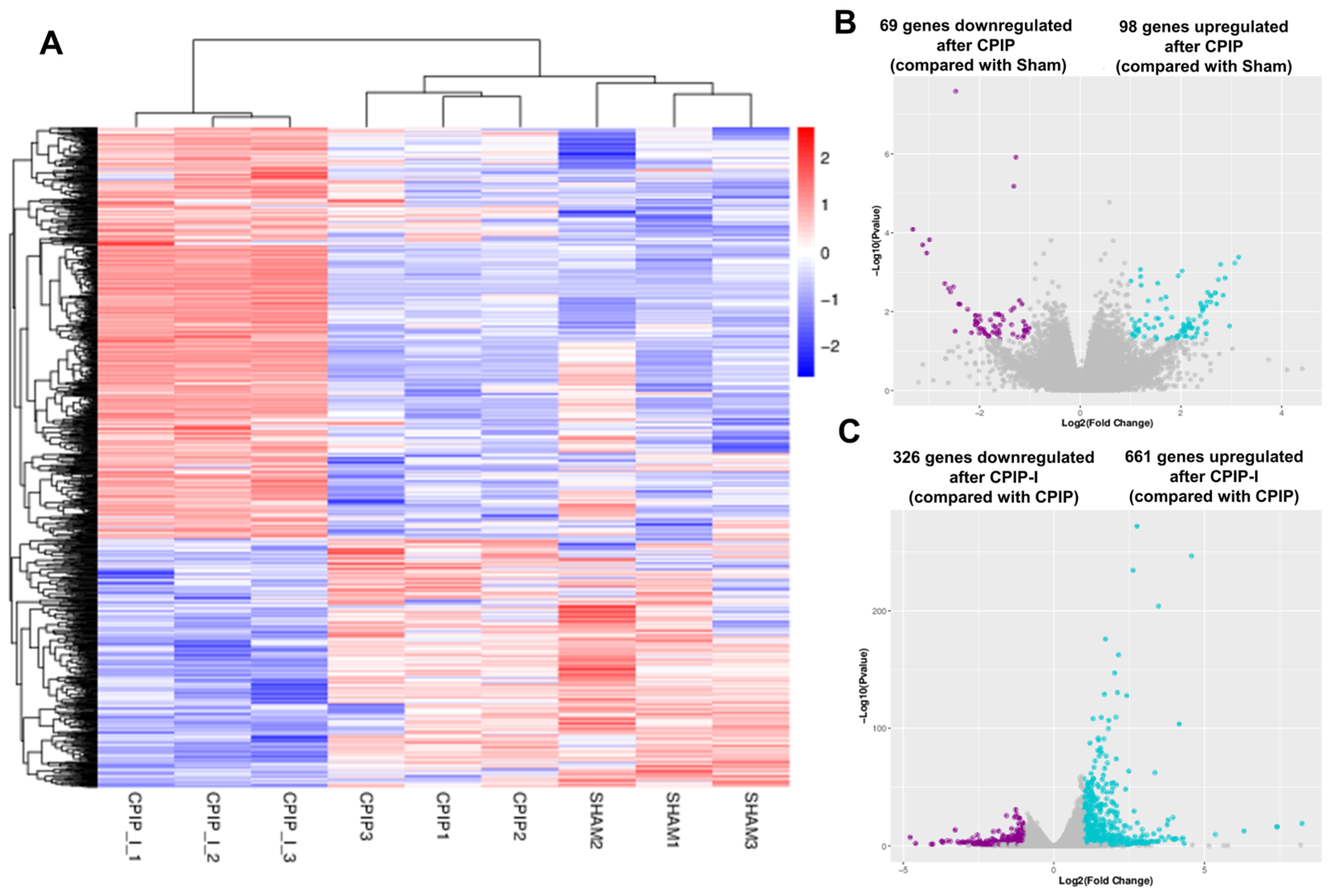
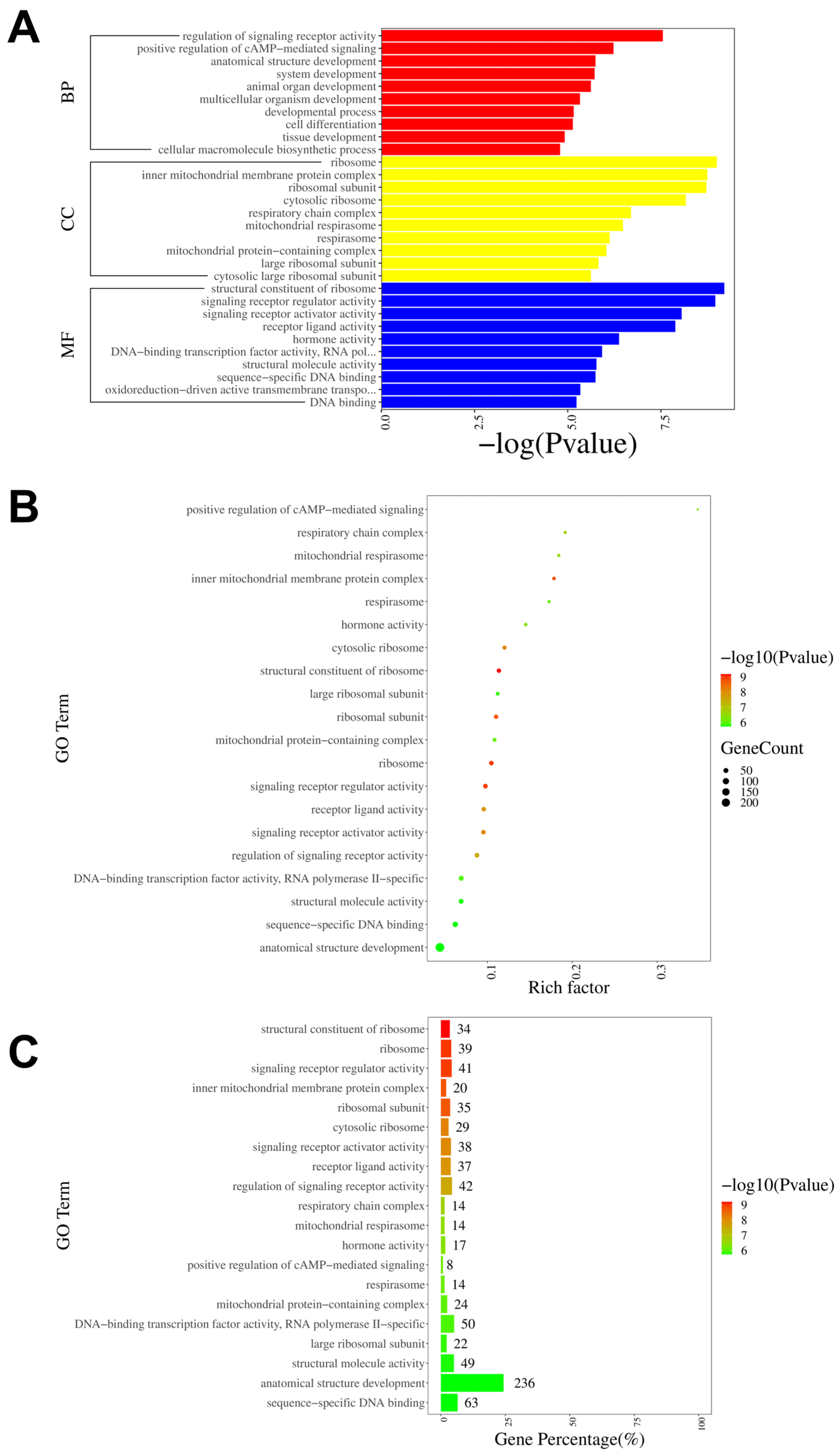
3.4. NR2B Induces Changes in the Gene Transcriptome Profiles of DRGs in Male Rats with CPIP
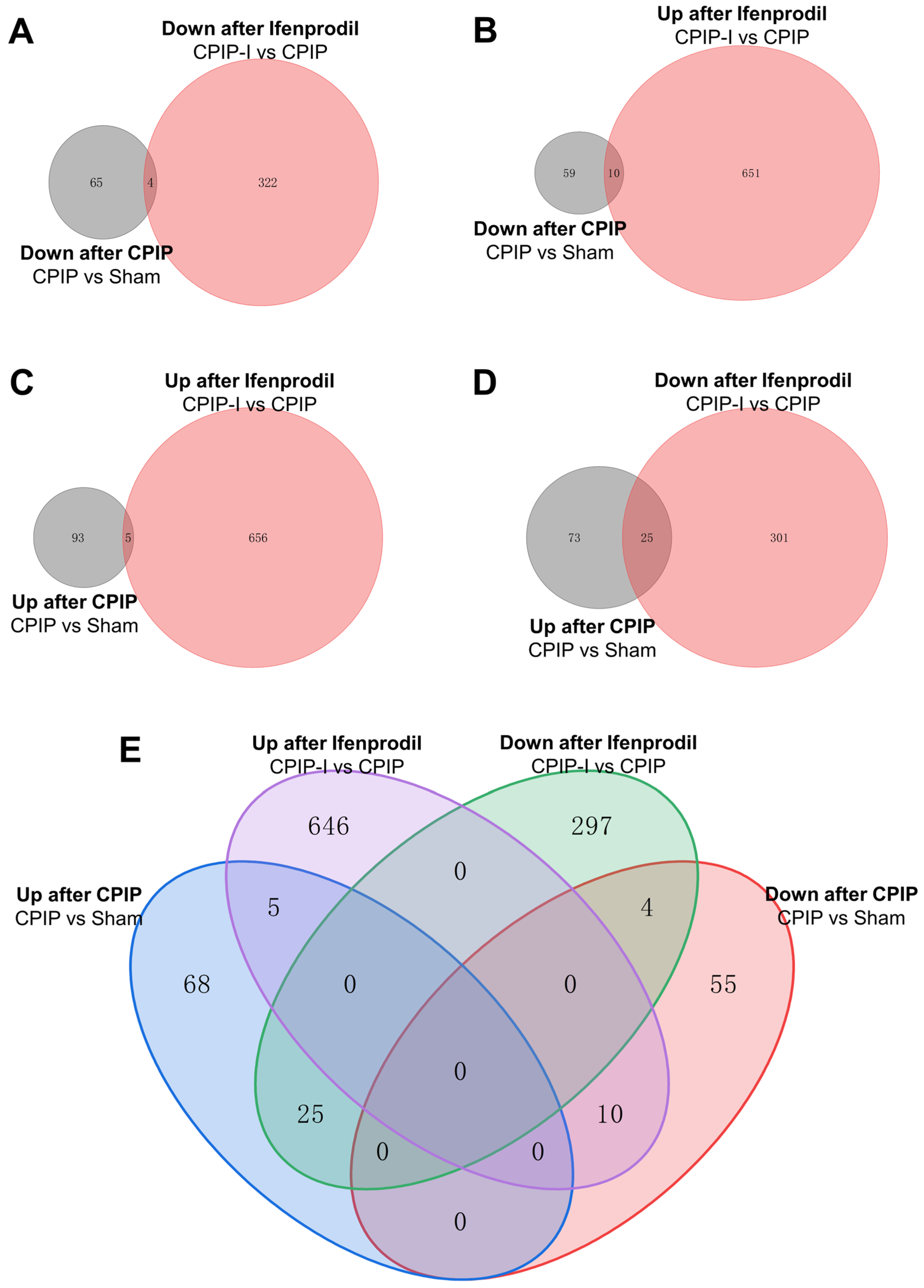
3.5. Analysis of DEGs in the DRGs of CPIP Male Rats after Intraplantar Injection of Ifenprodil
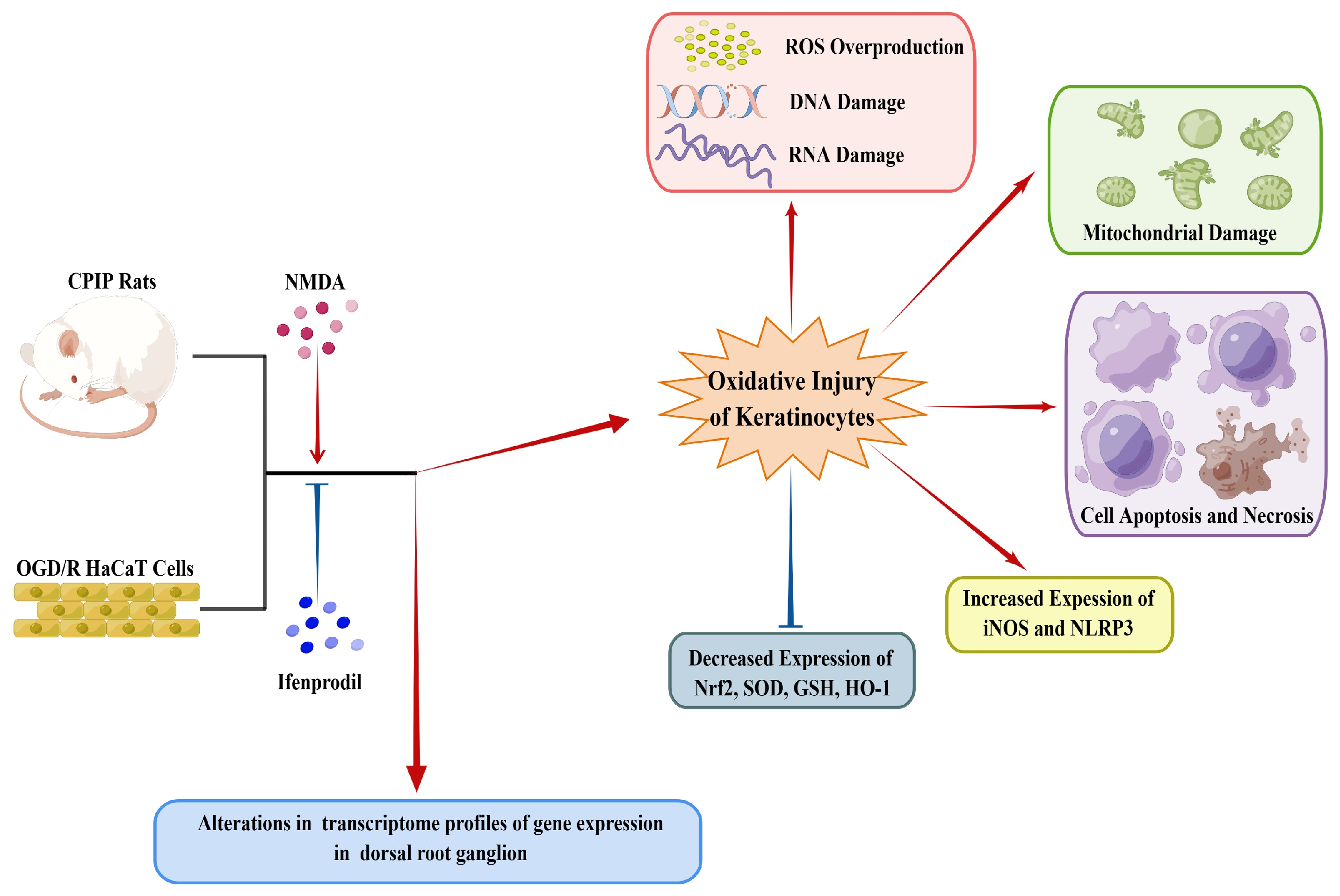
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicholas, M.; Vlaeyen, J.W.S.; Rief, W.; Barke, A.; Aziz, Q.; Benoliel, R.; Cohen, M.; Evers, S.; Giamberardino, M.A.; Goebel, A.; et al. The IASP classification of chronic pain for ICD-11: Chronic primary pain. Pain 2019, 160, 28–37. [Google Scholar] [CrossRef]
- David Clark, J.; Tawfik, V.L.; Tajerian, M.; Kingery, W.S. Autoinflammatory and autoimmune contributions to complex regional pain syndrome. Mol. Pain 2018, 14, 1744806918799127. [Google Scholar] [CrossRef]
- Birklein, F.; Ajit, S.K.; Goebel, A.; Perez, R.; Sommer, C. Complex regional pain syndrome—Phenotypic characteristics and potential biomarkers. Nat. Rev. Neurol. 2018, 14, 272–284. [Google Scholar] [CrossRef]
- Arcidiacono, U.A.; Armocida, D.; Pesce, A.; Maiotti, M.; Proietti, L.; D’Andrea, G.; Santoro, A.; Frati, A. Complex Regional Pain Syndrome after Spine Surgery: A Rare Complication in Mini-Invasive Lumbar Spine Surgery: An Updated Comprehensive Review. J. Clin. Med. 2022, 11, 7409. [Google Scholar] [CrossRef]
- Kessler, A.; Yoo, M.; Calisoff, R. Complex regional pain syndrome: An updated comprehensive review. NeuroRehabilitation 2020, 47, 253–264. [Google Scholar] [CrossRef]
- Shim, H.; Rose, J.; Halle, S.; Shekane, P. Complex regional pain syndrome: A narrative review for the practising clinician. Br. J. Anaesth. 2019, 123, e424–e433. [Google Scholar] [CrossRef]
- Baronio, M.; Sadia, H.; Paolacci, S.; Prestamburgo, D.; Miotti, D.; Guardamagna, V.A.; Natalini, G.; Sullivan, S.G.B.; Bertelli, M. Molecular Aspects of Regional Pain Syndrome. Pain Res. Manag. 2020, 2020, 7697214. [Google Scholar] [CrossRef]
- Zhu, H.; Wen, B.; Xu, L.; Huang, Y. Identification of Potential Inflammation-Related Genes and Key Pathways Associated with Complex Regional Pain Syndrome. Biomolecules 2023, 13, 772. [Google Scholar] [CrossRef]
- Wen, B.; Pan, Y.; Cheng, J.; Xu, L.; Xu, J. The Role of Neuroinflammation in Complex Regional Pain Syndrome: A Comprehensive Review. J. Pain Res. 2023, 16, 3061–3073. [Google Scholar] [CrossRef]
