
The Antioxidant Mechanism of Peptides Extracted from Tuna Protein Revealed Using a Molecular Docking Simulation

Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Preparation of Polypeptide from Tuna
2.3. Peptide Sequence Identification with Q-TOF MS/MS
2.4. Overall Activity Prediction of TPP
2.5. Antioxidative Activity and Toxicity Prediction of Effective Peptides
2.6. Antioxidant Active Peptide Comparison
2.7. Validation of Antioxidant Activity
2.7.1. Determination of DPPH Free Radical Scavenging Rate
2.7.2. Determination of Hydroxyl Radical Scavenging Rate
2.7.3. Determination of Superoxide Anion Scavenging Rate
2.8. Cell Culture and Antioxidant Test
2.9. Molecular Docking
2.10. Statistical Analyses
3. Results and Discussion
3.1. Results of Enzymatic Hydrolysis of TPP
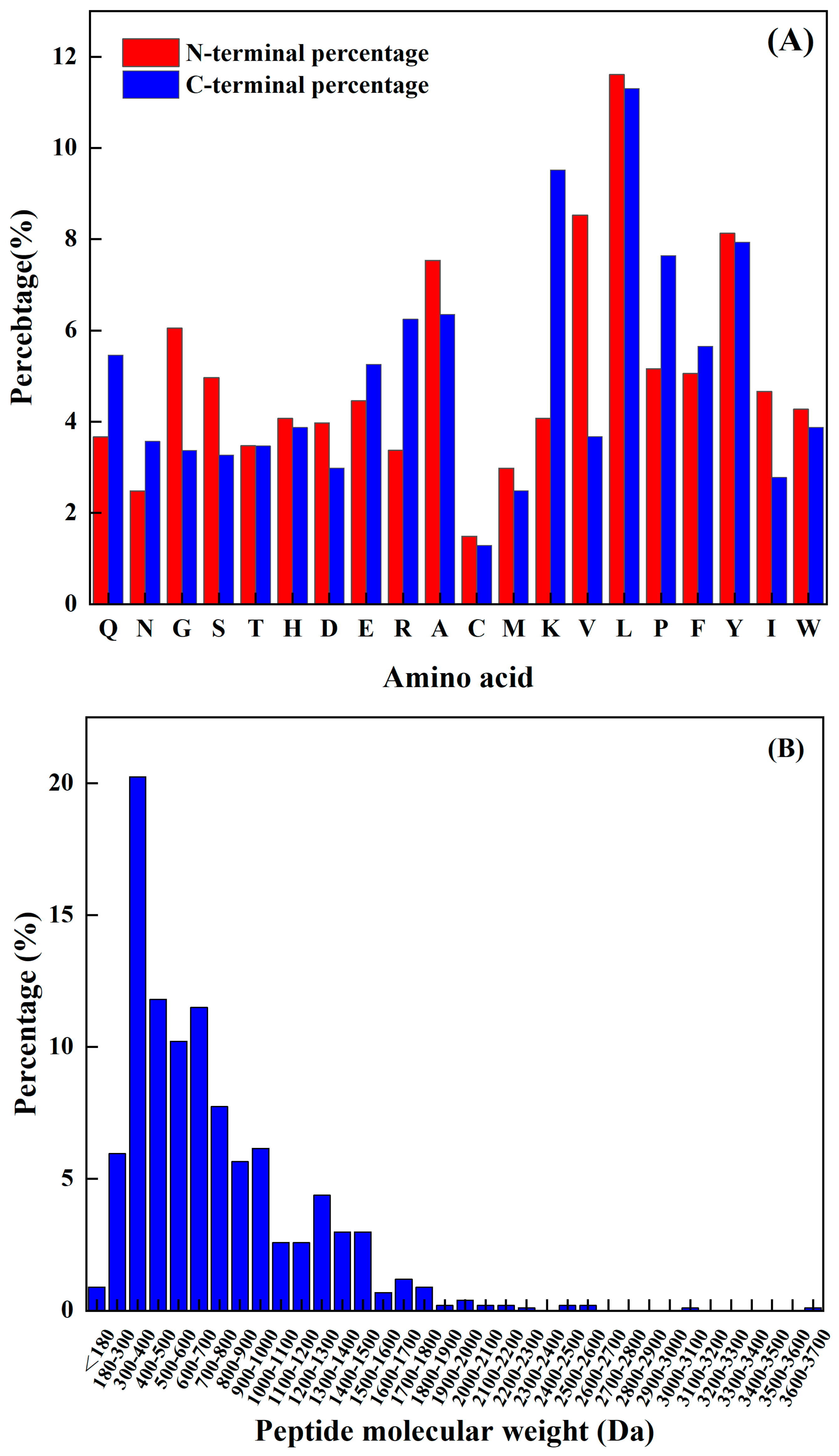
3.2. Atlas Characterization Analysis of TPP
3.3. Prediction of Overall Peptides Activity and Toxicity
3.4. Prediction of Antioxidant Activity of Overall Functionally Active Peptides
3.5. Structural Comparison of Antioxidant Active Peptides
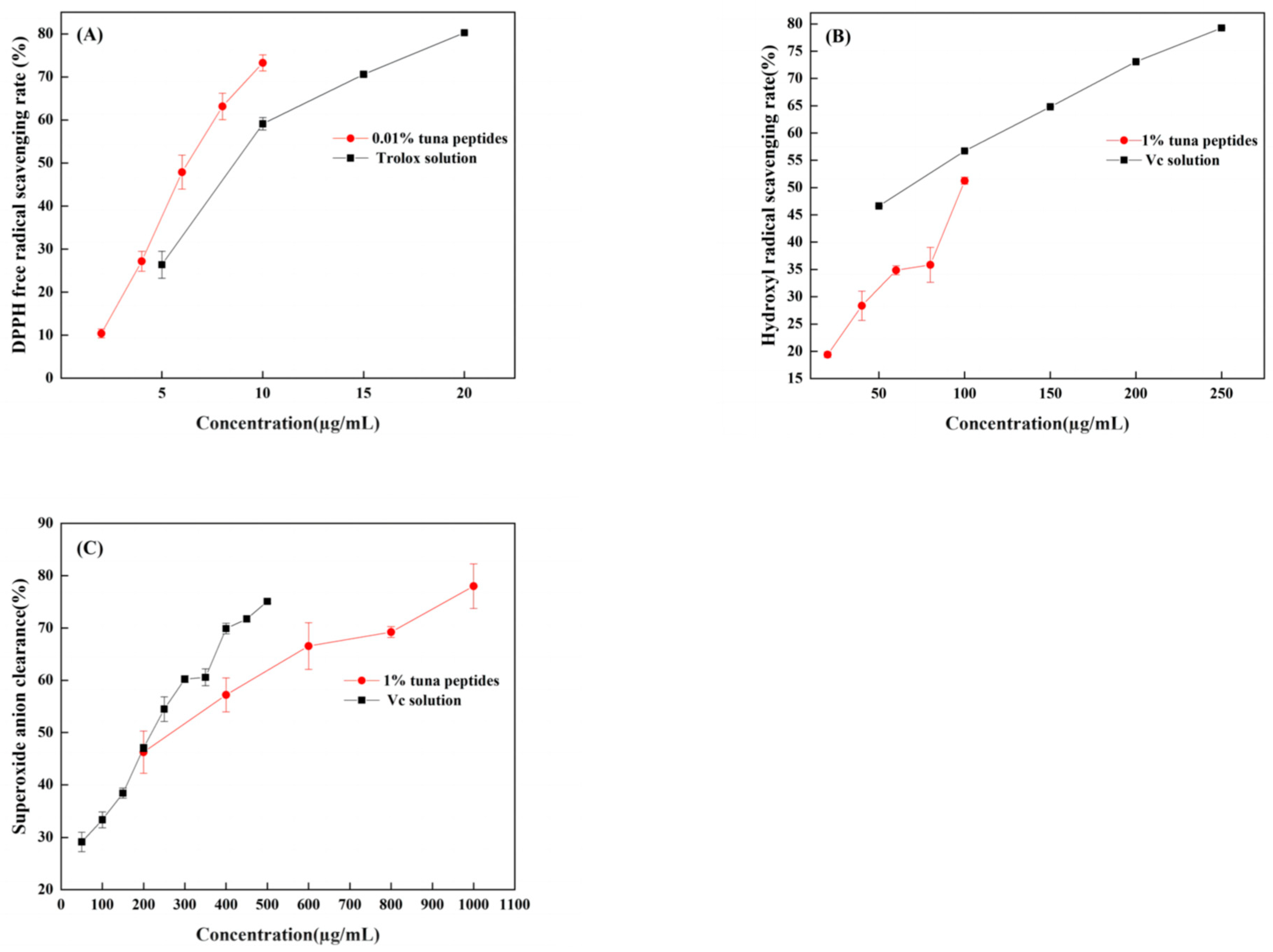
3.6. Chemical Verification of Antioxidant Activity of Peptides
3.7. Effects of Active Peptides on Cellular Antioxidant Enzymes
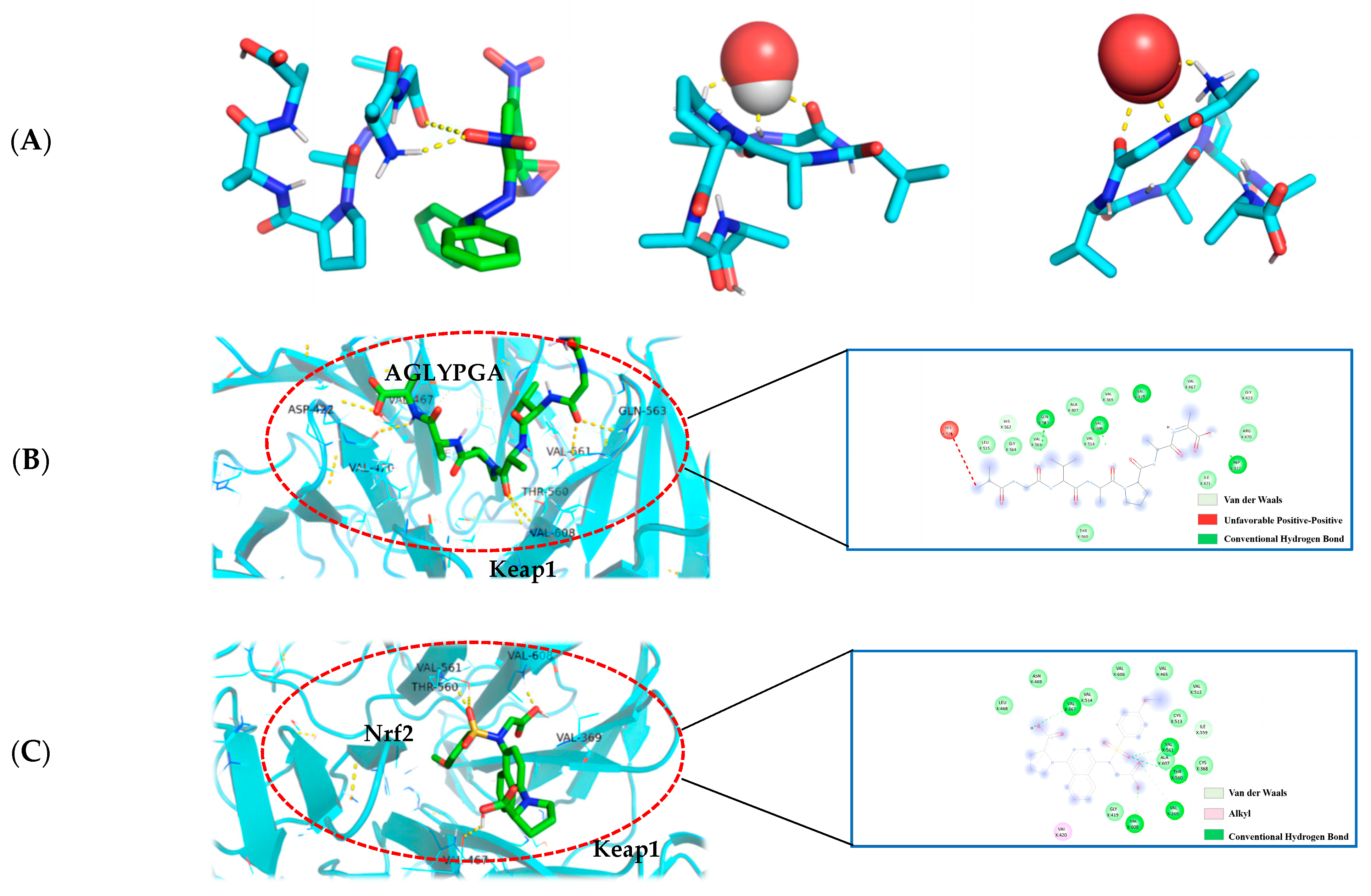
3.8. Optimal Docking Model of Peptides and DPPH, Hydroxyl Radical, Superoxide Anion, and Keap1
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guerard, F.; Dufosse, L.; Broise, D. Enzymatic hydrolysis of proteins from yellowfin tuna (Thunnus albacares) wastes using Alcalase. J. Mol. Catal. B Enzym. 2001, 11, 1051–1059. [Google Scholar] [CrossRef]
- Ren, J.Y.; Wang, H.Y.; Zhao, M.M. Enzymatic hydrolysis of grass carp myofibrillar protein and antioxidant properties of hydrolysate. Czech J. Food Sci. 2010, 28, 475–484. [Google Scholar] [CrossRef]
