Abstract
Since the gut microbiota plays a pivotal role in host homeostasis and energy balance, changes in its composition can be associated with disease states through the promotion of immune-mediated inflammatory disorders and increasing intestinal permeability, ultimately leading to the impairment of intestinal barrier function. Za’atar is one of the most popular plant-based foods in the Eastern Mediterranean region. Za’atar is a mixture of different plant leaves, fruits, and seeds and contains hundreds of antioxidant compounds, especially polyphenols, and fiber, with pre-clinical and clinical evidence suggesting health-promoting effects in cardiovascular and metabolic disease. Za’atar compounds have also been studied from a gastrointestinal perspective, concerning both gut microbiota and gastrointestinal diseases. Antioxidants such as Za’atar polyphenols may provide beneficial effects in the complex interplay between the diet, gut microbiota, and intestinal permeability. To our knowledge, no studies have reported the effects of the whole Za’atar mixture, however, based on the pre-clinical studies published on components and single compounds found in Za’atar, we provide a clinical overview of the possible effects on the gastrointestinal tract, focusing mainly on carvacrol, rosmarinic acid, gallic acid, and other polyphenols. We also cover the potential clinical applications of Za’atar mixture as a possible nutraceutical in disorders involving the gastrointestinal tract.
1. Introduction
The gastrointestinal tract is considered the largest surface of the human body exposed to the external environment [1]. The gut barrier has several essential functions, especially the absorption of nutrients and defense against harmful macromolecules [2]. The gut barrier is a complex multilayer system consisting of both an external “physical” barrier and an inner “functional” immune barrier. The physiological “healthy” interaction of both barriers ensures the selective permeability of nutrients, water, and bacterial products. Different factors can alter the function, physiology, and homeostasis of the gastrointestinal tract potentially leading to the development of a wide range of functional and inflammatory gastrointestinal disorders. Furthermore, the long-term effects of stress could affect gut-liver and gut-brain interactions. Stress-related factors include altered gastrointestinal motility and increased intestinal permeability with negative effects on the diversity of gut microbiota [3]. A close relationship exists between nutrients (dietary fiber, protein, fat), gut microbiota and its products (e.g., short-chain fatty acids, lipopolysaccharides), and the intestinal barrier in both health and disease [1]. Although enormous effort has been put into studies to investigate the role of dietary intervention in improving intestinal barrier function and preventing increased intestinal permeability in animals, the relevance of these results for human health is still poorly understood. Za’atar mixture is the most popular plant-based food in the Eastern Mediterranean area. The original traditional Lebanese mixture consists of various plants, fruits, and seeds. These plants contain a panel of biologically active compounds—such as polyphenolic compounds (PC), essential oils (EOs), and fiber—some of which have displayed potential health-promoting effects in cardiovascular disease and metabolic syndrome. The effects of the whole Za’atar mixture are still missing, however, studies on some components and compounds found in Za’atar on the gastrointestinal tract have also been studied regarding gut microbiota and intestinal permeability in both animals and humans. Herein we provide an overview of the intestinal barrier from a functional perspective, as well as in terms of intestinal permeability and its association with gut microbiota. In addition, we focused on the Za’atar mixture, described its general history and cultural aspects, and then we detailed the components, chemical/phytochemical composition, and nutritional profile. Finally, we implemented the review with all available data on the potential effects of Za’atar components on the gastrointestinal tract and gut microbiota including cellular, animal, and human studies. Since there are no studies describing the effects of the whole Za’atar mixture, here we focused on the effects of single components and/or single (poly)phenolic compounds to highlight to possible health-promoting effects of the mixture.
2. Gut Physiology and Microbiota
The microbial barrier comprises the first level of the gut barrier and consists of hundreds of trillions of resident microorganisms including bacteria, viruses, fungi, bacteriophages, and protists [4]. In the human gut, the two major phyla include Firmicutes (mainly gram-positive bacteria with facultative anaerobes, anaerobes, bacilli, and cocci) and Bacteroidetes (mainly gram-negative Bacteroides, Alistipes, Parabacteroides, and Prevotella spp.) [5,6]. The gut microbiota plays an essential role in the digestion of nutrient and non-nutrient molecules, vitamin synthesis and metabolism, and the biotransformation of intestinal primary bile acids (BAs) to secondary BAs, which play a key role in lipid digestion and hormone-like agonist signaling. In addition, the gut microbiota has a crucial impact on the induction and function of the human immune system by influencing the maturation of several immune cells in intestinal lymphoid tissue and mucosa. The gut microbiota produces physiologically active molecules such as short-chain fatty acids (SCFAs) that have direct effects on the gut barrier and distant organs including, but not limited to, the brain, liver, and heart [7]. In addition, gut microbiota modulate nutritionally derived metabolites such as tryptophan which exerts a protective effect against intestinal inflammation and bacterial overgrowth [8]. The gut microbiota converts tryptophan to serotonin, transforms food polyphenols to bioactive smaller polyphenols, and affects the availability of neurotransmitters [9,10].
