

Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Ameliorate Aging-Induced BTB Impairment in Porcine Testes by Activating Autophagy and Inhibiting ROS/NLRP3 Inflammasomes via the AMPK/mTOR Signaling Pathway

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Porcine Testes
2.2. Porcine BMSC Separation, Character Determination, and Differentiation
2.3. Porcine SC Segregation and Culture
2.4. Purification and Identification of Exos
2.5. PKH26 Staining for Exos
2.6. Cell Treatment
2.7. Measurement of Autophagic Flux
2.8. Senescence-Associated β-Galactosidase (SA-β-Gal) Staining
2.9. ROS and Antioxidant Assessment
2.10. IL-1β Determination
2.11. Animal Grouping and Age Modeling
2.12. Western Blotting Analysis
2.13. Histological Analysis
2.14. Immunohistochemical Staining
2.15. Evaluation of Oxidative Stress in Testes
2.16. Transmission Electron Microscopy (TEM)
2.17. Data Analysis
3. Results
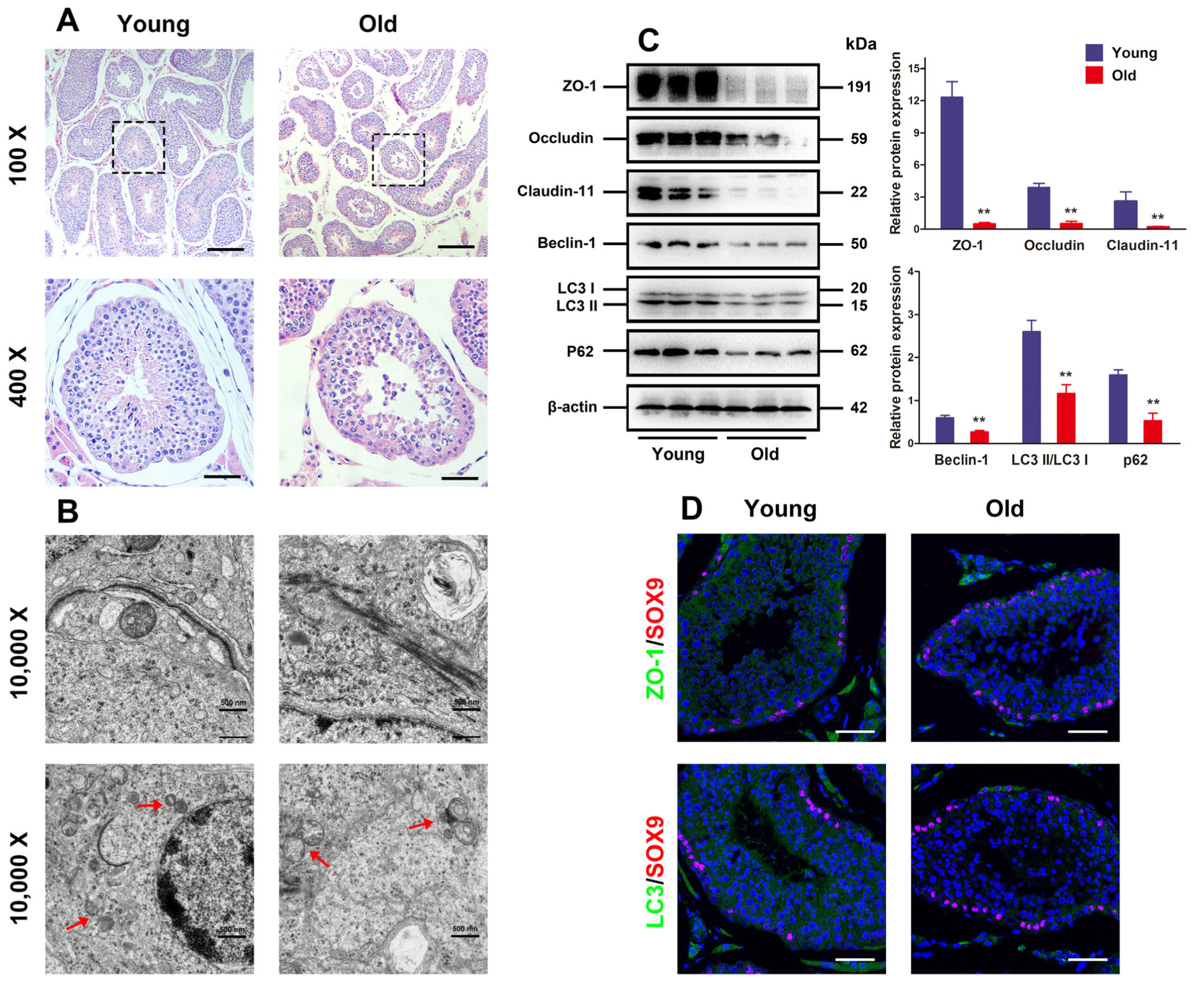
3.1. Autophagy Defect and BTB Dysfunction in Old Porcine Testes
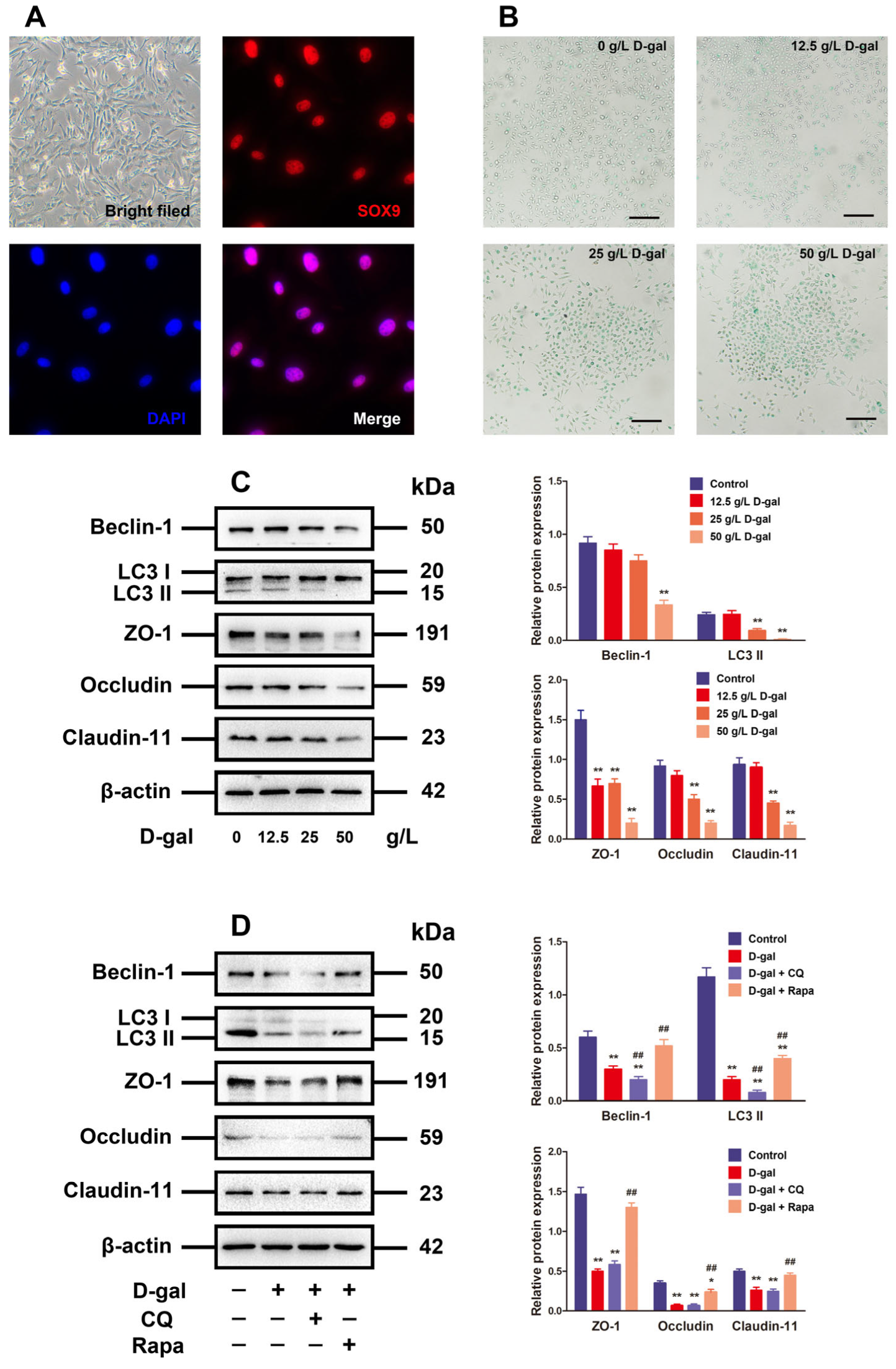
3.2. Alteration of Autophagy Affected TJ Function in D-Gal-Induced Aging Porcine SCs
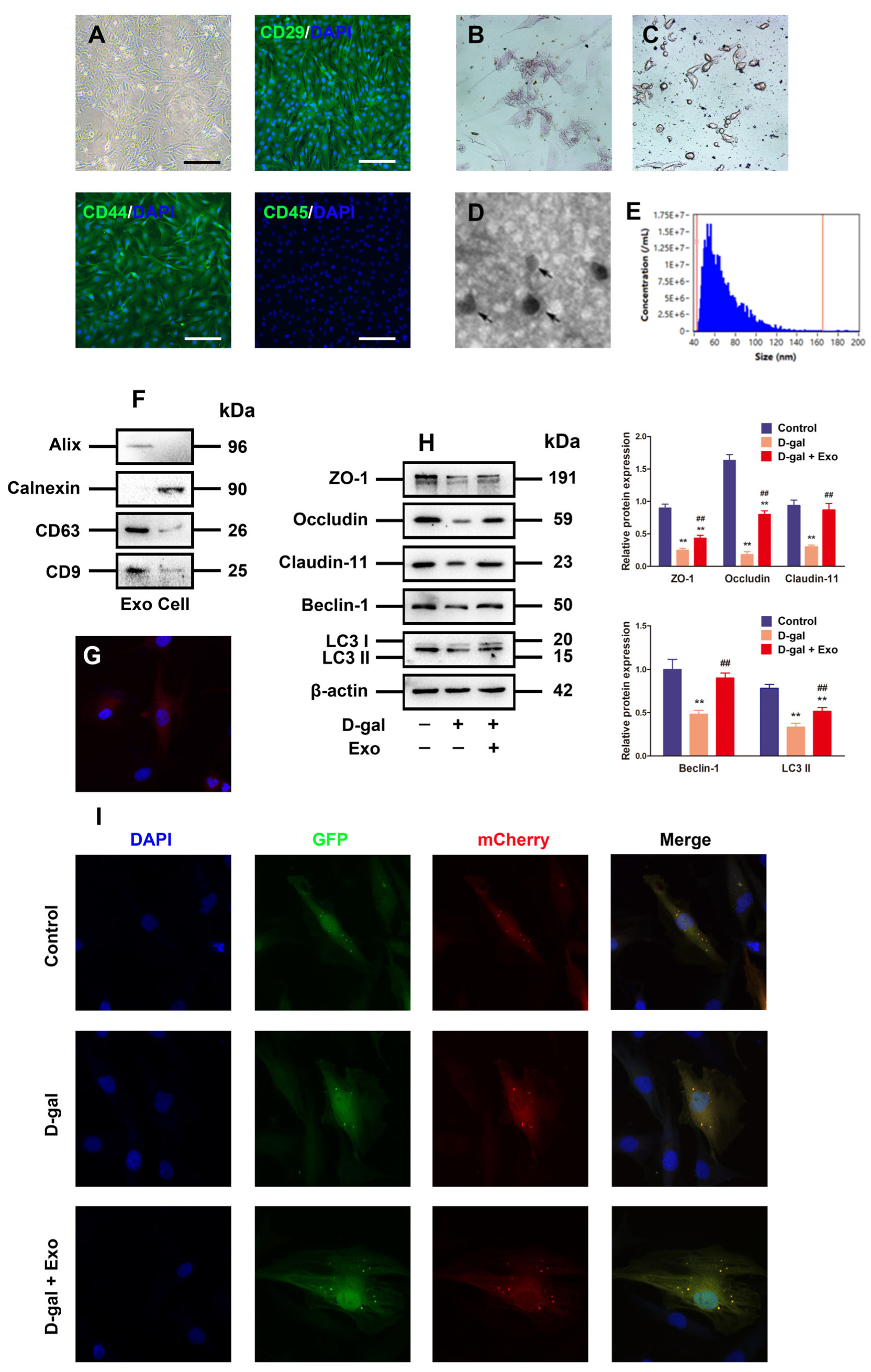
3.3. Characterization of BMSC-Exos and Exos Inhibited D-Gal-Induced Autophagy Defect and TJ Dysfunction in Porcine SCs
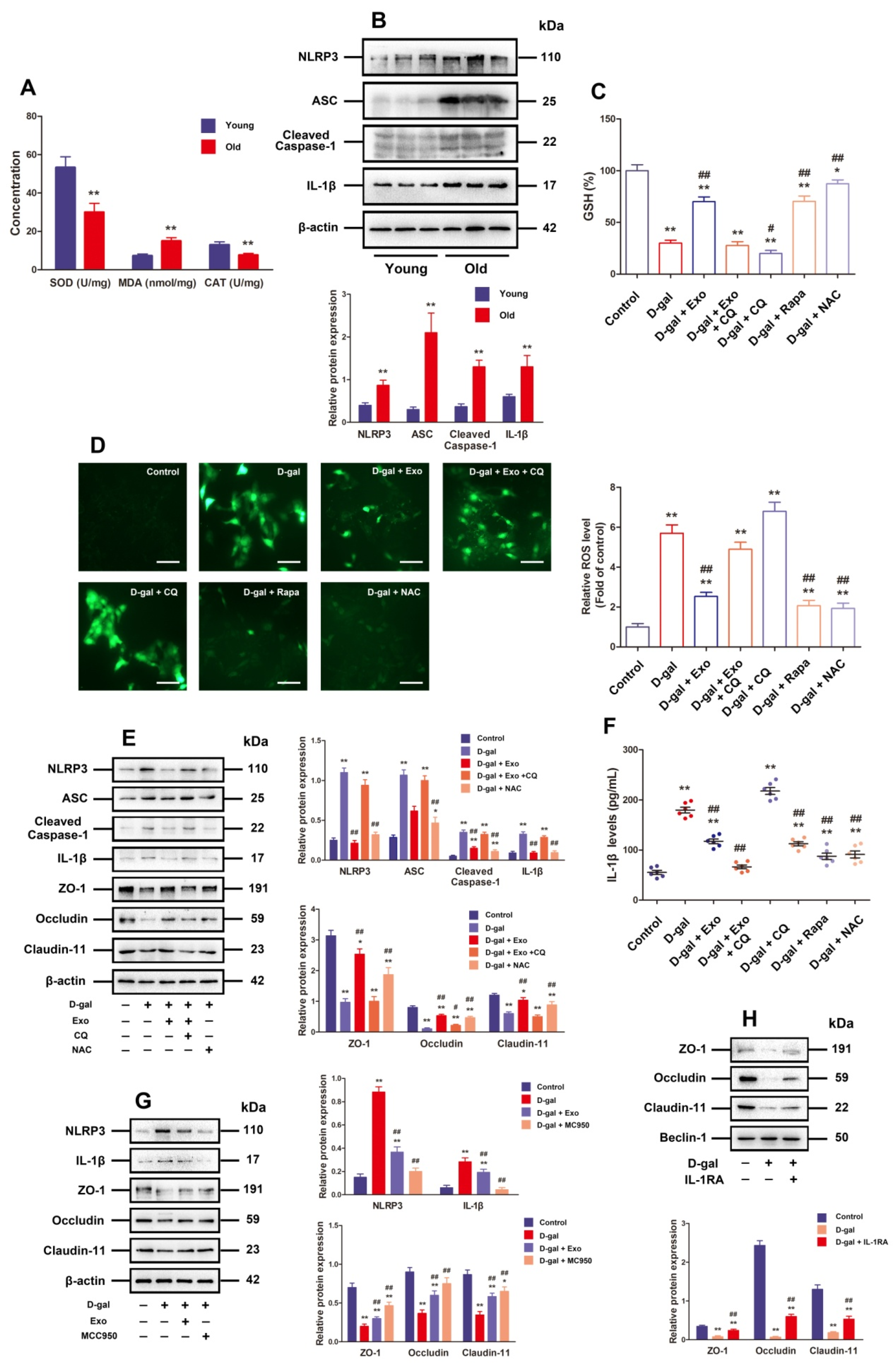
3.4. BMSC-Exos Recovered TJ Function through Promoting Autophagic Flux to Inhibit NLRP3 Inflammasome Activation Plus ROS Generation in D-Gal-Induced Aging Porcine SCs
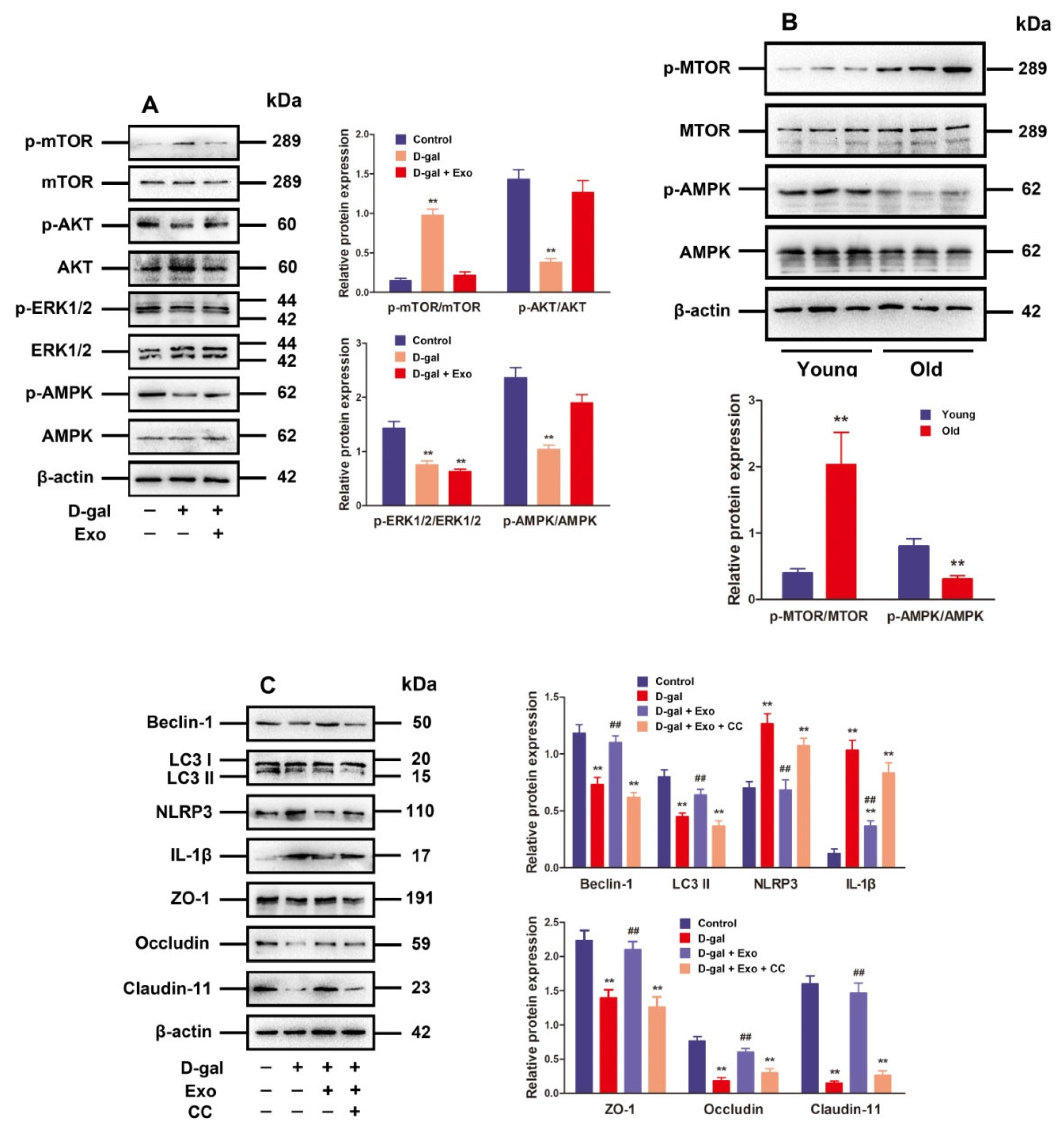
3.5. BMSC-Exos Regulated Autophagy and TJ Function through the AMPK/mTOR Signaling Pathway
