
Sulforaphane—A Compound with Potential Health Benefits for Disease Prevention and Treatment: Insights from Pharmacological and Toxicological Experimental Studies

 ,
,  ,
,  , , and
, , and
Abstract
1. Introduction
2. SFN—General Information
3. Mechanisms of SFN-Linked Beneficial Effects
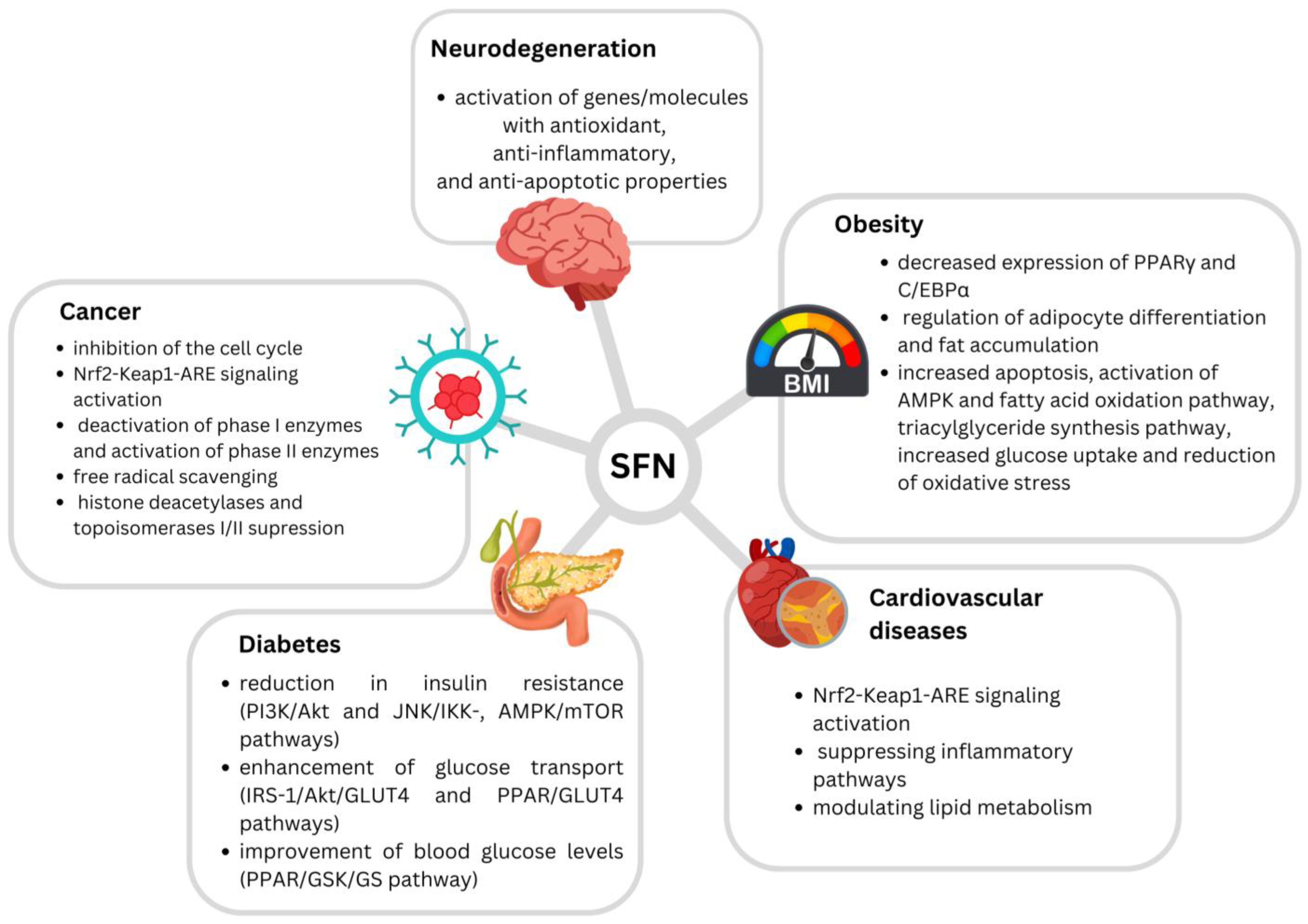
4. The Potential of SFN in Disease Prevention and Treatment
4.1. Anticancerogenic Effects
4.2. Antidiabetic/Anti-Obesogenic Effects
4.3. Cardiovascular-Protective Effects
4.4. Neuroprotective Effects
5. SFN’s Protective Effects against Toxic Substances
6. Toxic Effects
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Ng, R.; Sutradhar, R.; Yao, Z.; Wodchis, W.P.; Rosella, L.C. Smoking, Drinking, Diet and Physical Activity—Modifiable Lifestyle Risk Factors and Their Associations with Age to First Chronic Disease. Int. J. Epidemiol. 2020, 49, 113–130. [Google Scholar] [CrossRef] [PubMed]
- Barouki, R.; Gluckman, P.D.; Grandjean, P.; Hanson, M.; Heindel, J.J. Developmental Origins of Non-Communicable Disease: Implications for Research and Public Health. Environ. Health 2012, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Mishra, V.K.; Srivastava, S.; Muhammad, T.; Murthy, P.V. Relationship between Tobacco Use, Alcohol Consumption and Non-Communicable Diseases among Women in India: Evidence from National Family Health Survey-2015-16. BMC Public Health 2022, 22, 713. [Google Scholar] [CrossRef] [PubMed]
- Iahtisham-Ul-Haq; Khan, S.; Awan, K.A.; Iqbal, M.J. Sulforaphane as a Potential Remedy against Cancer: Comprehensive Mechanistic Review. J. Food Biochem. 2022, 46, e13886. [Google Scholar] [CrossRef] [PubMed]
- Aune, D.; Giovannucci, E.; Boffetta, P.; Fadnes, L.T.; Keum, N.; Norat, T.; Greenwood, D.C.; Riboli, E.; Vatten, L.J.; Tonstad, S. Fruit and Vegetable Intake and the Risk of Cardiovascular Disease, Total Cancer and All-Cause Mortality—A Systematic Review and Dose-Response Meta-Analysis of Prospective Studies. Int. J. Epidemiol. 2017, 46, 1029–1056. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Li, M.; Yin, R. Phytochemical Content, Health Benefits, and Toxicology of Common Edible Flowers: A Review (2000–2015). Crit. Rev. Food Sci. Nutr. 2016, 56, S130–S148. [Google Scholar] [CrossRef]
- Kaiser, A.E.; Baniasadi, M.; Giansiracusa, D.; Giansiracusa, M.; Garcia, M.; Fryda, Z.; Wong, T.L.; Bishayee, A. Sulforaphane: A Broccoli Bioactive Phytocompound with Cancer Preventive Potential. Cancers 2021, 13, 4796. [Google Scholar] [CrossRef]
- Xie, X.; Chang, X.; Chen, L.; Huang, K.; Huang, J.; Wang, S.; Shen, X.; Liu, P.; Huang, H. Berberine Ameliorates Experimental Diabetes-Induced Renal Inflammation and Fibronectin by Inhibiting the Activation of RhoA/ROCK Signaling. Mol. Cell Endocrinol. 2013, 381, 56–65. [Google Scholar] [CrossRef]
- Novelle, M.G.; Wahl, D.; Diéguez, C.; Bernier, M.; de Cabo, R. Resveratrol Supplementation: Where Are We Now and Where Should We Go? Ageing Res. Rev. 2015, 21, 1–15. [Google Scholar] [CrossRef]
- Shrimali, D.; Shanmugam, M.K.; Kumar, A.P.; Zhang, J.; Tan, B.K.H.; Ahn, K.S.; Sethi, G. Targeted Abrogation of Diverse Signal Transduction Cascades by Emodin for the Treatment of Inflammatory Disorders and Cancer. Cancer Lett. 2013, 341, 139–149. [Google Scholar] [CrossRef]
- Hu, C.; Huang, Z.; Sun, B.; Liu, M.; Tang, L.; Chen, L. Metabolomic Profiles in Zebrafish Larvae Following Probiotic and Perfluorobutanesulfonate Coexposure. Environ. Res. 2022, 204, 112380. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Qi, Y.; Zhao, J.; Jiang, H.; Chen, X.; Ren, J. Synthesis and Biological Evaluation of Sulforaphane Derivatives as Potential Antitumor Agents. Eur. J. Med. Chem. 2013, 64, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Bozic, D.; Baralić, K.; Živančević, K.; Miljaković, E.A.; Ćurčić, M.; Antonijević, B.; Djordjević, A.B.; Bulat, Z.; Zhang, Y.; Yang, L.; et al. Predicting Sulforaphane-Induced Adverse Effects in Colon Cancer Patients via in Silico Investigation. Biomed. Pharmacother. 2022, 146, 112598. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Mao, X.; Du, M. Metabolism, Absorption, and Anti-Cancer Effects of Sulforaphane: An Update. Crit. Rev. Food Sci. Nutr. 2022, 62, 3437–3452. [Google Scholar] [CrossRef] [PubMed]
