
Effect of Chronically Suppressed Plasma Angiotensin II on Regulation of the CYP4A/20-HETE Pathway in the Dahl Salt-Sensitive Rat

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Groups
2.2. Isolated Vessel Experiments
2.3. 20-HETE Production in Cerebral Arteries
2.4. Dihydroethidium Fluorescence
2.5. Statistical Analysis
3. Results
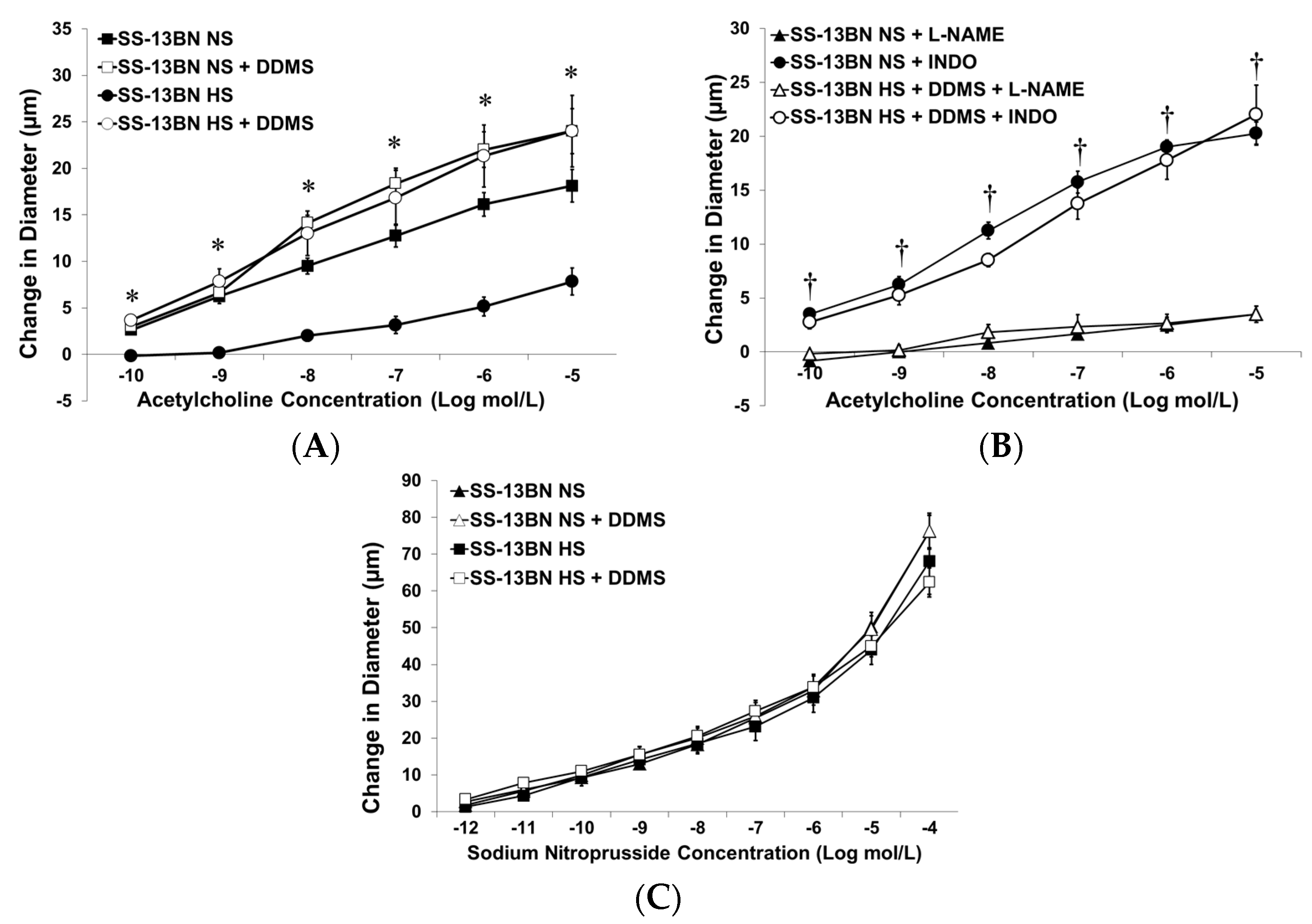
3.1. Vascular Responses
3.1.1. Vessel Diameter and Active Tone in SS-13BN Consomic Rats and Dahl SS Rats Receiving Osmotic Minipumps
3.1.2. Effect of Chromosome 13 Substitution on Responses of MCA to Acetylcholine (ACh)
3.1.3. Effect of Low-Dose ANG II Infusion on Vascular Responses in MCA from Dahl SS Rats
3.2. Vascular 20-HETE Production
3.2.1. 20-HETE Levels in Cerebral Arteries of SS-13BN Rats
3.2.2. 20-HETE Levels in Cerebral Arteries from Saline- or ANGII-Infused Dahl SS Rats
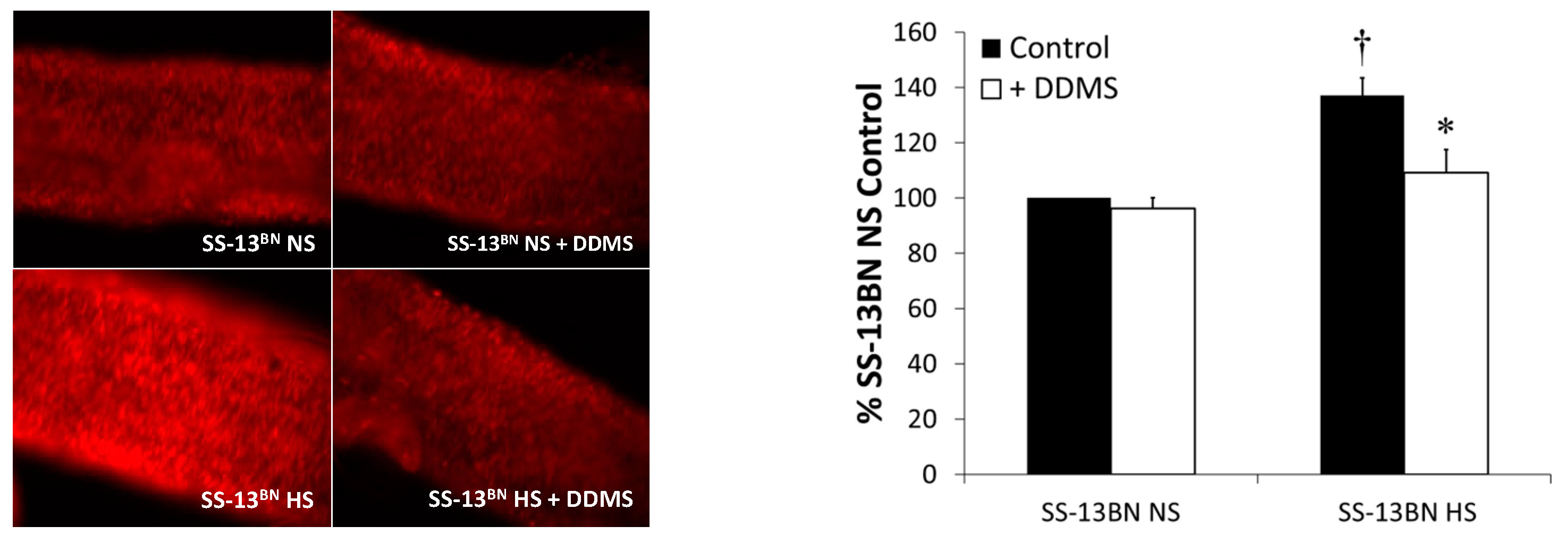
3.3. Dihydroethidium (DHE) Fluorescence
Vascular Reactive Oxygen Species Evaluated by DHE Fluorescence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Preston, R.A.; Materson, B.J.; Reda, D.J.; Williams, D.W.; Hamburger, R.J.; Cushman, W.C.; Anderson, R.J. Age-race subgroup compared with renin profile as predictors of blood pressure response to antihypertensive therapy. Department of Veterans Affairs Cooperative Study Group on Antihypertensive Agents. JAMA 1998, 280, 1168–1172. [Google Scholar] [CrossRef]
- Price, D.A.; Fisher, N.D. The renin-angiotensin system in blacks: Active, passive, or what? Curr. Hypertens. Rep. 2003, 5, 225–230. [Google Scholar] [CrossRef]
- Mackenzie, I.S.; Brown, M.J. Molecular and clinical investigations in patients with low-renin hypertension. Clin. Exp. Nephrol. 2009, 13, 1–8. [Google Scholar] [CrossRef]
- Helmer, O.M.; Judson, W.E. Metabolic studies on hypertensive patients with suppressed plasma renin activity not due to hyperaldosternosm. Circulation 1968, 38, 965–976. [Google Scholar] [CrossRef]
- Rapp, J.P.; Wang, S.M.; Dene, H. A genetic polymorphism in the renin gene of Dahl rats cosegregates with blood pressure. Science 1989, 243, 542–544. [Google Scholar] [CrossRef]
- Rapp, J.P.; McPartland, R.P.; Sustarsic, D.L. A qualitative difference in plasma renin activity in Dahl rats susceptible or resistant to salt-induced hypertension. Biochem. Genet. 1980, 18, 1087–1096. [Google Scholar] [CrossRef]
- Weber, D.S.; Lombard, J.H. Angiotensin II AT1 receptors preserve vasodilator reactivity in skeletal muscle resistance arteries. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H2196–H2202. [Google Scholar] [CrossRef]