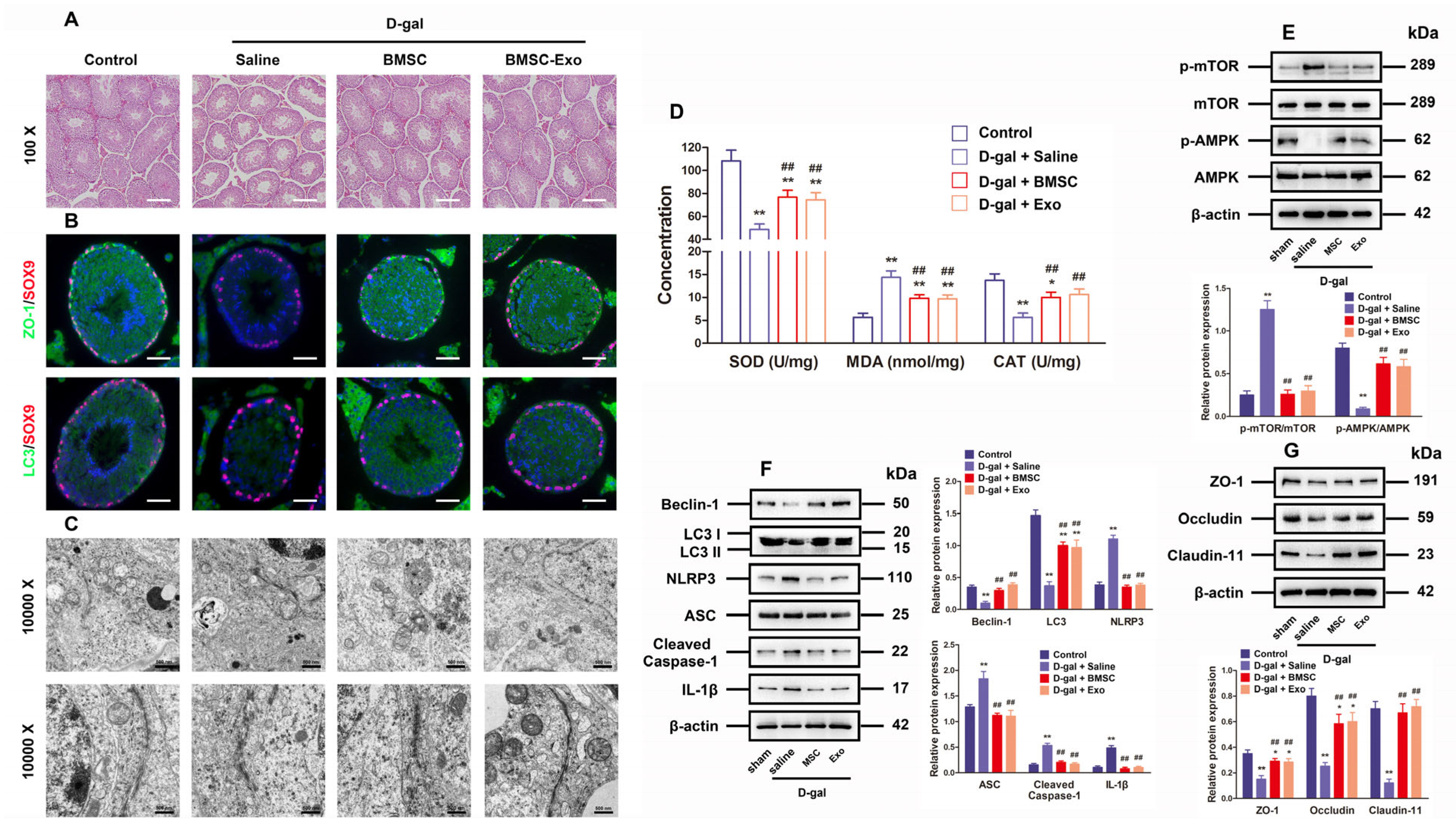
3.6. BMSCs and BMSC-Exos Improved Autophagy and BTB Function as Well as Inhibited ROS Generation and NLRP3 Inflammasome Activation in the Testes of D-Gal-Induced Aging Mouse Model
4. Discussion
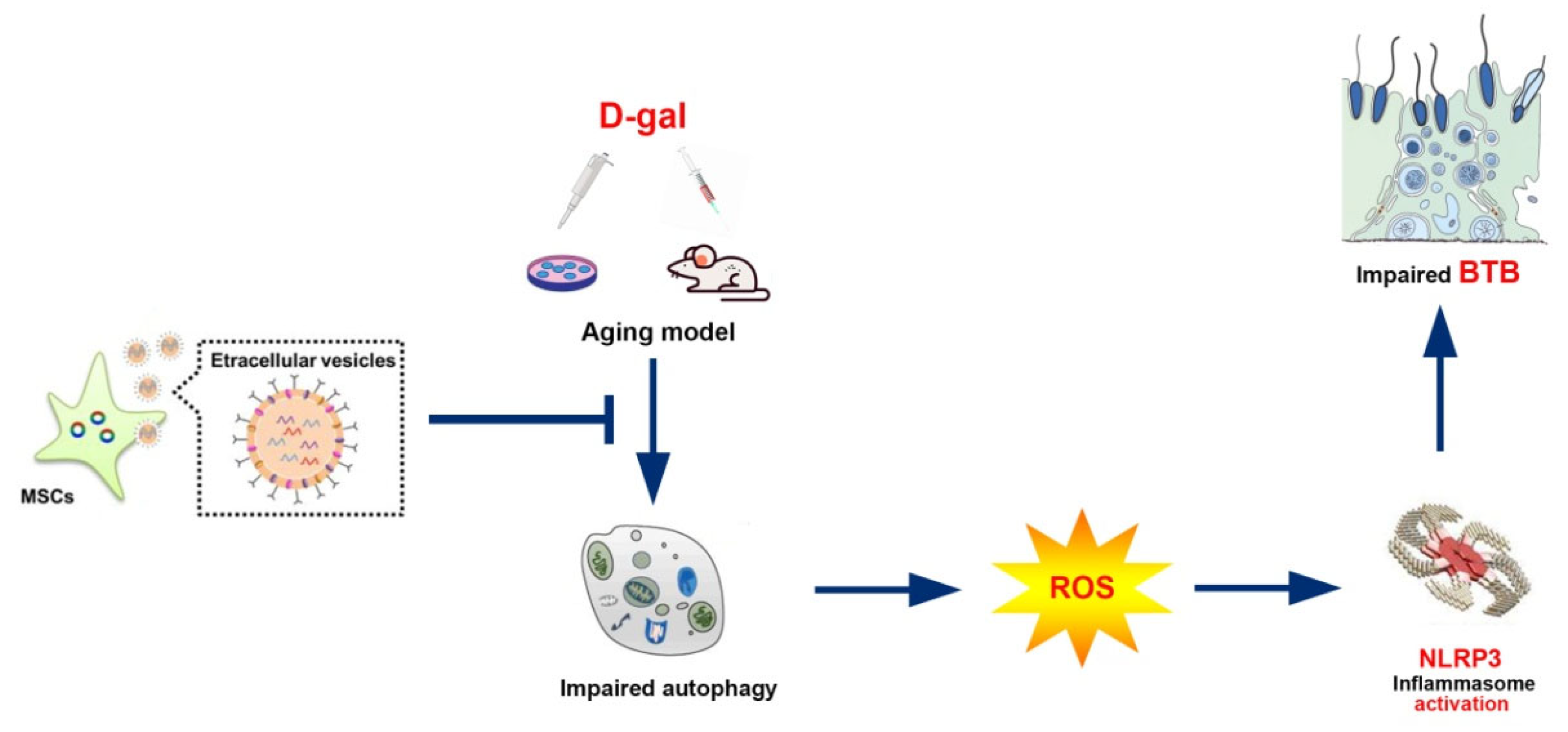
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Almeida, S.; Rato, L.; Sousa, M.; Alves, M.G.; Oliveira, P.F. Fertility and sperm quality in the aging Male. Curr. Pharm. Des. 2017, 23, 4429–4437. [Google Scholar] [CrossRef] [PubMed]
- Decaroli, M.C.; Rochira, V. Aging and sex hormones in males. Virulence 2016, 8, 545–570. [Google Scholar] [CrossRef] [PubMed]
- Paul, C.; Robaire, B. Impaired function of the blood-testis barrier during aging is preceded by a decline in cell adhesion proteins and GTPases. PLoS ONE 2013, 8, e84354. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Fang, Z.; Wang, Q.; Sheng, X.; Qi, X.; Xing, K.; Guo, Y.; Ni, H.; Wang, X.; Zhang, Y. Curcumin Ameliorates Age-Induced Tight Junction Impaired in Porcine Sertoli Cells by Inactivating the NLRP3 Inflammasome through the AMPK/SIRT3/SOD2/mtROS Signaling Pathway. Oxidative Med. Cell. Longev. 2023, 2023, 1708251. [Google Scholar] [CrossRef] [PubMed]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative stress and autophagy: The clash between damage and metabolic needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Yamamuro, T.; Nakamura, S.; Yamano, Y.; Endo, T.; Yanagawa, K.; Tokumura, A.; Matsumura, T.; Kobayashi, K.; Mori, H.; Enokidani, Y.; et al. Rubicon prevents autophagic degradation of GATA4 to promote Sertoli cell function. PLoS Genet. 2021, 17, e1009688. [Google Scholar] [CrossRef]