- Rafiei, H.; Ashrafizadeh, M.; Ahmadi, Z. MicroRNAs as Novel Targets of Sulforaphane in Cancer Therapy: The Beginning of a New Tale? Phyther. Res. 2020, 34, 721–728. [Google Scholar] [CrossRef]
- Liang, J.; Hänsch, G.M.; Hübner, K.; Samstag, Y. Sulforaphane as Anticancer Agent: A Double-Edged Sword? Tricky Balance between Effects on Tumor Cells and Immune Cells. Adv. Biol. Regul. 2019, 71, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Sestili, P.; Fimognari, C. Cytotoxic and Antitumor Activity of Sulforaphane: The Role of Reactive Oxygen Species. Biomed. Res. Int. 2015, 2015, 402386. [Google Scholar] [CrossRef]
- Bansal, M.; Singh, N.; Pal, S.; Dev, I.; Ansari, K.M. Chemopreventive Role of Dietary Phytochemicals in Colorectal Cancer. Adv. Mol. Toxicol. 2018, 12, 69–121. [Google Scholar]
- Yagishita, Y.; Fahey, J.W.; Dinkova-Kostova, A.T.; Kensler, T.W. Broccoli or Sulforaphane: Is It the Source or Dose That Matters? Molecules 2019, 24, 3593. [Google Scholar] [CrossRef]
- Houghton, C.A.; Fassett, R.G.; Coombes, J.S. Sulforaphane and Other Nutrigenomic Nrf2 Activators: Can the Clinician’s Expectation Be Matched by the Reality? Oxid. Med. Cell Longev. 2016, 2016, 7857186. [Google Scholar] [CrossRef]
- Houghton, C.A. Sulforaphane: Its “Coming of Age” as a Clinically Relevant Nutraceutical in the Prevention and Treatment of Chronic Disease. Oxid. Med. Cell Longev. 2019, 2019, 2716870. [Google Scholar] [CrossRef]
- Su, X.; Jiang, X.; Meng, L.; Dong, X.; Shen, Y.; Xin, Y. Anticancer Activity of Sulforaphane: The Epigenetic Mechanisms and the Nrf2 Signaling Pathway. Oxid. Med. Cell Longev. 2018, 2018, 5438179. [Google Scholar] [CrossRef]
- Kamal, M.M.; Akter, S.; Lin, C.-N.; Nazzal, S. Sulforaphane as an Anticancer Molecule: Mechanisms of Action, Synergistic Effects, Enhancement of Drug Safety, and Delivery Systems. Arch. Pharm. Res. 2020, 43, 371–384. [Google Scholar] [CrossRef]
- Janczewski, Ł. Sulforaphane and Its Bifunctional Analogs: Synthesis and Biological Activity. Molecules 2022, 27, 1750. [Google Scholar] [CrossRef]
- Mangla, B.; Javed, S.; Sultan, M.H.; Kumar, P.; Kohli, K.; Najmi, A.; Alhazmi, H.A.; Al Bratty, M.; Ahsan, W. Sulforaphane: A Review of Its Therapeutic Potentials, Advances in Its Nanodelivery, Recent Patents, and Clinical Trials. Phyther. Res. 2021, 35, 5440–5458. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, X.; Zhao, S.; Ma, C.; Cui, J.; Zheng, Y. Sulforaphane Protects against Cardiovascular Disease via Nrf2 Activation. Oxid. Med. Cell Longev. 2015, 2015, 407580. [Google Scholar] [CrossRef]
- Russo, M.; Spagnuolo, C.; Russo, G.L.; Skalicka-Woźniak, K.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.F.; Nabavi, S.M. Nrf2 Targeting by Sulforaphane: A Potential Therapy for Cancer Treatment. Crit. Rev. Food Sci. Nutr. 2018, 58, 1391–1405. [Google Scholar] [CrossRef]
- Babu, P.V.A.; Petersen, C.; Jia, Z. Sulforaphane and Atherosclerosis. In Glucosinolates; Springer: Cham, Switzerland, 2017; pp. 319–337. [Google Scholar]
- Martins, T.; Colaço, B.; Venâncio, C.; Pires, M.J.; Oliveira, P.A.; Rosa, E.; Antunes, L.M. Potential Effects of Sulforaphane to Fight Obesity. J. Sci. Food Agric. 2018, 98, 2837–2844. [Google Scholar] [CrossRef]