- Weber, D.S.; Lombard, J.H. Elevated salt intake impairs dilation of rat skeletal muscle resistance arteries via ANG II suppression. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H500–H506. [Google Scholar] [CrossRef]
- Lombard, J.H.; Sylvester, F.A.; Phillips, S.A.; Frisbee, J.C. High-salt diet impairs vascular relaxation mechanisms in rat middle cerebral arteries. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H1124–H1133. [Google Scholar] [CrossRef]
- Zhu, J.; Huang, T.; Lombard, J.H. Effect of high-salt diet on vascular relaxation and oxidative stress in mesenteric resistance arteries. J. Vasc. Res. 2007, 44, 382–390. [Google Scholar] [CrossRef]
- Zhu, J.; Mori, T.; Huang, T.; Lombard, J.H. Effect of high-salt diet on NO release and superoxide production in rat aorta. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H575–H583. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Drenjancevic-Peric, I.; McEwen, S.; Friesema, J.; Schulta, D.; Yu, M.; Roman, R.J.; Lombard, J.H. Role of superoxide and angiotensin II suppression in salt-induced changes in endothelial Ca2+ signaling and NO production in rat aorta. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H929–H938. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.J.; Lombard, J.H. Low-dose angiotensin II infusion restores vascular function in cerebral arteries of high salt-fed rats by increasing copper/zinc superoxide dimutase expression. Am. J. Hypertens. 2013, 26, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Lukaszewicz, K.M.; Lombard, J.H. Role of the CYP4A/20-HETE pathway in vascular dysfunction of the Dahl salt-sensitive rat. Clin. Sci. 2013, 124, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.J.; Lombard, J.H. Introgression of the Brown Norway renin allele onto the Dahl salt-sensitive genetic background increases Cu/Zn SOD expression in cerebral arteries. Am. J. Hypertens. 2011, 24, 563–568. [Google Scholar] [CrossRef]
- Cowley, A.W.; Roman, R.J.; Kaldunski, M.L.; Dumas, P.; Dickhout, J.G.; Greene, A.S.; Jacob, H.J. Brown Norway chromosome 13 confers protection from high salt to consomic Dahl S rat. Hypertension 2001, 37, 456–461. [Google Scholar] [CrossRef]
- Drenjancevic-Peric, I.; Lombard, J.H. Introgression of chromosome 13 in Dahl salt-sensitive genetic background restores cerebral vascular relaxation. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H957–H962. [Google Scholar] [CrossRef]