- Aman, Y.; Schmauck-Medina, T.; Hansen, M.; Morimoto, R.I.; Simon, A.K.; Bjedov, I.; Palikaras, K.; Simonsen, A.; Johansen, T.; Tavernarakis, N.; et al. Autophagy in healthy aging and disease. Nat. Aging 2021, 1, 634–650. [Google Scholar] [CrossRef]
- Daenen, K.; Andries, A.; Mekahli, D.; Van Schepdael, A.; Jouret, F.; Bammens, B. Oxidative stress in chronic kidney disease. Pediatr. Nephrol. 2019, 34, 975–991. [Google Scholar] [CrossRef]
- Dodson, M.; Darley-Usmar, V.; Zhang, J. Cellular metabolic and autophagic pathways: Traffic control by redox signaling. Free Radic. Biol. Med. 2013, 63, 207–221. [Google Scholar] [CrossRef]
- Kaarniranta, K.; Uusitalo, H.; Blasiak, J.; Felszeghy, S.; Kannan, R.; Kauppinen, A.; Salminen, A.; Sinha, D.; Ferrington, D. Mechanisms of mitochondrial dysfunction and their impact on age-related macular degeneration. Prog. Retin. Eye Res. 2020, 79, 100858. [Google Scholar] [CrossRef]
- Park, Y.J.; Pang, M.G. Mitochondrial Functionality in Male Fertility: From Spermatogenesis to Fertilization. Antioxidants 2021, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Asadi, A.; Ghahremani, R.; Abdolmaleki, A.; Rajaei, F. Role of sperm apoptosis and oxidative stress in male infertility: A narrative review. Int. J. Reprod. Biomed. 2021, 19, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Hu, H.; Yin, L.; Wang, L.; Luo, K.; Luo, N. Arachidonic acid impairs the function of the blood-testis barrier via triggering mitochondrial complex-ROS-P38 MAPK axis in hyperthermal Sertoli cells. Ecotoxicol. Environ. Saf. 2023, 252, 114598. [Google Scholar] [CrossRef] [PubMed]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef]