- Baron, R.; Hans, G.; Dickenson, A.H. Peripheral input and its importance for central sensitization. Ann. Neurol. 2013, 74, 630–636. [Google Scholar] [CrossRef]
- Kuner, R. Central mechanisms of pathological pain. Nat. Med. 2010, 16, 1258–1266. [Google Scholar] [CrossRef]
- Xu, X.; Yu, C.; Xu, L.; Xu, J. Emerging roles of keratinocytes in nociceptive transduction and regulation. Front. Mol. Neurosci. 2022, 15, 982202. [Google Scholar] [CrossRef]
- Xu, X.; Tao, X.; Huang, P.; Lin, F.; Liu, Q.; Xu, L.; Xu, J.; Huang, Y. N-methyl-d-aspartate receptor subunit 2B on keratinocyte mediates peripheral and central sensitization in chronic post-ischemic pain in male rats. Brain Behav. Immun. 2020, 87, 579–590. [Google Scholar] [CrossRef]
- Kaushik, A.S.; Strath, L.J.; Sorge, R.E. Dietary Interventions for Treatment of Chronic Pain: Oxidative Stress and Inflammation. Pain Ther. 2020, 9, 487–498. [Google Scholar] [CrossRef]
- Hsiao, H.T.; Lin, Y.C.; Wang, J.C.; Tsai, Y.C.; Liu, Y.C. Hypoxia inducible factor-1α inhibition produced anti-allodynia effect and suppressed inflammatory cytokine production in early stage of mouse complex regional pain syndrome model. Clin. Exp. Pharmacol. Physiol. 2016, 43, 355–359. [Google Scholar] [CrossRef]
- Li, X.; Yin, C.; Hu, Q.; Wang, J.; Nie, H.; Liu, B.; Tai, Y.; Fang, J.; Du, J.; Shao, X.; et al. Nrf2 Activation Mediates Antiallodynic Effect of Electroacupuncture on a Rat Model of Complex Regional Pain Syndrome Type-I through Reducing Local Oxidative Stress and Inflammation. Oxid. Med. Cell. Longev. 2022, 2022, 8035109. [Google Scholar] [CrossRef]
- Coderre, T.J.; Xanthos, D.N.; Francis, L.; Bennett, G.J. Chronic post-ischemia pain (CPIP): A novel animal model of complex regional pain syndrome-type I (CRPS-I; reflex sympathetic dystrophy) produced by prolonged hindpaw ischemia and reperfusion in the rat. Pain 2004, 112, 94–105. [Google Scholar] [CrossRef]
- Gao, X.; Chen, F.; Xu, X.; Liu, J.; Dong, F.; Liu, Y. Ro25-6981 alleviates neuronal damage and improves cognitive deficits by attenuating oxidative stress via the Nrf2/ARE pathway in ischemia/reperfusion rats. J. Stroke Cerebrovasc. Dis. 2022, 32, 106971. [Google Scholar] [CrossRef]
- Fischer, G.; Mutel, V.; Trube, G.; Malherbe, P.; Kew, J.N.; Mohacsi, E.; Heitz, M.P.; Kemp, J.A. Ro 25-6981, a highly potent and selective blocker of N-methyl-D-aspartate receptors containing the NR2B subunit. Characterization in vitro. J. Pharmacol. Exp. Ther. 1997, 283, 1285–1292. [Google Scholar]