- Çakır, I.; Lining Pan, P.; Hadley, C.K.; El-Gamal, A.; Fadel, A.; Elsayegh, D.; Mohamed, O.; Rizk, N.M.; Ghamari-Langroudi, M. Sulforaphane Reduces Obesity by Reversing Leptin Resistance. Elife 2022, 11, e67368. [Google Scholar] [CrossRef]
- Masuda, M.; Yoshida-Shimizu, R.; Mori, Y.; Ohnishi, K.; Adachi, Y.; Sakai, M.; Kabutoya, S.; Ohminami, H.; Yamanaka-Okumura, H.; Yamamoto, H.; et al. Sulforaphane Induces Lipophagy through the Activation of AMPK-MTOR-ULK1 Pathway Signaling in Adipocytes. J. Nutr. Biochem. 2022, 106, 109017. [Google Scholar] [CrossRef]
- Wang, M.; Chen, M.; Guo, R.; Ding, Y.; Zhang, H.; He, Y. The Improvement of Sulforaphane in Type 2 Diabetes Mellitus (T2DM) and Related Complications: A Review. Trends Food Sci. Technol. 2022, 129, 397–407. [Google Scholar] [CrossRef]
- Schepici, G.; Bramanti, P.; Mazzon, E. Efficacy of Sulforaphane in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 8637. [Google Scholar] [CrossRef]
- Mahn, A.; Castillo, A. Potential of Sulforaphane as a Natural Immune System Enhancer: A Review. Molecules 2021, 26, 752. [Google Scholar] [CrossRef]
- Georgikou, C.; Yin, L.; Gladkich, J.; Xiao, X.; Sticht, C.; de la Torre, C.; Gretz, N.; Gross, W.; Schäfer, M.; Karakhanova, S.; et al. Inhibition of MiR30a-3p by Sulforaphane Enhances Gap Junction Intercellular Communication in Pancreatic Cancer. Cancer Lett. 2020, 469, 238–245. [Google Scholar] [CrossRef]
- Ishiura, Y.; Ishimaru, H.; Watanabe, T.; Fujimuro, M. Sulforaphane Exhibits Cytotoxic Effects against Primary Effusion Lymphoma Cells by Suppressing P38MAPK and AKT Phosphorylation. Biol. Pharm. Bull. 2019, 42, 2109–2112. [Google Scholar] [CrossRef]
- Xu, Y.; Han, X.; Li, Y.; Min, H.; Zhao, X.; Zhang, Y.; Qi, Y.; Shi, J.; Qi, S.; Bao, Y.; et al. Sulforaphane Mediates Glutathione Depletion via Polymeric Nanoparticles to Restore Cisplatin Chemosensitivity. ACS Nano 2019, 13, 13445–13455. [Google Scholar] [CrossRef]
- Lin, L.-C.; Yeh, C.-T.; Kuo, C.-C.; Lee, C.-M.; Yen, G.-C.; Wang, L.-S.; Wu, C.-H.; Yang, W.-C.V.; Wu, A.T.H. Sulforaphane Potentiates the Efficacy of Imatinib against Chronic Leukemia Cancer Stem Cells through Enhanced Abrogation of Wnt/β-Catenin Function. J. Agric. Food Chem. 2012, 60, 7031–7039. [Google Scholar] [CrossRef]
- Carrasco-Pozo, C.; Tan, K.N.; Rodriguez, T.; Avery, V.M. The Molecular Effects of Sulforaphane and Capsaicin on Metabolism upon Androgen and Tip60 Activation of Androgen Receptor. Int. J. Mol. Sci. 2019, 20, 5384. [Google Scholar] [CrossRef]
- Clarke, J.D.; Dashwood, R.H.; Ho, E. Multi-Targeted Prevention of Cancer by Sulforaphane. Cancer Lett. 2008, 269, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Castro, N.P.; Rangel, M.C.; Merchant, A.S.; MacKinnon, G.; Cuttitta, F.; Salomon, D.S.; Kim, Y.S. Sulforaphane Suppresses the Growth of Triple-Negative Breast Cancer Stem-like Cells In Vitro and In Vivo. Cancer Prev. Res. 2019, 12, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Tsubura, A. Sulforaphane Inhibits the Growth of KPL-1 Human Breast Cancer Cells In Vitro and Suppresses the Growth and Metastasis of Orthotopically Transplanted KPL-1 Cells in Female Athymic Mice. Oncol. Rep. 2011, 26, 603–608. [Google Scholar] [CrossRef]
- Lu, Z.; Ren, Y.; Yang, L.; Jia, A.; Hu, Y.; Zhao, Y.; Zhao, W.; Yu, B.; Zhao, W.; Zhang, J.; et al. Inhibiting Autophagy Enhances Sulforaphane-Induced Apoptosis via Targeting NRF2 in Esophageal Squamous Cell Carcinoma. Acta Pharm. Sin. B 2021, 11, 1246–1260. [Google Scholar] [CrossRef]