- Bai, B.; Yang, Y.; Wang, Q.; Li, M.; Tian, C.; Liu, Y.; Aung, L.H.H.; Li, P.F.; Yu, T.; Chu, X.M. NLRP3 inflammasome in endothelial dysfunction. Cell Death Dis. 2020, 11, 776. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Liu, C.; Wang, L.; Sui, C.; Zhang, H. Therapeutic Role of Mesenchymal Stem Cell-Derived Extracellular Vesicles in Female Reproductive Diseases. Front. Endocrinol. 2021, 12, 665645. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Roy, P.; Corbitt, C.; Kakar, S.S. Application of Stem Cell Therapy for Infertility. Cells 2021, 10, 1613. [Google Scholar] [CrossRef]
- Fraile, M.; Eiro, N.; Costa, L.A.; Martín, A.; Vizoso, F.J. Aging and Mesenchymal Stem Cells: Basic Concepts, Challenges and Strategies. Biology 2022, 11, 1678. [Google Scholar] [CrossRef]
- Wang, S.S.; Zhang, X.; Ke, Z.Z.; Wen, X.Y.; Li, W.D.; Liu, W.B.; Zhuang, X.D.; Liao, L.Z. D-galactose-induced cardiac ageing: A review of model establishment and potential interventions. J. Cell. Mol. Med. 2022, 26, 5335–5359. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, T.; Liu, S.; Chen, Y. Effects of bone marrow mesenchymal stem cells on ovarian and testicular function in aging Sprague-Dawley rats induced by D-galactose. Cell Cycle 2020, 19, 2340–2350. [Google Scholar] [CrossRef]