- Yang, S.; Park, S.H.; Oh, S.W.; Kwon, K.; Yu, E.; Lee, C.W.; Son, Y.K.; Kim, C.; Lee, B.H.; Cho, J.Y.; et al. Antioxidant Activities and Mechanisms of Tomentosin in Human Keratinocytes. Antioxidants 2022, 11, 990. [Google Scholar] [CrossRef]
- Jiang, B.W.; Zhang, W.J.; Wang, Y.; Tan, L.P.; Bao, Y.L.; Song, Z.B.; Yu, C.L.; Wang, S.Y.; Liu, L.; Li, Y.X. Convallatoxin induces HaCaT cell necroptosis and ameliorates skin lesions in psoriasis-like mouse models. Biomed. Pharmacother. 2020, 121, 109615. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, C.H.; Hwang, B.S.; Choi, K.M.; Yang, I.J.; Kim, G.Y.; Choi, Y.H.; Park, C.; Jeong, J.W. Protective Effects of Oenothera biennis against Hydrogen Peroxide-Induced Oxidative Stress and Cell Death in Skin Keratinocytes. Life 2020, 10, 255. [Google Scholar] [CrossRef]
- Kim, J.S.; Lim, S.S. LED Light-Induced ROS Differentially Regulates Focal Adhesion Kinase Activity in HaCaT Cell Viability. Curr. Issues Mol. Biol. 2022, 44, 1235–1246. [Google Scholar] [CrossRef]
- Bruehl, S.; Warner, D.S. An Update on the Pathophysiology of Complex Regional Pain Syndrome. Anesthesiology 2010, 113, 713–725. [Google Scholar] [CrossRef]
- Ragavendran, J.V.; Laferrière, A.; Khorashadi, M.; Coderre, T.J. Pentoxifylline reduces chronic post-ischaemia pain by alleviating microvascular dysfunction. Eur. J. Pain 2014, 18, 406–414. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, L.; Xu, D.; Zhang, W.; Zhang, Y.; Zhou, J.; Huang, W. Interaction between astrocytic colony stimulating factor and its receptor on microglia mediates central sensitization and behavioral hypersensitivity in chronic post ischemic pain model. Brain Behav. Immun. 2018, 68, 248–260. [Google Scholar] [CrossRef]
- Liu, Y.; Liang, Y.; Gao, M.; Li, Y.; Zhao, T.; Zhao, Y. Animal Models of Complex Regional Pain Syndrome Type I. J. Pain Res. 2021, 14, 3711–3721. [Google Scholar] [CrossRef]
- Arifin, W.N.; Zahiruddin, W.M. Sample Size Calculation in Animal Studies Using Resource Equation Approach. Malays. J. Med. Sci. 2017, 24, 101–105. [Google Scholar] [CrossRef]
- Sadler, K.E.; Patitucci, T.N.; Stucky, C.L. Ex Vivo Skin-Teased Fiber Recordings from Tibial NerveTibial nerves. In Contemporary Approaches to the Study of Pain: From Molecules to Neural Networks; Seal, R.P., Ed.; Springer: New York, NY, USA, 2022; pp. 71–95. [Google Scholar]
- Liu, B.; Tai, Y.; Caceres, A.I.; Achanta, S.; Balakrishna, S.; Shao, X.; Fang, J.; Jordt, S.E. Oxidized Phospholipid OxPAPC Activates TRPA1 and Contributes to Chronic Inflammatory Pain in Mice. PLoS ONE 2016, 11, e0165200. [Google Scholar] [CrossRef]