- Byun, S.; Shin, S.H.; Park, J.; Lim, S.; Lee, E.; Lee, C.; Sung, D.; Farrand, L.; Lee, S.R.; Kim, K.H.; et al. Sulforaphene Suppresses Growth of Colon Cancer-Derived Tumors via Induction of Glutathione Depletion and Microtubule Depolymerization. Mol. Nutr. Food Res. 2016, 60, 1068–1078. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, Z.; Zhou, C.; Chen, K.; Li, X.; Wang, Z.; Wu, Z.; Ma, J.; Ma, Q.; Duan, W. Activation of Nrf2 by Sulforaphane Inhibits High Glucose-Induced Progression of Pancreatic Cancer via AMPK Dependent Signaling. Cell Physiol. Biochem. 2018, 50, 1201–1215. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, J.; Ge, M.; Zhang, Q.; Wang, X.; Zhu, J.; Xie, C.; Li, X.; Zhong, C.; Han, H. Sulforaphane Inhibits Epithelial–Mesenchymal Transition by Activating Extracellular Signal-Regulated Kinase 5 in Lung Cancer Cells. J. Nutr. Biochem. 2019, 72, 108219. [Google Scholar] [CrossRef]
- Rai, R.; Gong Essel, K.; Mangiaracina Benbrook, D.; Garland, J.; Daniel Zhao, Y.; Chandra, V. Preclinical Efficacy and Involvement of AKT, MTOR, and ERK Kinases in the Mechanism of Sulforaphane against Endometrial Cancer. Cancers 2020, 12, 1273. [Google Scholar] [CrossRef]
- Rudzinska-Radecka, M.; Janczewski, Ł.; Gajda, A.; Godlewska, M.; Chmielewska-Krzesinska, M.; Wasowicz, K.; Podlasz, P. The Anti-Tumoral Potential of Phosphonate Analog of Sulforaphane in Zebrafish Xenograft Model. Cells 2021, 10, 3219. [Google Scholar] [CrossRef]
- Eom, Y.S.; Shah, F.H.; Kim, S.J. Sulforaphane Induces Cell Differentiation, Melanogenesis and Also Inhibit the Proliferation of Melanoma Cells. Eur. J. Pharmacol. 2022, 921, 174894. [Google Scholar] [CrossRef]
- Chen, C.Y.; Kao, C.L.; Liu, C.M. The Cancer Prevention, Anti-Inflammatory and Anti-Oxidation of Bioactive Phytochemicals Targeting the TLR4 Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 2729. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Q.; Liu, J.; Zhang, Z.; Ma, X.; Zhang, Y.; Zhu, J.; Thring, R.W.; Wu, M.; Gao, Y.; et al. Sulforaphane Alleviates High Fat Diet-Induced Insulin Resistance via AMPK/Nrf2/GPx4 Axis. Biomed. Pharmacother. 2022, 152, 113273. [Google Scholar] [CrossRef] [PubMed]
- Axelsson, A.S.; Tubbs, E.; Mecham, B.; Chacko, S.; Nenonen, H.A.; Tang, Y.; Fahey, J.W.; Derry, J.M.J.; Wollheim, C.B.; Wierup, N.; et al. Sulforaphane Reduces Hepatic Glucose Production and Improves Glucose Control in Patients with Type 2 Diabetes. Sci. Transl. Med. 2017, 9, eaah4477. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, H.; Chen, X. Protective Effects of Sulforaphane on Diabetic Retinopathy: Activation of the Nrf2 Pathway and Inhibition of NLRP3 Inflammasome Formation. Exp. Anim. 2019, 68, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, X.; Zhou, W.; Men, H.; Bao, T.; Sun, Y.; Wang, Q.; Tan, Y.; Keller, B.B.; Tong, Q.; et al. Ferroptosis Is Essential for Diabetic Cardiomyopathy and Is Prevented by Sulforaphane via AMPK/NRF2 Pathways. Acta Pharm. Sin. B 2022, 12, 708–722. [Google Scholar] [CrossRef]
- Khaleel, S.A.; Raslan, N.A.; Alzokaky, A.A.; Ewees, M.G.; Ashour, A.A.; Abdel-Hamied, H.E.; Abd-Allah, A.R. Contrast Media (Meglumine Diatrizoate) Aggravates Renal Inflammation, Oxidative DNA Damage and Apoptosis in Diabetic Rats Which Is Restored by Sulforaphane through Nrf2/HO-1 Reactivation. Chem. Biol. Interact. 2019, 309, 108689. [Google Scholar] [CrossRef] [PubMed]
- Bastaki, S. Diabetes Mellitus and Its Treatment. Int. J. Diabetes Metab. 2005, 13, 111–134. [Google Scholar] [CrossRef]
- Liu, Y.; Fu, X.; Chen, Z.; Luo, T.; Zhu, C.; Ji, Y.; Bian, Z. The Protective Effects of Sulforaphane on High-Fat Diet-Induced Obesity in Mice through Browning of White Fat. Front. Pharmacol. 2021, 12, 665894. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, Y.; Lou, Y.; Cui, W.; Miao, L. Sulforaphane Suppresses Obesity-Related Glomerulopathy-Induced Damage by Enhancing Autophagy via Nrf2. Life Sci. 2020, 258, 118153. [Google Scholar] [CrossRef]
- Poletto Bonetto, J.H.; Luz de Castro, A.; Fernandes, R.O.; Corssac, G.B.; Cordero, E.A.; Schenkel, P.C.; Sander da Rosa Araujo, A.; Belló-Klein, A. Sulforaphane Effects on Cardiac Function and Calcium-Handling–Related Proteins in 2 Experimental Models of Heart Disease: Ischemia-Reperfusion and Infarction. J. Cardiovasc. Pharmacol. 2022, 79, 325–334. [Google Scholar] [CrossRef]