- Kim, Y.C.; Guan, K.L. mTOR: A pharmacologic target for autophagy regulation. J. Clin. Investig. 2015, 125, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, H.; Shang, Y.; Liu, W.; Song, Z.; Zhao, H.; Wang, L.; Jia, P.; Gao, F.; Xu, Z.; et al. Autophagy is required for ectoplasmic specialization assembly in sertoli cells. Autophagy 2016, 12, 814–832. [Google Scholar] [CrossRef]
- Huang, L.; Chen, Y.; Liu, R.; Li, B.; Fei, X.; Li, X.; Liu, G.; Li, Y.; Xu, B.; Fang, W. P-Glycoprotein Aggravates Blood Brain Barrier Dysfunction in Experimental Ischemic Stroke by Inhibiting Endothelial Autophagy. Aging Dis. 2022, 13, 1546–1561. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Yang, W.; Cui, W.; Li, C.; Ma, C.; Ji, X.; Hong, J.; Qu, Z.; Chen, J.; Liu, A.; et al. Therapeutic potential and mechanisms of mesenchymal stem cell-derived exosomes as bioactive materials in tendon-bone healing. J. Nanobiotechnol. 2023, 21, 14. [Google Scholar] [CrossRef]
- Pu, X.; Zhang, L.; Zhang, P.; Xu, Y.; Wang, J.; Zhao, X.; Dai, Z.; Zhou, H.; Zhao, S.; Fan, A. Human UC-MSC-derived exosomes facilitate ovarian renovation in rats with chemotherapy-induced premature ovarian insufficiency. Front. Endocrinol. 2023, 14, 1205901. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Börger, V.; Mohamud, Y.A.; Tertel, T.; Stambouli, O.; Murke, F.; Freund, N.; Kleinschnitz, C.; Herz, J.; Gunzer, M.; et al. Postischemic Neuroprotection Associated With Anti-Inflammatory Effects by Mesenchymal Stromal Cell-Derived Small Extracellular Vesicles in Aged Mice. Stroke 2022, 53, e14–e18. [Google Scholar] [CrossRef]
- Shin, J.Y.; Kim, D.Y.; Lee, J.; Shin, Y.J.; Kim, Y.S.; Lee, P.H. Priming mesenchymal stem cells with α-synuclein enhances neuroprotective properties through induction of autophagy in Parkinsonian models. Stem Cell Res. Ther. 2022, 13, 483. [Google Scholar] [CrossRef]