The mucosal barrier is the first line of defense against external pathogens. It separates the external from the internal environment, protects against toxins and the passage of bacteria, and facilitates the transport of water, nutrients, ions, and solutes [11] (Figure 1). The intestinal mucosa plays the dual role of enforcing both defense and digestion by adapting to the colonization by commensal bacteria that participate in digestive processes and the induction of the intestinal immune system [12]. The epithelial intestinal barrier consists of a highly organized complex of intercellular apical tight junction proteins and tight junction-associated proteins including claudin, occludin, intracellular plaque zonula occludens (ZO) 1 and 2, cingulin, and other junctional adhesion proteins [13]. These tight junctions distribute cells into apical and basolateral regions. Despite being the first line of defense in the intestinal lumen, the epithelial barrier manages to simultaneously regulate the passive diffusion of macromolecules and solutes beneficial to the host and to prevent the passage of harmful pathogens [14] (Figure 1).
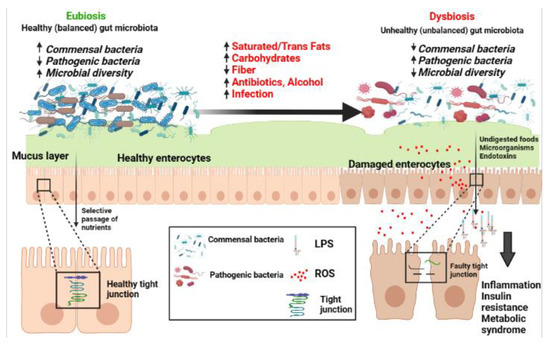
Figure 1.
The transition from a balanced (Eubiosis) to an unbalanced (dysbiosis) gut microbiota and the different factors involved in gut dysbiosis. Abbreviations: LPS: lipopolysaccharide; ROS: reactive oxygen species; ↑ increased; ↓ decreased. The figure is developed using https://biorender.com/ (accessed on 12 December 2022).
The dysbiosis of gut microbiota is defined as an altered microbial community with an increased population of pathogenic bacteria resulting in the increased release of bacterial endotoxins, especially lipopolysaccharides (LPS) that can alter the function of the intestinal barrier on a fundamental level by LPS-induced damage of enterocytes, leading to increased intestinal permeability. The intestine becomes “leaky,” allowing for the passage of LPS into the bloodstream, in turn activating inflammatory pathways resulting in systemic inflammation and potentially metabolic disorders [15,16,17,18].
3. Za’atar Mixture
Over the past ten years, interest in Za’atar has greatly increased within the scientific and industrial communities. Recent advances have been made in unraveling the multilevel properties of Za’atar plants or mixtures in terms of health-promoting effects, as well as cultivation, processing, production, and packing [19,20,21]. Za’atar is of economic and cultural importance in the Mediterranean region. More interestingly, it possesses nutraceutical and bio-functional properties as it is rich in (poly)phenolic compounds (PC), minerals, and fiber. So, it comes as no surprise, that there is both national and international interest in Za’atar [22,23,24].
The term “Za’atar” is used to refer to both a type of plant as well as to a mixture of plants and spices [25]. Za’atar plants include Origanum, Satureja, Thymbra, and Thymus with high content of essential oil content, especially carvacrol and thymol [22,23,24]. As a mixture, the composition of Za’atar can vary by region and household, usually depending on the local availability of ingredients. Typically, in Lebanon, Za’atar mixture includes dried leaves of Origanum syriacum (also referred to as Za’atar) and Thymbra spicata (also known as wild Za’atar) mixed with Rhus coriaria ground fruit (sumac), toasted sesame seeds, and salt (Figure 2). Za’atar is commonly mixed with olive oil to form a spread for Lebanese flatbreads named “Mankoushe” [26]. The detailed components and polyphenolic composition and contents of the Za’atar mixture are shown in Figure 2.