- Jayakumar, T.; Chen, W.-F.; Lu, W.-J.; Chou, D.-S.; Hsiao, G.; Hsu, C.-Y.; Sheu, J.-R.; Hsieh, C.-Y. A Novel Antithrombotic Effect of Sulforaphane via Activation of Platelet Adenylate Cyclase: Ex Vivo and In Vivo Studies. J. Nutr. Biochem. 2013, 24, 1086–1095. [Google Scholar] [CrossRef]
- Zhang, J.; Dong, Y.; Zhou, M.; Wo, X.; Niu, S.; Shao, E.; Liu, X.; Jin, H.; Zhao, W. Sulforaphane Protects Myocardium from Ischemia-Reperfusion Injury by Regulating CaMKIIN2 and CaMKIIδ. Biochem. Biophys. Res. Commun. 2022, 605, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Chen, Q.; Sun, Y.; Wang, X.; Lv, L.; Zhang, L.; Liu, J.; Zhao, S.; Wang, X. Sulforaphane Protection against the Development of Doxorubicin-induced Chronic Heart Failure Is Associated with Nrf2 Upregulation. Cardiovasc. Ther. 2017, 35, e12277. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.-T.; Yang, H.-Y.; Wang, W.; Wu, Q.-Q.; Tian, Y.-R.; Jia, J.-P. Sulforaphane Inhibits the Generation of Amyloid-β Oligomer and Promotes Spatial Learning and Memory in Alzheimer’s Disease (PS1V97L) Transgenic Mice. J. Alzheimer’s Dis. 2018, 62, 1803–1813. [Google Scholar] [CrossRef]
- Bahn, G.; Park, J.-S.; Yun, U.J.; Lee, Y.J.; Choi, Y.; Park, J.S.; Baek, S.H.; Choi, B.Y.; Cho, Y.S.; Kim, H.K.; et al. NRF2/ARE Pathway Negatively Regulates BACE1 Expression and Ameliorates Cognitive Deficits in Mouse Alzheimer’s Models. Proc. Natl. Acad. Sci. USA 2019, 116, 12516–12523. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.; Zhang, M.; Chen, Z.; Wu, X.; Xia, Z.; Chai, J.; Yin, X. Sulforaphane Prevents PC12 Cells from Oxidative Damage via the Nrf2 Pathway. Mol. Med. Rep. 2019, 19, 4890–4896. [Google Scholar] [CrossRef]
- Zhang, R.; Miao, Q.-W.; Zhu, C.-X.; Zhao, Y.; Liu, L.; Yang, J.; An, L. Sulforaphane Ameliorates Neurobehavioral Deficits and Protects the Brain from Amyloid β Deposits and Peroxidation in Mice with Alzheimer-Like Lesions. Am. J. Alzheimer’s Dis. Other Dementias® 2015, 30, 183–191. [Google Scholar] [CrossRef]
- Lee, S.; Choi, B.; Kim, J.; LaFerla, F.M.; Park, J.H.Y.; Han, J.; Lee, K.W.; Kim, J. Sulforaphane Upregulates the Heat Shock Protein Co-Chaperone CHIP and Clears Amyloid-β and Tau in a Mouse Model of Alzheimer’s Disease. Mol. Nutr. Food Res. 2018, 62, 1800240. [Google Scholar] [CrossRef] [PubMed]
- Pu, D.; Zhao, Y.; Chen, J.; Sun, Y.; Lv, A.; Zhu, S.; Luo, C.; Zhao, K.; Xiao, Q. Protective Effects of Sulforaphane on Cognitive Impairments and AD-like Lesions in Diabetic Mice Are Associated with the Upregulation of Nrf2 Transcription Activity. Neuroscience 2018, 381, 35–45. [Google Scholar] [CrossRef]
- Wang, W.; Wei, C.; Quan, M.; Li, T.; Jia, J. Sulforaphane Reverses the Amyloid-β Oligomers Induced Depressive-Like Behavior. J. Alzheimer’s Dis. 2020, 78, 127–137. [Google Scholar] [CrossRef]
- Morroni, F.; Tarozzi, A.; Sita, G.; Bolondi, C.; Zolezzi Moraga, J.M.; Cantelli-Forti, G.; Hrelia, P. Neuroprotective Effect of Sulforaphane in 6-Hydroxydopamine-Lesioned Mouse Model of Parkinson’s Disease. Neurotoxicology 2013, 36, 63–71. [Google Scholar] [CrossRef]
- Zhou, Q.; Chen, B.; Wang, X.; Wu, L.; Yang, Y.; Cheng, X.; Hu, Z.; Cai, X.; Yang, J.; Sun, X.; et al. Sulforaphane Protects against Rotenone-Induced Neurotoxicity in Vivo: Involvement of the MTOR, Nrf2 and Autophagy Pathways. Sci. Rep. 2016, 6, 32206. [Google Scholar] [CrossRef] [PubMed]