- Liu, L.; Jin, X.; Hu, C.F.; Li, R.; Zhou, Z.; Shen, C.X. Exosomes Derived from Mesenchymal Stem Cells Rescue Myocardial Ischaemia/Reperfusion Injury by Inducing Cardiomyocyte Autophagy Via AMPK and Akt Pathways. Cell Physiol. Biochem. 2017, 43, 52–68. [Google Scholar] [CrossRef]
- Bellavite, P. Neuroprotective Potentials of Flavonoids: Experimental Studies and Mechanisms of Action. Antioxidants 2023, 12, 280. [Google Scholar] [CrossRef]
- Yang, X.; Wang, R.; Zhou, H.; Wang, L.; Wang, R.; Li, H.; Tan, B.; Wu, Q.; Xu, X.; Cui, L.; et al. β-hydroxybutyrate Alleviates Learning and Memory Impairment Through the SIRT1 Pathway in D-Galactose-Injured Mice. Front. Pharmacol. 2021, 12, 751028. [Google Scholar] [CrossRef]
- Liao, L.Z.; Chen, Z.C.; Wang, S.S.; Liu, W.B.; Zhao, C.L.; Zhuang, X.D. NLRP3 inflammasome activation contributes to the pathogenesis of cardiocytes aging. Aging 2021, 13, 20534–20551. [Google Scholar] [CrossRef]
- Tan, Y.; Huang, Y.; Mei, R.; Mao, F.; Yang, D.; Liu, J.; Xu, W.; Qian, H.; Yan, Y. HucMSC-derived exosomes delivered BECN1 induces ferroptosis of hepatic stellate cells via regulating the xCT/GPX4 axis. Cell Death Dis. 2022, 13, 319. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, Y.; Du, Z.; Wu, T.; Yang, C. Hair follicle mesenchymal stem cell exosomal lncRNA H19 inhibited NLRP3 pyroptosis to promote diabetic mouse skin wound healing. Aging 2023, 15, 791–809. [Google Scholar] [CrossRef]