- Shi, Y.; Yuan, S.; Tang, S.J. Reactive Oxygen Species (ROS) are Critical for Morphine Exacerbation of HIV-1 gp120-Induced Pain. J. Neuroimmune Pharmacol. 2021, 16, 581–591. [Google Scholar] [CrossRef]
- Bittar, A.; Jun, J.; La, J.H.; Wang, J.; Leem, J.W.; Chung, J.M. Reactive oxygen species affect spinal cell type-specific synaptic plasticity in a model of neuropathic pain. Pain 2017, 158, 2137–2146. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, J.; Zhang, X.; Guo, Z.; Wu, W.; Chen, Z.; Li, J. Reactive Oxygen Species Mediate Low Back Pain by Upregulating Substance P in Intervertebral Disc Degeneration. Oxid. Med. Cell. Longev. 2021, 2021, 6681815. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, J. Sanggenon C Ameliorates Cerebral Ischemia-Reperfusion Injury by Inhibiting Inflammation and Oxidative Stress through Regulating RhoA-ROCK Signaling. Inflammation 2020, 43, 1476–1487. [Google Scholar] [CrossRef]
- Li, S.; Jiang, D.; Rosenkrans, Z.T.; Barnhart, T.E.; Ehlerding, E.B.; Ni, D.; Engle, J.W.; Cai, W. Aptamer-Conjugated Framework Nucleic Acids for the Repair of Cerebral Ischemia-Reperfusion Injury. Nano Lett. 2019, 19, 7334–7341. [Google Scholar] [CrossRef]
- Mukherjee, A.; Sarkar, S.; Jana, S.; Swarnakar, S.; Das, N. Neuro-protective role of nanocapsulated curcumin against cerebral ischemia-reperfusion induced oxidative injury. Brain Res. 2019, 1704, 164–173. [Google Scholar] [CrossRef]
- He, R.; Jiang, Y.; Shi, Y.; Liang, J.; Zhao, L. Curcumin-laden exosomes target ischemic brain tissue and alleviate cerebral ischemia-reperfusion injury by inhibiting ROS-mediated mitochondrial apoptosis. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 117, 111314. [Google Scholar] [CrossRef]
- Groeneweg, G.; Huygen, F.J.; Coderre, T.J.; Zijlstra, F.J. Regulation of peripheral blood flow in complex regional pain syndrome: Clinical implication for symptomatic relief and pain management. BMC Musculoskelet. Disord. 2009, 10, 116. [Google Scholar] [CrossRef]
- Yang, J.; Qi, J.; Xiu, B.; Yang, B.; Niu, C.; Yang, H. Reactive Oxygen Species Play a Biphasic Role in Brain Ischemia. J. Investig. Surg. 2019, 32, 97–102. [Google Scholar] [CrossRef]
- Coderre, T.J.; Bennett, G.J. A hypothesis for the cause of complex regional pain syndrome-type I (reflex sympathetic dystrophy): Pain due to deep-tissue microvascular pathology. Pain Med. 2010, 11, 1224–1238. [Google Scholar] [CrossRef]