- Jazwa, A.; Rojo, A.I.; Innamorato, N.G.; Hesse, M.; Fernández-Ruiz, J.; Cuadrado, A. Pharmacological Targeting of the Transcription Factor Nrf2 at the Basal Ganglia Provides Disease Modifying Therapy for Experimental Parkinsonism. Antioxid. Redox Signal. 2011, 14, 2347–2360. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Cui, W.; Liu, J.; Li, R.; Liu, Q.; Xie, X.-H.; Ge, X.-L.; Zhang, J.; Song, X.-J.; Wang, Y.; et al. Sulforaphane Ameliorates the Development of Experimental Autoimmune Encephalomyelitis by Antagonizing Oxidative Stress and Th17-Related Inflammation in Mice. Exp. Neurol. 2013, 250, 239–249. [Google Scholar] [CrossRef]
- Yoo, I.-H.; Kim, M.-J.; Kim, J.; Sung, J.-J.; Park, S.T.; Ahn, S.-W. The Anti-Inflammatory Effect of Sulforaphane in Mice with Experimental Autoimmune Encephalomyelitis. J. Korean Med. Sci. 2019, 34, e197. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Li, P.; Yu, L.H.; Li, L.; Long, M.; Liu, M.D.; He, J. Bin Sulforaphane Protect against Cadmium-Induced Oxidative Damage in Mouse Leydigs Cells by Activating Nrf2/ARE Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 630. [Google Scholar] [CrossRef]
- He, Q.; Luo, Y.; Xie, Z. Sulforaphane Ameliorates Cadmium Induced Hepatotoxicity through the Up-Regulation of /Nrf2/ARE Pathway and the Inactivation of NF-ΚB. J. Funct. Foods 2021, 77, 104297. [Google Scholar] [CrossRef]
- Lv, Y.; Jiang, H.; Li, S.; Han, B.; Liu, Y.; Yang, D.; Li, J.; Yang, Q.; Wu, P.; Zhang, Z. Sulforaphane Prevents Chromium-Induced Lung Injury in Rats via Activation of the Akt/GSK-3β/Fyn Pathway. Environ. Pollut. 2020, 259, 113812. [Google Scholar] [CrossRef]
- Thangapandiyan, S.; Ramesh, M.; Miltonprabu, S.; Hema, T.; Jothi, G.B.; Nandhini, V. Sulforaphane Potentially Attenuates Arsenic-Induced Nephrotoxicity via the PI3K/Akt/Nrf2 Pathway in Albino Wistar Rats. Environ. Sci. Pollut. Res. 2019, 26, 12247–12263. [Google Scholar] [CrossRef] [PubMed]
- Thangapandiyana, S.; Ramesha, M.; Hemaa, T.; Uddinc, S.M.M.S.; Nandhinia, V.; Jothia, G.B. Sulforaphane Potentially Ameliorates Arsenic Induced Hepatotoxicity in Albino Wistar Rats: Implication of PI3K/Akt/Nrf2 Signaling Pathway. Cell Physiol. Biochem. 2019, 52, 1203–1222. [Google Scholar] [CrossRef]
- Yang, D.; Han, B.; Baiyun, R.; Lv, Z.; Wang, X.; Li, S.; Lv, Y.; Xue, J.; Liu, Y.; Zhang, Z. Sulforaphane Attenuates Hexavalent Chromium-Induced Cardiotoxicity via the Activation of the Sesn2/AMPK/Nrf2 Signaling Pathway. Metallomics 2020, 12, 2009–2020. [Google Scholar] [CrossRef]
- Hong, L.; Xu, Y.; Wang, D.; Zhang, Q.; Li, X.; Xie, C.; Wu, J.; Zhong, C.; Fu, J.; Geng, S. Sulforaphane Ameliorates Bisphenol A-Induced Hepatic Lipid Accumulation by Inhibiting Endoplasmic Reticulum Stress. Sci. Rep. 2023, 13, 1147. [Google Scholar] [CrossRef]
- Ogunlade, B.; Adelakun, S.; Iteire, K. Sulforaphane Response on Aluminum-Induced Oxidative Stress, Alterations in Sperm Characterization and Testicular Histomorphometry in Wistar Rats. Int. J. Reprod. Biomed. 2020, 18, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Bozic, D.; Živančević, K.; Baralić, K.; Miljaković, E.A.; Djordjević, A.B.; Ćurčić, M.; Bulat, Z.; Antonijević, B.; Đukić-Ćosić, D. Conducting Bioinformatics Analysis to Predict Sulforaphane-Triggered Adverse Outcome Pathways in Healthy Human Cells. Biomed. Pharmacother. 2023, 160, 114316. [Google Scholar] [CrossRef] [PubMed]
- Živančević, K.; Bozic, D.; Baralić, K.; Ćurčić, M.; Antonijević Miljaković, E.; Antonijević, B.; Đukić-Ćosić, D. In Silico Prediction of Physicochemical, Pharmacokinetic and Toxicological Properties of Sulforaphane. Maced. Pharm. Bull. 2022, 68, 331–332. [Google Scholar] [CrossRef]
- Socała, K.; Nieoczym, D.; Kowalczuk-Vasilev, E.; Wyska, E.; Wlaź, P. Increased Seizure Susceptibility and Other Toxicity Symptoms Following Acute Sulforaphane Treatment in Mice. Toxicol. Appl. Pharmacol. 2017, 326, 43–53. [Google Scholar] [CrossRef]
- Tao, S.; Rojo de la Vega, M.; Chapman, E.; Ooi, A.; Zhang, D.D. The Effects of NRF2 Modulation on the Initiation and Progression of Chemically and Genetically Induced Lung Cancer. Mol. Carcinog. 2018, 57, 182–192. [Google Scholar] [CrossRef]