- Durand, M.J.; Moreno, C.; Greene, A.S.; Lombard, J.H. Impaired Relaxation of Cerebral Arteries in the Absence of Elevated Salt Intake in Normotensive Congenic Rats Carrying the Dahl Salt-Sensitive Renin Gene. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1865–H1874. [Google Scholar] [CrossRef]
- Drenjancevic-Peric, I.; Lombard, J.H. Reduced angiotensin II and oxidative stress contribute to impaired vasodilation in Dahl salt-sensitive rats on low-salt diet. Hypertension 2005, 45, 687–691. [Google Scholar] [CrossRef]
- Mattson, D.L.; Dwinell, M.R.; Greene, A.S.; Kwitek, A.E.; Roman, R.J.; Jacob, H.J.; Cowley, A.W., Jr. Chromosome substitution reveals the genetic basis of Dahl salt-sensitive hypertension and renal disease. Am. J. Physiol. Renal Physiol. 2008, 295, F837–F842. [Google Scholar] [CrossRef]
- Wang, J.; Roman, R.J.; Falck, J.R.; de la Cruz, L.; Lombard, J.H. Effects of high-salt diet on CYP450-4A omega-hydroxylase expression and active tone in mesenteric resistance arteries. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H1557–H1565. [Google Scholar] [CrossRef]
- Wang, J.; Schmidt, J.R.; Roman, R.J.; Anjaiah, S.; Falck, J.R.; Lombard, J.H. Modulation of vascular O2 responses by cytochrome 450-4A omega-hydroxylase metabolites in Dahl salt-sensitive rats. Microcirculation 2009, 16, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Matic, A.; Jukic, I.; Mihaljevic, Z.; Kolobaric, N.; Stupin, A.; Kozina, N.; Bujak, I.T.; Kibel, A.; Lombard, J.H.; Drenjancevic, I. Low-dose angiotensin II supplementation restores flow-induced dilation mechanisms in cerebral arteries of Sprague-Dawley rats on a high salt diet. J. Hypertens. 2022, 40, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Fredricks, K.T.; Liu, Y.; Lombard, J.H. Response of extraparenchymal resistance arteries of rat skeletal muscle to reduced PO2. Am. J. Physiol. 1994, 267, H706–H715. [Google Scholar] [CrossRef]
- Dunn, K.M.; Renic, M.; Flasch, A.K.; Harder, D.R.; Falck, J.; Roman, R.J. Elevated production of 20-HETE in the cerebral vasculature contributes to severity of ischemic stroke and oxidative stress in spontaneously hypertensive rats. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H2455–H2465. [Google Scholar] [CrossRef]
- Drenjancevic-Peric, I.; Frisbee, J.C.; Lombard, J.H. Skeletal muscle arteriolar reactivity in SS.BN13 consomic rats and Dahl salt-sensitive rats. Hypertension 2003, 41, 1012–1015. [Google Scholar] [CrossRef]