- Jae, Y.J.; Zhong, J.Q.; Se, K.K. Purification and characterization of an antioxidant peptide obtained from tuna backbone protein by enzymatic hydrolysis. Process Biochem. 2007, 42, 840–846. [Google Scholar] [CrossRef]
- Hsu, K.C. Purification of antioxidative peptides prepared from enzymatic hydrolysate of tuna dark muscle by-product. Food Chem. 2010, 122, 42–48. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Duithie, S.J.; Jenkinson, A.M.; Crozier, A. The effects of cranberry juice consumption on antioxidant status and biomarkers relating to heart disease and cancer in healthy human volunteers. Eur. J. Nutr. 2006, 45, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Girgih, A.T.; He, R.; Malomo, S. Structural and functional characterization of hemp seed (Cannabis sativa L.) protein-derived antioxidant and antihypertensive peptides. J. Funct. Foods 2014, 6, 384–394. [Google Scholar] [CrossRef]
- Luca, P.Z.; Giulio, R. Molecular Docking: Shifting Paradigms in Drug Discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef]
- Kitchen, D.B.; Decornez, H.; Furr, J.R.; Bajorath, J. Docking and scoring in virtual screening for drug discovery: Methods and applications. Nat. Rev. Drug Discov. 2004, 3, 935–949. [Google Scholar] [CrossRef]
- Jo, C.; Khan, F.F.; Khan, M.I.; Iqbal, J. Marine bioactive peptides: Types, structures, and physiological functions. Food Rev. Int. 2017, 33, 44–61. [Google Scholar] [CrossRef]
- Liu, J.; Wang, C.; Wang, Z.; Zhang, C.; Lu, S.; Liu, J. The antioxidant and free radical scavenging activities of extract and fractions from corn silk (Zea mays L.) and related flavone glycosides. Food Chem. 2011, 126, 261–269. [Google Scholar] [CrossRef]
- Cao, Y.; Chu, Q.; Ye, J. Determination of hydroxyl radical by capillary electrophoresis and studies on hydroxyl radical scavenging activities of Chinese herbs. Anal. Bioanal. Chem. 2003, 376, 691–695. [Google Scholar] [CrossRef]