- Kombairaju, P.; Ma, J.; Thimmulappa, R.K.; Yan, S.G.; Gabrielson, E.; Singh, A.; Biswal, S. Prolonged Sulforaphane Treatment Does Not Enhance Tumorigenesis in Oncogenic K-Ras and Xenograft Mouse Models of Lung Cancer. J. Carcinog. 2012, 11, 8. [Google Scholar] [CrossRef]
- Ushida, Y.; Suganuma, H.; Yanaka, A. Low-Dose of the Sulforaphane Precursor Glucoraphanin as a Dietary Supplement Induces Chemoprotective Enzymes in Humans. Food Nutr. Sci. 2015, 06, 1603–1612. [Google Scholar] [CrossRef]
- AICR. Broccoli and Cruciferous Vegetables: Reduce Overall Cancer Risk. Available online: https://www.aicr.org/cancer-prevention/food-facts/broccoli-cruciferous-vegetables/ (accessed on 13 December 2023).
- NHS. 5 a Day Portion Sizes. Available online: https://www.nhs.uk/live-well/eat-well/5-a-day/portion-sizes/ (accessed on 13 December 2023).


{kind=link}
| Animal Species | Treatment Doses | Duration | Results | References |
|---|---|---|---|---|
| Transgenic zebrafish Tg | 3 µmol phosphate derivative of sulforaphane | 48 h |
| Rudzinska-Radecka et al. (2021) [48] |
| Transgenic zebrafish Tg | 30 and 50 µmol |
| Eom et al. (2022) [49] | |
| Balb/C nude mice | 50 mg/kg ip | 5 weeks |
| Castro et al. (2019) [41] |
| Athymic BALB/c mice transplanted with KPL-1 cells | 25 and 50 mg/kg ip | 26 days, 5 times per week |
| Tsubura et al. (2011) [42] |
| Nude mice |
1 and 5 mg/kg/day ip | 13 days |
| Byun et al. (2016) [44] |
| BALB/c mice inoculated with ECa109 cells |
5 mg/kg, ip | 2 weeks, every other day |
| Lu et al. (2021) [43] |
| BALB/c nude mice (transgenic pancreatic cancer mice) |
50 mg/kg, ip | 120 days, every other day |
| Chen et al. (2018) [50] |
| SCID mice inoculated with Ishikawa cells | 50 mg/kg ip | 30 days |
| Rai et al. (2020) [47] |
| BALB/c nude female mice inoculated with H1299 |
25 and 50 mg/kg ip | 21 days, every 3 days |
| Chen et al. (2019) [46] |
| Effects | Animal Species | Treatment Doses | Duration | Results | References |
|---|---|---|---|---|---|
| Antidiabetic/anti-obesogenic effects | C57BL/6 mice |
10 mg/kg ip | 30 days |
| Liu et al. (2021) [57] |
| Nrf2−/− (knockout) mice with a C57BL6/J genetic background Wild-type mice | 10 mg/kg ip | 2 months, every other day |
| Lu et al. (2020) [58] | |
|
Wistar rats C57BL/6J BomTAC mice C57BL/6j mice |
2.5 mg/kg 5 mg/kg 10 mg/kg 10 mg/kg ip | 15 weeks; 3 times per week; 14 days; 9–12 days; 4 weeks |
| Axelsson et al. (2017) [52] | |
|
AMPKalfa2 knockout mice Wild-type C57BL/6J mice |
0.5 mg/kg sc | 3 months, 5 times per week |
| Wang et al. (2022) [32] | |
| Wistar rats |
3 mg/kg ip | Single dose |
| Khaleel et al. (2019) [55] | |
| Male Sprague Dawley rats |
1 mg/kg 0.5 mg/kg ip | 12 weeks |
| Li et al. (2019) [53] |
| Effects | Animal Species | Treatment Doses | Duration | Results | References |
|---|---|---|---|---|---|
| Cardiovascular health benefits | ICR strain mice | 0.125–0.5 mg/kg iv | Single dose |
| Jayakumar et al. (2013) [60] |
| C57BL/6N mice | 50 µg/kg ip | 3 days |
| Zhang et al. (2022) [61] | |
| MI model rats | 5 mg/kg ip | 25 days |
| Poletto Bonetto et al. (2022) [59] | |
| Sprague Dawley rats | 0.5 mg/kg sc | 6 weeks |
| Bai et al. (2017) [62] |
| Effects | Animal Species | Treatment Doses | Duration | Results | References |
|---|---|---|---|---|---|
| Neuroprotective effects | C57BL/6 mice |
25 mg/kg oral | 80 days |
| Zhang et al. (2015) [66] |
| 3 × Tg-AD mice |
10 or 50 mg/kg gavage | 8 weeks, 6 days per week |
| Lee et al. (2018) [67] | |
| Type II diabetes mellitus transgenic mice | 1 mg/kg ip | 28 days |
| Pu et al. (2018) [68] | |
| PS1V97L transgenic mice | 5 mg/kg ip | 4 months |
| Hou et al. (2018) [63] | |
| C57Bl/6 mice | 5 mg/kg ip | 4 weeks, 2 times per week |