- Klafke, J.Z.; da Silva, M.A.; Rossato, M.F.; de Prá, S.D.; Rigo, F.K.; Walker, C.I.; Bochi, G.V.; Moresco, R.N.; Ferreira, J.; Trevisan, G. Acute and chronic nociceptive phases observed in a rat hind paw ischemia/reperfusion model depend on different mechanisms. Pflugers Arch. 2016, 468, 229–241. [Google Scholar] [CrossRef]
- Motterlini, R.; Foresti, R. Heme oxygenase-1 as a target for drug discovery. Antioxid. Redox Signal. 2014, 20, 1810–1826. [Google Scholar] [CrossRef]
- Kesherwani, V.; Atif, F.; Yousuf, S.; Agrawal, S.K. Resveratrol protects spinal cord dorsal column from hypoxic injury by activating Nrf-2. Neuroscience 2013, 241, 80–88. [Google Scholar] [CrossRef]
- Shaw, P.; Chattopadhyay, A. Nrf2-ARE signaling in cellular protection: Mechanism of action and the regulatory mechanisms. J. Cell. Physiol. 2020, 235, 3119–3130. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Paoletti, P.; Bellone, C.; Zhou, Q. NMDA receptor subunit diversity: Impact on receptor properties, synaptic plasticity and disease. Nat. Rev. Neurosci. 2013, 14, 383–400. [Google Scholar] [CrossRef]
- Zhu, X.; Dong, J.; Shen, K.; Bai, Y.; Zhang, Y.; Lv, X.; Chao, J.; Yao, H. NMDA receptor NR2B subunits contribute to PTZ-kindling-induced hippocampal astrocytosis and oxidative stress. Brain Res. Bull. 2015, 114, 70–78. [Google Scholar] [CrossRef]
- Uçeyler, N.; Eberle, T.; Rolke, R.; Birklein, F.; Sommer, C. Differential expression patterns of cytokines in complex regional pain syndrome. Pain 2007, 132, 195–205. [Google Scholar] [CrossRef]
- Sandireddy, R.; Yerra, V.G.; Areti, A.; Komirishetty, P.; Kumar, A. Neuroinflammation and oxidative stress in diabetic neuropathy: Futuristic strategies based on these targets. Int. J. Endocrinol. 2014, 2014, 674987. [Google Scholar] [CrossRef]
- Sibille, K.T.; Steingrímsdóttir, Ó.A.; Fillingim, R.B.; Stubhaug, A.; Schirmer, H.; Chen, H.; McEwen, B.S.; Nielsen, C.S. Investigating the Burden of Chronic Pain: An Inflammatory and Metabolic Composite. Pain Res. Manag. 2016, 2016, 7657329. [Google Scholar] [CrossRef]
- Allison, D.J.; Thomas, A.; Beaudry, K.; Ditor, D.S. Targeting inflammation as a treatment modality for neuropathic pain in spinal cord injury: A randomized clinical trial. J. Neuroinflamm. 2016, 13, 152. [Google Scholar] [CrossRef]