- Misak, A.; Grman, M.; Bacova, Z.; Rezuchova, I.; Hudecova, S.; Ondriasova, E. Polysulfides and products of H2S/S-nitrosoglutathione in comparison to H2S, glutathione and antioxidant Trolox are potent scavengers of superoxide anion radical and produce hydroxyl radical by decomposition of H2O2. Nitric Oxide 2018, 76, 136–151. [Google Scholar] [CrossRef]
- Vera, M.D.; Joulli, M.M. Natural products as probes of cell biology: 20 years of didemnin research. Med. Res. Rev. 2002, 22, 102–145. [Google Scholar] [CrossRef]
- Kim, S.Y.; Je, J.Y.; Kim, S.K. Purification and characterization of antioxidant peptide from hoki (Johnius belengerii) frame protein by gastrointestinal digestion. J. Nutr. Biochem. 2007, 18, 31–38. [Google Scholar] [CrossRef]
- Lapsongphon, N.; Yongsawatdigul, J. Production and purification of antioxidant peptides from a mungbean meal hydrolysate by Virgibacillus sp. sk37 proteinase. Food Chem. 2013, 141, 992–999. [Google Scholar] [CrossRef]
- Niranjan, R.; Eresha, M.; Won, K.J.; Jae, Y.J.; Se, K.K. Purification of a radical scavenging peptide from fermented mussel sauce and its antioxidant properties. Food Res. Int. 2005, 38, 175–182. [Google Scholar] [CrossRef]
- Shi, C.; Liu, M.; Zhao, H.; Lv, Z.; Liang, L.; Zhang, B. A Novel Insight into Screening for Antioxidant Peptides from Hazelnut Protein: Based on the Properties of Amino Acid Residues. Antioxidants 2021, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Huang, J.; Huan, R.; Chen, L.; Yi, C.; Liu, D.; Wang, M.; Liu, C.; He, H. New insights into the structure-activity relationships of antioxidative peptide PMRGGGGYHY. Food Chem. 2020, 1, 337. [Google Scholar] [CrossRef] [PubMed]