| Morroni et al. (2013) [70] | |
| C57BL/6 mice | 50 mg/kg ip | 60 days, every other day |
| Zhou et al. (2016) [71] | |
|
Wild-type mice Nrf2-KO mice | 50 mg/kg ip |
| Jazwa et al. (2011) [72] | ||
| EAE C57Bl/6 mice | 50 mg/kg ip | 14 days |
| Yoo et al. (2019) [74] | |
| C57BL/6 mice | 50 mg/kg ip | 22 days |
| Li et al. (2013) [73] | |
| Sprague Dawley male rats | 5 mg/kg ip | 7 days |
| Wang et al. (2020) [69] |
| Species | Toxic Substance | Sulforaphane (SFN) | Results | References |
|---|---|---|---|---|
| human hepatocyte cell line (LO2) and C57/BL6J mice | bisphenol A (BPA): 100, 1000 nM for 24 h | SFN (0.25 μM, 0.5 μM) for 24 h |
| Hong et al., 2023 [81] |
| mouse Leydig (TM3) cells | Cadmium (Cd): 10 μmol/L for 24 h | SFN (2.5, 5, 10 μmol/L) for 24 h |
| Yang et al., 2019 [75] |
| mouse HepG2 cells | in vitro: cadmium chloride (CdCl2): 20 μM for 24 h; in vivo: cadmium chloride (CdCl2): 10 mg/kg b.w/day, per os, 4 weeks | in vitro: SFN (0–80 μM) for 24 h; in vivo: SFN (0.5, 1, 2 mg/kg b.w/day), per os, 6 weeks |
| He et al., 2021 [76] |
| mouse alveolar type II epithelial cell line (MLE-12) | in vitro: potassium dichromate (K2Cr2O7): 1 mg/mL for 24 h after SFN | in vitro: pretreating with 0.1 mM SFN for 30 min |
| Lv et al., 2020 [77] |
| 35-day Cr-induced pulmonary toxicity model | in vivo: potassium dichromate (K2Cr2O7): 6, 4, and 2 mg/kg b.w/day, per os, 35 days | in vivo: SFN (4 mg/kg b.w/day), subcutaneous injection |
| |
| rats | potassium dichromate (K2Cr2O7): 4 mg/kg b.w/day, intraperitoneal injection, 4 weeks | SFN (4 mg/kg b.w/day), subcutaneous injection 1 h after K2Cr2O7 treatment, 4 weeks |
| Yang et al., 2020 [80] |
| rats | sodium arsenite (NaAsO2): 5 mg/kg b.w/day, per os, 28 days | SFN (80 mg/kg b.w/day), per os, 28 days |
| Thangapandiyan et al., 2019 [78] |
| rats | sodium arsenite (NaAsO2) (5 mg/kg b.w/day), oral, 4 weeks | SFN (20, 40, 80 mg/kg b.w/day), oral, 4 weeks (administered 90 min before As) |
| Thangapandiyana et al., 2019 [79] |
| rats | aluminum chloride (AlCl3) (100 mg/kg b.w/day), per os | SFN (100 mg/kg b.w), per os |
| Ogunlade et al., 2020 [82] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baralić, K.; Živanović, J.; Marić, Đ.; Bozic, D.; Grahovac, L.; Antonijević Miljaković, E.; Ćurčić, M.; Buha Djordjevic, A.; Bulat, Z.; Antonijević, B.; et al. Sulforaphane—A Compound with Potential Health Benefits for Disease Prevention and Treatment: Insights from Pharmacological and Toxicological Experimental Studies. Antioxidants 2024, 13, 147. https://doi.org/10.3390/antiox13020147
Baralić K, Živanović J, Marić Đ, Bozic D, Grahovac L, Antonijević Miljaković E, Ćurčić M, Buha Djordjevic A, Bulat Z, Antonijević B, et al. Sulforaphane—A Compound with Potential Health Benefits for Disease Prevention and Treatment: Insights from Pharmacological and Toxicological Experimental Studies. Antioxidants. 2024; 13(2):147. https://doi.org/10.3390/antiox13020147
Chicago/Turabian StyleBaralić, Katarina, Jovana Živanović, Đurđica Marić, Dragica Bozic, Lazar Grahovac, Evica Antonijević Miljaković, Marijana Ćurčić, Aleksandra Buha Djordjevic, Zorica Bulat, Biljana Antonijević, and et al. 2024. "Sulforaphane—A Compound with Potential Health Benefits for Disease Prevention and Treatment: Insights from Pharmacological and Toxicological Experimental Studies" Antioxidants 13, no. 2: 147. https://doi.org/10.3390/antiox13020147
APA StyleBaralić, K., Živanović, J., Marić, Đ., Bozic, D., Grahovac, L., Antonijević Miljaković, E., Ćurčić, M., Buha Djordjevic, A., Bulat, Z., Antonijević, B., & Đukić-Ćosić, D. (2024). Sulforaphane—A Compound with Potential Health Benefits for Disease Prevention and Treatment: Insights from Pharmacological and Toxicological Experimental Studies. Antioxidants, 13(2), 147. https://doi.org/10.3390/antiox13020147