- Hannig, G.; Tchernychev, B.; Kurtz, C.B.; Bryant, A.P.; Currie, M.G.; Silos-Santiago, I. Guanylate cyclase-C/cGMP: An emerging pathway in the regulation of visceral pain. Front. Mol. Neurosci. 2014, 7, 31. [Google Scholar] [CrossRef]
- Silos-Santiago, I.; Hannig, G.; Eutamene, H.; Ustinova, E.E.; Bernier, S.G.; Ge, P.; Graul, C.; Jacobson, S.; Jin, H.; Liong, E.; et al. Gastrointestinal pain: Unraveling a novel endogenous pathway through uroguanylin/guanylate cyclase-C/cGMP activation. Pain 2013, 154, 1820–1830. [Google Scholar] [CrossRef]
- Huang, X.; Deng, J.; Xu, T.; Xin, W.; Zhang, Y.; Ruan, X. Downregulation of metallothionein-2 contributes to oxaliplatin-induced neuropathic pain. J. Neuroinflamm. 2021, 18, 91. [Google Scholar] [CrossRef]
- Oki, G.; Wada, T.; Iba, K.; Aiki, H.; Sasaki, K.; Imai, S.I.; Sohma, H.; Matsumoto, K.; Yamaguchi, M.; Fujimiya, M.; et al. Metallothionein deficiency in the injured peripheral nerves of complex regional pain syndrome as revealed by proteomics. Pain 2012, 153, 532–539. [Google Scholar] [CrossRef]
- Heo, J.H.; Lee, S.H.; Chang, K.H.; Han, E.H.; Lee, S.G.; Choi, D.W.; Kim, S.W. Identification of differentially expressed genes by gabapentin in cultured dorsal root ganglion in a rat neuropathic pain model. Biomol. Ther. 2013, 21, 126–131. [Google Scholar] [CrossRef][Green Version]
- Zhang, J.; Mei, Z.; Yao, W.; Zhao, C.; Wu, S.; Ouyang, J. SIX1 induced HULC modulates neuropathic pain and Schwann cell oxidative stress after sciatic nerve injury. Gene 2023, 882, 147655. [Google Scholar] [CrossRef]
- Lee, E.; Takita, C.; Wright, J.L.; Slifer, S.H.; Martin, E.R.; Urbanic, J.J.; Langefeld, C.D.; Lesser, G.J.; Shaw, E.G.; Hu, J.J. Genome-wide enriched pathway analysis of acute post-radiotherapy pain in breast cancer patients: A prospective cohort study. Hum. Genom. 2019, 13, 28. [Google Scholar] [CrossRef]
- Chen, J.; Guo, P.; Liu, X.; Liao, H.; Chen, K.; Wang, Y.; Qin, J.; Yang, F. Sinomenine alleviates diabetic peripheral neuropathic pain through inhibition of the inositol-requiring enzyme 1 alpha-X-box binding protein 1 pathway by downregulating prostaglandin-endoperoxide synthase 2. J. Diabetes Investig. 2023, 14, 364–375. [Google Scholar] [CrossRef]
- Husain, S.F.; Lam, R.W.M.; Hu, T.; Ng, M.W.F.; Liau, Z.Q.G.; Nagata, K.; Khanna, S.; Lam, Y.; Bhakoo, K.; Ho, R.C.M.; et al. Locating the Site of Neuropathic Pain In Vivo Using MMP-12-Targeted Magnetic Nanoparticles. Pain Res. Manag. 2019, 2019, 9394715. [Google Scholar] [CrossRef]