- Kaarniranta, K.; Blasiak, J.; Liton, P.; Boulton, M.; Klionsky, D.J.; Sinha, D. Autophagy in age-related macular degeneration. Autophagy 2023, 19, 388–400. [Google Scholar] [CrossRef]
- Biasizzo, M.; Kopitar-Jerala, N. Interplay Between NLRP3 Inflammasome and Autophagy. Front. Immunol. 2020, 11, 591803. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Malik, S.A.; Amir, M.; Kumar, P.; Cingolani, F.; Wen, J.; Liu, Y.; Zhao, E.; Farris, A.B.; Raeman, R.; et al. Decreased Hepatocyte Autophagy Leads to Synergistic IL-1β and TNF Mouse Liver Injury and Inflammation. Hepatology 2020, 72, 595–608. [Google Scholar] [CrossRef]
- Wu, J.; Yang, Y.; Liu, L.; Zhu, W.; Liu, M.; Yu, X.; Li, W. ROS-AMPK/mTOR-dependent enterocyte autophagy is involved in the regulation of Giardia infection-related tight junction protein and nitric oxide levels. Front. Immunol. 2023, 14, 1120996. [Google Scholar] [CrossRef]
- Su, Y.; Zhang, W.; Zhang, R.; Yuan, Q.; Wu, R.; Liu, X.; Wuri, J.; Li, R.; Yan, T. Activation of Cholinergic Anti-Inflammatory Pathway Ameliorates Cerebral and Cardiac Dysfunction After Intracerebral Hemorrhage Through Autophagy. Front. Immunol. 2022, 13, 870174. [Google Scholar] [CrossRef] [PubMed]
- Keshtkar, S.; Azarpira, N.; Ghahremani, M.H. Mesenchymal stem cell-derived extracellular vesicles: Novel frontiers in regenerative medicine. Stem Cell Res. Ther. 2018, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Qiu, G.; Zheng, G.; Ge, M.; Wang, J.; Huang, R.; Shu, Q.; Xu, J. Functional proteins of mesenchymal stem cell-derived extracellular vesicles. Stem Cell Res. Ther. 2019, 10, 359. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Yan, J.; Qiao, L.; Zeng, J.; Cao, F.; Sheng, X.; Qi, X.; Long, C.; Liu, B.; Wang, X.; et al. Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Ameliorate Aging-Induced BTB Impairment in Porcine Testes by Activating Autophagy and Inhibiting ROS/NLRP3 Inflammasomes via the AMPK/mTOR Signaling Pathway. Antioxidants 2024, 13, 183. https://doi.org/10.3390/antiox13020183
Zhou Y, Yan J, Qiao L, Zeng J, Cao F, Sheng X, Qi X, Long C, Liu B, Wang X, et al. Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Ameliorate Aging-Induced BTB Impairment in Porcine Testes by Activating Autophagy and Inhibiting ROS/NLRP3 Inflammasomes via the AMPK/mTOR Signaling Pathway. Antioxidants. 2024; 13(2):183. https://doi.org/10.3390/antiox13020183
Chicago/Turabian StyleZhou, Yi, Jiale Yan, Limin Qiao, Jiaqin Zeng, Fuyu Cao, Xihui Sheng, Xiaolong Qi, Cheng Long, Bingying Liu, Xiangguo Wang, and et al. 2024. "Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Ameliorate Aging-Induced BTB Impairment in Porcine Testes by Activating Autophagy and Inhibiting ROS/NLRP3 Inflammasomes via the AMPK/mTOR Signaling Pathway" Antioxidants 13, no. 2: 183. https://doi.org/10.3390/antiox13020183
APA StyleZhou, Y., Yan, J., Qiao, L., Zeng, J., Cao, F., Sheng, X., Qi, X., Long, C., Liu, B., Wang, X., Yao, H., & Xiao, L. (2024). Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Ameliorate Aging-Induced BTB Impairment in Porcine Testes by Activating Autophagy and Inhibiting ROS/NLRP3 Inflammasomes via the AMPK/mTOR Signaling Pathway. Antioxidants, 13(2), 183. https://doi.org/10.3390/antiox13020183