- Drenjancevic-Peric, I.; Phillips, S.A.; Falck, J.R.; Lombard, J.H. Restoration of normal vascular relaxation mechanisms in cerebral arteries by chromosomal substitution in consomic SS.S.13BN rats. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H188–H195. [Google Scholar] [CrossRef]
- McEwen, S.T.; Schmidt, J.R.; Somberg, L.; Cruz Lde, L.; Lombard, J.H. Time-course and mechanisms of restored vascular relaxation by reduced salt intake and angiotensin II infusion in rats fed a high-salt diet. Microcirculation 2009, 16, 220–234. [Google Scholar] [CrossRef]
- Puntarulo, S.; Cederbaum, A.I. Production of reactive oxygen species by microsomes enriched in specific human cytochrome P450 enzymes. Free Radic. Biol. Med. 1998, 24, 1324–1330. [Google Scholar] [CrossRef]
- Bondy, S.C.; Naderi, S. Contribution of hepatic cytochrome P450 systems to the generation of reactive oxygen species. Biochem. Pharmacol. 1994, 48, 155–159. [Google Scholar] [CrossRef]
- Eid, A.A.; Gorin, Y.; Fagg, B.M.; Maalouf, R.; Barnes, J.L.; Block, K.; Abboud, H.E. Mechanisms of podocyte injury in diabetes: Role of cytochrome P450 and NADPH oxidases. Diabetes 2009, 58, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Hama-Tomioka, K.; Kinoshita, H.; Azma, T.; Nakahata, K.; Matsuda, N.; Hatakeyama, N.; Kikuchi, H.; Hatano, Y. The role of 20-hydroxyeicosatetraenoic acid in cerebral arteriolar constriction and the inhibitory effect of propofol. Anesth. Analg. 2009, 109, 1935–1942. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Han, Y.; Bao, Y.; Li, W.; Li, X.; Shen, X.; Wang, X.; Yao, F.; O’Rourke, S.T.; Sun, C. 20-HETE increases NADPH oxidase-derived ROS production and stimulates the L-type Ca2+ channel via a PKC-dependent mechanism in cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1109–H1117. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Cheng, J.; Deng, H.; Kemp, R.; Ishizuka, T.; Nasjletti, A.; Schwartzman, M.L. Vascular cytochrome P450 4A expression and 20-hydroxyeicosatetraenoic acid synthesis con.ntribute to endothelial dysfunction in androgen-induced hypertension. Hypertension 2007, 50, 123–129. [Google Scholar] [CrossRef]
- Cheng, J.; Ou, J.S.; Singh, H.; Falck, J.R.; Narsimhaswamy, D.; Pritchard, K.A., Jr.; Schwartzman, M.L. 20-hydroxyeicosatetraenoic acid causes endothelial dysfunction via eNOS uncoupling. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1018–H1026. [Google Scholar] [CrossRef]