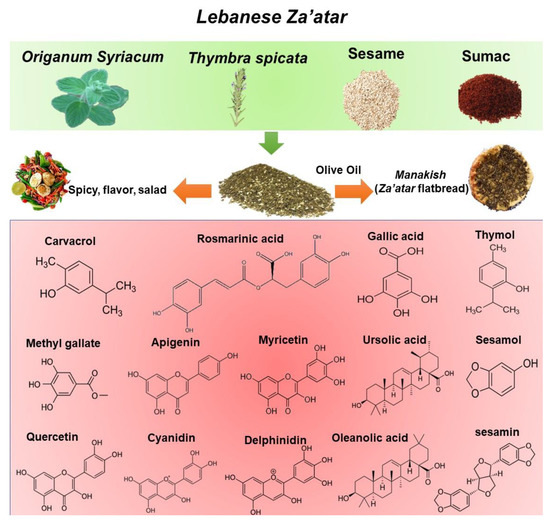
Figure 2.
The composition of Za’atar mixture with polyphenols.
In traditional books on Lebanese herbal medicine Za’atar (or Sa’atar) is presented as a special and distinct class of edible and medicinal herbs (O. syriacum and T. spicata) for the treatment of gastrointestinal diseases. Specifically, it is described that these herbs protect the intestinal mucosal barrier, reduce abdominal pain, and help prevent constipation by keeping the stool moist. These traditional books on herbal medicine also describe the gastroprotective and hepatoprotective effects of Za’atar plants [23]. Sumac, a component of the Za’atar mixture, is traditionally used in folk medicine for the treatment of chronic diarrhea, vomiting, and hemorrhoids. Water infusions of sumac could reduce inflammation, relax and protect the stomach, protect the liver, and help the bile reach the intestine [27].
3.1. Chemical and Phytochemical Composition of Mixed Za’atar
O. syriacum and T. spicata extracts contain high amounts of antioxidant compounds such as polyphenols including phenolic acids and flavonoids with a wide range of biological and pharmacological activities [28,29,30,31,32]. Rosmarinic acid (RA) is one of the main polyphenolic substances in both plants [24,33,34,35]. Ursolic and oleanolic acids are also present in high quantities in O. syriacum extracts [35]. Phenolic monoterpenes, especially carvacrol (CVL), are abundant in both essential oils and organic extracts, with the total essential oil content ranging from 60 to 70% [36,37]. One of the most abundant subclasses of polyphenols present in O. syriacum and T. spicata extracts are flavonoids—mostly luteolin and apigenin in aglycone or glucoside form [38,39] (Figure 2).
Sesame (Sesamum indicum L.) is an annual plant belonging to the Pedaliaceae family. Sesame seeds have been used for several thousands of years in Eastern, Mediterranean, and African cultures to flavor foods [40]. Nutritionally, sesame seeds are rich in oil (50–60%) and protein (18–25%) and contain carbohydrates (13.5%) and ash (5%) [41]. Sesame seeds contain high levels of unsaturated fatty acids—mainly oleic (43%), linoleic (35%), palmitic (11%), and stearic acids (7%)—collectively comprising 96% of the total fatty acid content [42]. Sesame seeds also contain protein, especially high levels of methionine, tocopherol, and phytosterol, as well as minerals and lignans, such as sesamol and sesamin—natural phenolic compounds and major lignans [43] (Table 1).
Rhus coriaria or sumac is rich in hydrolyzable tannins such as gallic acid, methyl gallate, and their derivatives, [44,45]. The most abundant organic acids found in R. coraria fruit are malic acid isomers and their derivatives. Sumac is abundant also in phenolic acids [46] and flavonoids including myricitrin, apigenin, quercetin, and kaempferol as aglycone or glycoside derivatives [46,47]. Cyanidin and delphinidin derivatives are the most abundant anthocyanin present in sumac and give its fruits their rich red color [44].

Table 1.
The components of Za’atar mixture by name and useful contents.
Table 1.