- Hang, J.; Li, M.; Zhang, G.; Tian, Y.U.; Kong, F.; Xiong, S. Identification of novel antioxidant peptides from snakehead (Channa argus) soup generated during gastrointestinal digestion and insights into the anti-oxidation mechanisms. Food Chem. 2021, 337, 127921. [Google Scholar] [CrossRef]
- Didik, H.U.; Nashi, W. Identifications small molecules inhibitor of p53-mortalin complex for cancer drug using virtual screening. Bioinformation 2021, 89, 426–429. [Google Scholar] [CrossRef]
- Kensler, T.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.T.; Huang, R.; Sedykh, A.; Wang, W.; Xia, M.; Zhu, H. Mechanism profiling of hepatotoxicity caused by oxidative stress using antioxidant response element reporter gene assay models and big data. Environ. Health Perspect. 2016, 124, 634–641. [Google Scholar] [CrossRef]
- Wang, N.; Wang, W.; Sadiq, F.A.; Wang, S.; Liu, C.; Jin, J. Involvement of Nrf2 and Keap1 in the activation of antioxidant responsive element (ARE) by chemopreventive agent peptides from soft-shelled turtle. Process Biochem. 2020, 92, 174–181. [Google Scholar] [CrossRef]
- Yin, J.; Han, Y.; Liu, M. Structure-guided discovery of antioxidant peptides bounded to the Keap1 receptor as hunter for potential dietary antioxidants. Food Chem. 2022, 373, 130999. [Google Scholar] [CrossRef]
- Tonolo, F.; Folda, A.; Cesaro, L.; Scalcon, V.; Marin, O.; Ferro, S.; Rigobello, M.P. Milk-derived bioactive peptides exhibit antioxidant activity through the Keap1-Nrf2 signaling pathway. J. Funct. Foods 2020, 64, 103696. [Google Scholar] [CrossRef]
- Han, J.; Huang, Z.; Tang, S.; Lu, C.; Wan, H.; Zhou, J.; Su, X. The novel peptides ICRD and LCGEC screened from tuna roe show antioxidative activity via Keap1/Nrf2-ARE pathway regulation and gut microbiota modulation. Food Chem. 2020, 327, 127094. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, J.; Nie, S.; Ding, L.; Wang, L.; Liu, J. Direct inhibition of Keap1–Nrf2 interaction by egg-derived peptides DKK and DDW revealed by molecular docking and fluorescence polarization. RSC Adv. 2017, 7, 34963–34971. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Sequence | Molecular Weight | Activity Score | Peptide Sequence | Molecular Weight | Activity Score |
|---|---|---|---|---|---|
| FDTFLPM | 869.3993 | 0.918121 | AEPAPAPAPAPEPAPAPA | 1620.8147 | 0.614793 |
| PGGVPFLP | 814.4225 | 0.895559 | DLGGGTF | 665.302 | 0.612586 |
| PAPAPAPAAGGT | 982.5059 | 0.844022 | AGPPGSTGP | 739.35 | 0.601717 |
| ASGPINF | 704.3493 | 0.842426 | GPSGPAGPTA | 739.35 | 0.596112 |
| APAPAPAPAAG | 911.4476 | 0.803473 | AAGPPGSTGP | 810.3871 | 0.593769 |
| APVAPGYGGG | 876.3977 | 0.802741 | LPGGGPVL | 707.433 | 0.586057 |
| GPAGASGPAGPR | 993.4991 | 0.782288 | KAEPAPAPAPAPEPAPAPA | 1748.9097 | 0.585361 |
| APAPAPAPAPAPA | 1168.624 | 0.767656 | VILPVPAF | 854.5266 | 0.582872 |
| AGPSGPR | 640.3292 | 0.765178 | SKTPGLM | 746.3997 | 0.582128 |
| APAPAPAPAPA | 929.497 | 0.762092 | PFDQDDWE | 1050.3931 | 0.579299 |
| APAPAPAPA | 761.4071 | 0.760005 | HVDPDNF | 842.3558 | 0.574675 |
| AAPAPAPAPAPAPA | 1168.624 | 0.75938 | KKAEPAPAPAPAPEPAPAPA | 1877.0046 | 0.570387 |
| SGPINFT | 734.3599 | 0.753769 | AAIIKPL | 724.4847 | 0.565087 |
| ASGPINFT | 805.397 | 0.751915 | AEPAPAPAPAPEPAPAPAAPA | 1859.9417 | 0.563469 |
| APAPAPAAG | 743.3452 | 0.748276 | ADFDAVL | 791.3701 | 0.556689 |
| IDFGMDL | 809.3629 | 0.746159 | AKKAEPAPAPAPAPEPAPAP | 1877.0046 | 0.555846 |
| AAAPAPAPAPAPAPA | 1239.6611 | 0.730839 | GRGGAGGL | 643.3401 | 0.548597 |
| GPSGPAG | 541.2496 | 0.728095 | EASGPINFT | 934.4396 | 0.546921 |
| APGKGIL | 654.4064 | 0.679452 | PGLSGAPG | 654.3337 | 0.542914 |
| SGPAGPA | 555.2653 | 0.667579 | DASLPGNYG | 892.3926 | 0.539879 |
| PGPSGPA | 581.2809 | 0.660942 | GERGAPGIGGP | 966.4882 | 0.539457 |