- Lim, E.F.; Hoghooghi, V.; Hagen, K.M.; Kapoor, K.; Frederick, A.; Finlay, T.M.; Ousman, S.S. Presence and activation of pro-inflammatory macrophages are associated with CRYAB expression in vitro and after peripheral nerve injury. J. Neuroinflamm. 2021, 18, 82. [Google Scholar] [CrossRef]
- Cui, C.Y.; Liu, X.; Peng, M.H.; Liu, Q.; Zhang, Y. Identification of key candidate genes and biological pathways in neuropathic pain. Comput. Biol. Med. 2022, 150, 106135. [Google Scholar] [CrossRef]
- Sun, W.; Yang, S.; Wu, S.; Ba, X.; Xiong, D.; Xiao, L.; Hao, Y. Transcriptome analysis reveals dysregulation of inflammatory and neuronal function in dorsal root ganglion of paclitaxel-induced peripheral neuropathy rats. Mol. Pain 2023, 19, 17448069221106167. [Google Scholar] [CrossRef]
- Du, Z.; Yin, S.; Song, X.; Zhang, L.; Yue, S.; Jia, X.; Zhang, Y. Identification of Differentially Expressed Genes and Key Pathways in the Dorsal Root Ganglion After Chronic Compression. Front. Mol. Neurosci. 2020, 13, 71. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene_ID | Gene Symbol | Full Gene Name | Log2 Fold Change (CPIP-I/CPIP) | p Value |
|---|---|---|---|---|
| ENSRNOG00000047545 | Adra2a | adrenoceptor alpha 2A | 1.05 | 2.40 × 10−7 |
| ENSRNOG00000017002 | Adrb1 | adrenoceptor beta 1 | 1.89 | 5.76 × 10−6 |
| ENSRNOG00000012674 | Adrb3 | adrenoceptor beta 3 | 2.41 | 4.00 × 10−3 |
| ENSRNOG00000011074 | Calcb | calcitonin-related polypeptide, beta | 1.02 | 1.13 × 10−21 |
| ENSRNOG00000019321 | Cck | cholecystokinin | 1.63 | 1.04 × 10−4 |
| ENSRNOG00000017556 | Chrm4 | cholinergic receptor, muscarinic 4 | 1.14 | 3.32 × 10−6 |
| ENSRNOG00000006397 | Chrm5 | cholinergic receptor, muscarinic 5 | −1.17 | 2.03 × 10−2 |
| ENSRNOG00000012703 | Crh | corticotropin-releasing hormone | 1.28 | 1.45 × 10−2 |
| ENSRNOG00000011145 | Crhr2 | corticotropin-releasing hormone receptor 2 | 1.16 | 1.84 × 10−4 |
| ENSRNOG00000008428 | Drd2 | dopamine receptor D2 | 1.13 | 4.47 × 10−3 |
| ENSRNOG00000013433 | Gnrh1 | gonadotropin-releasing hormone 1 | −2.06 | 1.91 × 10−2 |
| ENSRNOG00000024030 | Gpr35 | G protein-coupled receptor 35 | 1.24 | 6.08 × 10−7 |
| ENSRNOG00000010254 | Htr1a | 5-hydroxytryptamine receptor 1A | 1.32 | 1.88 × 10−4 |
| ENSRNOG00000002549 | Htr5b | 5-hydroxytryptamine (serotonin) receptor 5B | 1.02 | 2.95 × 10−4 |
| ENSRNOG00000047040 | Lhb | luteinizing hormone beta polypeptide | 1.20 | 1.93 × 10−2 |
| ENSRNOG00000015260 | Lpar3 | lysophosphatidic acid receptor 3 | 1.13 | 2.91 × 10−3 |