The components of Za’atar mixture by name and useful contents.
| Scientific Name | Local Name (Lebanon) | Used Part | Phenolic Contents mgGAE/g Dry Extract | Fiber g/100 g | Oils/Fatty Acids g/100 g | Essential Oils | Minerals (in Order of Abundance) | Vitamins | References |
|---|---|---|---|---|---|---|---|---|---|
| Origanum Syriacum | Za’atar (Zouba’a) | Leaves | 120–250 | 15 | 0.13 | Carvacrol, thymol | ND | C | [38,48,49,50] |
| Thymbra spicata | Wild Za’atar (Za’atar dakki) | Leaves | 90–250 | ND | PA OA | Carvacrol | ND | ND | [37,38,39] |
| Rhus coriaria | Sumac | Fruits | 48–140 | 14 | 7–18 (37.7% OA, 34.8% LA, 27.4% PA, 17.3% SA) | β-caryophyllene, α-pinene | K, Ca, Mg, P | B1, B2, B6, B12, C | [51,52,53,54] |
| Sesamum indicum | Sesame | Seeds | 7–80 | 6–10.8 | 51.9 OA (43%) LA (35%), PA (11%) SA (7%) | ND | Ca, K, P, Mg, Fe | Carotene, B1, B2, B3, E | [55,56,57] |
Abbreviations: PA—palmitic acid; OA—oleaic acid; SA—stearic acid; LA—linoleic acid, K-Potasium; Ca-calcium; Mg-magnesium; Fe-iron.
3.2. Effects of Za’atar Compounds on Microbiota and Intestinal Disorders: Cellular and Animal Studies
Preclinical evidence including cellular and animal studies explored the potential effects of polyphenols on gut microbiota and gut diseases. These compounds interact bidirectionally with the gut microbiota and can modulate the composition by exerting prebiotic-like mechanisms and inhibiting pathogenic bacteria [58,59]. On the other hand,, after the intake of polyphenol-rich foods, the gut microbiota plays a fundamental role in enhancing biotransformation and bioavailability and, thus, the bio-activity of polyphenols. The effect of polyphenols and polyphenol-rich foods on microbial growth, in vitro, in vivo, and ex vivo is well documented [59]. Studies reported that polyphenols can inhibit the growth of several pathogens, including E. coli, S. enteritidis, S. typhimurium and C. perfringens, L. monocytogenes, H. pylori, B. cereus, P. aeruginosa, and S. aureus [60,61,62,63]. Polyphenols and their metabolites showed a prebiotic-like effect on shaping gut microbiota and modulation of inflammatory responses by enhancing the abundance of beneficial bacteria including Lactobacillus, Bifidobacterium, Faecalibacterium prausnitzii, and Roseburia [64,65,66,67]. In the following paragraphs we report the available studies about the effects of Za’atar compounds on microbiota and gastrointestinal disorders, all findings summarized in the Table 2.
Table 2.
Summary of in vitro and in vivo studies on the effects of Za’atar bioactive compounds on gastrointestinal disorders and gut microbiota.
3.2.1. Origanum syriacum and Thymbra spicata Extracts
O. syriacum and T. spicata are considered ancient herbal remedies are known as biblical hyssop [109]. O. syriacum showed protective effects in an animal peptic ulcer model with a significant decrease in ulcer number and size, similar to that of clinically approved lansoprazole in the prophylactic model [69]. The antiparasitic potential of O. syriacum essential oils (EOs) against the nematode Anisakis simplex has been demonstrated by Lopez et al. [68]. It must be noted that patients infected with Anisakis simplex developed allergic responses and increased intestinal permeability [110,111]. The EOs obtained from O. syriacum demonstrated potent antifungal activity against several species of fungi potentially involved in intestinal infection and disease [112]. Similarly to O. syriacum, several studies have evaluated the composition and antimicrobial and antiparasitic effects of EOs derived from T. spicata [37,113,114,115].