| AGAPGPSGP | 709.3395 | 0.641999 | GEPGPAG | 583.2601 | 0.521885 |
| AFPPDVA | 715.3541 | 0.641548 | GPETGPRGAPGPA | 1061.5254 | 0.519873 |
| AGLYPGA | 675.3591 | 0.640776 | GESGNPGAPG | 841.3566 | 0.519609 |
| EASGPINF | 833.3919 | 0.629953 | VAPGKGIL | 753.4749 | 0.511749 |
| GNHAAIIKPL | 1033.592 | 0.625024 | AGAGPGA | 541.2496 | 0.510304 |
| APGAPGA | 539.2703 | 0.619753 | GGVPGGAPLAV | 893.497 | 0.505766 |
| Peptide Sequence | Peptide Length | Antioxidant Activity Score | Toxicity |
|---|---|---|---|
| AGLYPGA | 7 | 0.54491031 | no |
| GEPGPAG | 7 | 0.50003815 | no |
| LPGGGPVL | 8 | 0.53823709 | no |
| APVAPGYGGG | 10 | 0.53759086 | no |
| AAAPAPAPAPAPAPA | 15 | 0.53961134 | no |
| AEPAPAPAPAPEPAPAPA | 18 | 0.54290658 | no |
| AEPAPAPAPAPEPAPAPAAPA | 21 | 0.53858268 | no |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Liu, J.; Wang, C.; Wen, Z.; Zheng, B. The Antioxidant Mechanism of Peptides Extracted from Tuna Protein Revealed Using a Molecular Docking Simulation. Antioxidants 2024, 13, 166. https://doi.org/10.3390/antiox13020166
Guo X, Liu J, Wang C, Wen Z, Zheng B. The Antioxidant Mechanism of Peptides Extracted from Tuna Protein Revealed Using a Molecular Docking Simulation. Antioxidants. 2024; 13(2):166. https://doi.org/10.3390/antiox13020166
Chicago/Turabian StyleGuo, Xiaojun, Jiaxin Liu, Cheng Wang, Zhengshun Wen, and Bin Zheng. 2024. "The Antioxidant Mechanism of Peptides Extracted from Tuna Protein Revealed Using a Molecular Docking Simulation" Antioxidants 13, no. 2: 166. https://doi.org/10.3390/antiox13020166
APA StyleGuo, X., Liu, J., Wang, C., Wen, Z., & Zheng, B. (2024). The Antioxidant Mechanism of Peptides Extracted from Tuna Protein Revealed Using a Molecular Docking Simulation. Antioxidants, 13(2), 166. https://doi.org/10.3390/antiox13020166