| ENSRNOG00000036685 | Npb | neuropeptide B | 1.91 | 2.38 × 10−6 |
| ENSRNOG00000012390 | Npw | neuropeptide W | 1.59 | 7.06 × 10−5 |
| ENSRNOG00000009768 | Npy | neuropeptide Y | 3.95 | 5.88 × 10−25 |
| ENSRNOG00000004179 | Nts | neurotensin | −1.18 | 3.36 × 10−3 |
| ENSRNOG00000037839 | P2ry10 | purinergic receptor P2Y10 | −1.19 | 3.31 × 10−2 |
| ENSRNOG00000014231 | Pnoc | prepronociceptin | 1.04 | 3.74 × 10−3 |
| ENSRNOG00000009922 | Prlhr | prolactin releasing hormone receptor | 1.76 | 1.94 × 10−2 |
| ENSRNOG00000031535 | Ptgdr | prostaglandin D2 receptor | 1.34 | 3.09 × 10−3 |
| ENSRNOG00000031307 | Ptgdrl | prostaglandin D2 receptor-like | 1.47 | 3.93 × 10−8 |
| ENSRNOG00000000897 | Rxfp2 | relaxin/insulin-like family peptide receptor 2 | −2.65 | 8.32× 10−3 |
| ENSRNOG00000023126 | Rxfp3 | relaxin/insulin-like family peptide receptor 3 | 1.95 | 2.04 × 10−2 |
| ENSRNOG00000005370 | S1pr4 | sphingosine-1-phosphate receptor 4 | 1.16 | 4.59 × 10−2 |
| ENSRNOG00000020901 | S1pr5 | sphingosine-1-phosphate receptor 5 | 1.57 | 8.75 × 10−3 |
| ENSRNOG00000001837 | Sst | somatostatin | 2.02 | 2.56 × 10−6 |
| ENSRNOG00000004641 | Sstr4 | somatostatin receptor 4 | 1.09 | 4.59 × 10−3 |
| ENSRNOG00000003972 | Tshr | thyroid-stimulating hormone receptor | 1.50 | 3.23 × 10−2 |
| ENSRNOG00000006090 | Ucn | urocortin | 1.99 | 3.64 × 10−2 |
| ENSRNOG00000001416 | Vgf | VGF nerve growth factor inducible | 1.18 | 6.99 × 10−7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, B.; Zhu, H.; Xu, J.; Xu, L.; Huang, Y. NMDA Receptors Regulate Oxidative Damage in Keratinocytes during Complex Regional Pain Syndrome in HaCaT Cells and Male Rats. Antioxidants 2024, 13, 244. https://doi.org/10.3390/antiox13020244
Wen B, Zhu H, Xu J, Xu L, Huang Y. NMDA Receptors Regulate Oxidative Damage in Keratinocytes during Complex Regional Pain Syndrome in HaCaT Cells and Male Rats. Antioxidants. 2024; 13(2):244. https://doi.org/10.3390/antiox13020244
Chicago/Turabian StyleWen, Bei, He Zhu, Jijun Xu, Li Xu, and Yuguang Huang. 2024. "NMDA Receptors Regulate Oxidative Damage in Keratinocytes during Complex Regional Pain Syndrome in HaCaT Cells and Male Rats" Antioxidants 13, no. 2: 244. https://doi.org/10.3390/antiox13020244
APA StyleWen, B., Zhu, H., Xu, J., Xu, L., & Huang, Y. (2024). NMDA Receptors Regulate Oxidative Damage in Keratinocytes during Complex Regional Pain Syndrome in HaCaT Cells and Male Rats. Antioxidants, 13(2), 244. https://doi.org/10.3390/antiox13020244