- Ward, N.C.; Chen, K.; Li, C.; Croft, K.D.; Keaney, J.F., Jr. Chronic activation of AMP-activated protein kinase prevents 20-hydroxyeicosatetraenoic acid-induced endothelial dysfunction. Clin. Exp. Pharmacol. Physiol. 2011, 38, 328–333. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Groups | Number (n) | Max Diameter (µm) | Rest Diameter (µm) | Active Tone (%) |
|---|---|---|---|---|
| SS-13BN 1 NS 2 | 6 | 236 ± 2.0 | 137 ± 4.7 | 42 ± 2.1 |
| SS-13BN HS 3 | 6 | 245 ± 3.9 | 149 ± 3.3 | 39 ± 2.0 |
| SS-13BN NS + DDMS 4 | 6 | 237 ± 2.0 | 140 ± 6.0 | 40 ± 4.7 |
| SS-13BN HS + DDMS | 6 | 245 ± 3.9 | 133 ± 5.16 | 45 ± 2.9 |
| SS-13BN NS + L-NAME 5 | 6 | 246 ± 5.9 | 120 ± 10.1 | 51 ± 3.6 |
| SS-13BN HS + DDMS + L-NAME | 6 | 247 ± 4.7 | 134 ± 6.3 | 45 ± 2.3 |
| SS-13BN NS + INDO 6 | 4 | 257 ± 8.5 | 147 ± 14.6 | 45 ± 7.5 |
| SS-13BN HS + DDMS + INDO | 4 | 239 ± 3.6 | 150 ± 7.3 | 37 ± 3.4 |
| Experimental Groups | Number (n) | Max Diameter (µm) | Rest Diameter (µm) | Active Tone (%) |
|---|---|---|---|---|
| SS 1 + ANG II 2 | 8 | 231 ± 2.6 | 135 ± 2.0 | 42 ± 1.3 |
| SS + Saline | 8 | 235 ± 3.4 | 141 ± 3.2 | 40 ± 1.8 |
| SS + ANG II + DDMS 3 | 8 | 231 ± 2.6 | 145 ± 4.9 | 37 ± 2.5 |
| SS + Saline + DDMS | 8 | 235 ± 3.4 | 142 ± 5.2 | 40 ± 2.7 |
| SS + ANG II + L-NAME 4 | 6 | 240 ± 3.9 | 115 ± 5.4 * | 52 ± 2.4 |
| SS + Saline + DDMS + L-NAME | 6 | 240 ± 4.0 | 125 ± 5.0 | 48 ± 2.0 |
| SS + ANG II + INDO 5 | 3 | 248 ± 8.7 | 151 ± 6.3 | 37 ± 1.7 |
| SS + Saline + DDMS + INDO | 3 | 236 ± 3.5 | 138 ± 5.0 | 41 ± 2.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lukaszewicz, K.; Falck, J.R.; Lombard, J. Effect of Chronically Suppressed Plasma Angiotensin II on Regulation of the CYP4A/20-HETE Pathway in the Dahl Salt-Sensitive Rat. Antioxidants 2023, 12, 783. https://doi.org/10.3390/antiox12040783
Lukaszewicz K, Falck JR, Lombard J. Effect of Chronically Suppressed Plasma Angiotensin II on Regulation of the CYP4A/20-HETE Pathway in the Dahl Salt-Sensitive Rat. Antioxidants. 2023; 12(4):783. https://doi.org/10.3390/antiox12040783
Chicago/Turabian StyleLukaszewicz, Kathleen, John R. Falck, and Julian Lombard. 2023. "Effect of Chronically Suppressed Plasma Angiotensin II on Regulation of the CYP4A/20-HETE Pathway in the Dahl Salt-Sensitive Rat" Antioxidants 12, no. 4: 783. https://doi.org/10.3390/antiox12040783
APA StyleLukaszewicz, K., Falck, J. R., & Lombard, J. (2023). Effect of Chronically Suppressed Plasma Angiotensin II on Regulation of the CYP4A/20-HETE Pathway in the Dahl Salt-Sensitive Rat. Antioxidants, 12(4), 783. https://doi.org/10.3390/antiox12040783