3.2.2. Carvacrol, Thymol, and Essential Oils (EOs)
Over the past two decades, increasing evidence suggests that EOs derived from aromatic plants could exert a positive effect on gut microbiota and associated intestinal disorders. These oils exert selective antimicrobial effects against pathogenic bacteria, fungi, and parasites, and favorable effects on commensal and beneficial bacteria [116,117,118]. Supplementation with EOs rich in carvacrol and thymol in weaned piglets resulted in beneficial changes in gut microbiota composition demonstrated by an increase in the relative abundance of some beneficial species such as Bacilli, Lactobacillales, and Veillonellaceae [70]. Similarly, supplementation with oregano essential oil (OEO) rich in carvacrol and thymol resulted in changes in the gut microbiota accompanied by improved piglet health and performance [71]. OEO supplementation during lactation showed an increase in the relative abundance of Lactobacillaceae, as well as Fibrobacteriaceae and Akkermansiaceae involved in fiber digestion. At two and four weeks, a relative decrease in Enterobacteriaceae and an increase in butyrate-producing Lachnospiraceae was observed. The authors suggested that the beneficial effects of EOs containing mainly CVL were exerted through the modulation of gut microbiota. [71]. Ceppa et al. [119] observed that EOs mixture of oregano and thyme had a modulatory effect on gut microbiota in rainbow trout (Oncorhynchus mykiss). In a mouse model of antibiotic-associated gut dysbiosis and Clostridium difficile infection, CVL supplementation significantly reduced the incidence of diarrhea and improved gut dysbiosis. CVL had a positive effect on microbiome composition by enhancing the abundance of beneficial bacteria such, as Firmicutes, and reducing the proportion of harmful bacteria such as Proteobacteria [75]. These effects could also be attributed to the direct antibacterial properties of CVL against pathogenic bacteria. Infection with Clostridium perfringens can cause necrotic enteritis. In a C. perfringens infected broiler chicken model, EO supplementation (25% thymol, 25% carvacrol) alleviated gut lesions and enhanced serum antibody titers against the Newcastle disease virus. In the ileum, EOs increased the expression of occludin mRNA and inhibited TLR2 and TNF-α expression [76]. Additionally, supplementation with EOs resulted in changes in the host ileal microbial population—an increase in Lactobacillus crispatus and Lactobacillus agilis accompanied by a decrease in Lactobacillus salivarius and Lactobacillus johnsonii [77]. In a duck model, supplementation with EOs primarily containing thymol and carvacrol decreased microbial populations of coliforms, total aerobes, and lactose-negative Enterobacteria [72]. In addition to the direct immunostimulatory effects of EOs containing thymol and carvacrol demonstrated in a Juvenile hybrid tilapia animal model [78], EOs exerted an indirect effect through changes mediated by the microbiota. This is backed up by the observation that the treatment of germ-free zebrafish colonized by microbiota with EOs exerted a suppressive effect on levels of inflammatory markers such as serum amyloid and interleukin 1β, 8 and upregulated the expression of claudin-1 and occludin-2, two important tight junction proteins for intestinal permeability and integrity [78]. In a model using human intestinal epithelial cells Caco-2, CVL treatment resulted in a significant reduction of C. jejuni adhesion, invasion, and translocation to Caco-2 cells, as well as a reduction in C. jejuni motility, toxin production, and pathogenic gene expression [79]. Mooyotto et al. showed that CVL reduced C. difficile toxin production using an in vitro bacterial culture method and reduced C. difficile cytotoxicity using Vero cells [120] CVL was also able to reduce the invasive ability of Salmonella typhimurium in intestinal epithelial cells IPEC-J2 and inhibit bacterial viability and motility in vitro [121]. Additionally, CVL increased the relative abundance of Lactobacillus spp. in the chicken gut [80]. In a murine infection model of acute campylobacteriosis, CVL reduced disease symptoms by reducing intestinal apoptosis and pro-inflammatory immune responses not only in the intestine, but also in extra-intestinal organs such as the liver, as indicated by decreased serum levels of IFN-γ, TNF, MCP-1 and IL-6 [81]. Michiels et al., showed that CVL and thymol improved gut health in weaning piglets by reducing the number of intra-epithelial lymphocytes and concurrently increasing the villus height to crypt depth ratio in the distal small intestine [122]. Moreover, supplementation with carvacrol and thymol in weaning pigs reduced the intestinal oxidative stress caused by weaning, increased the Lactobacillus population, and decreased the number of Enterococcus and E. coli in the jejunum along with a significant decrease in the expression of TNF-α [83]. Another study showed that the supplementation of EOs containing CVL and thymol in pigs resulted in increased nitrogen digestibility and decreased emission of ammonia and total fecal nitrogen. Additionally, microbial protease and urease activities were inhibited by EO supplementation [123]. Recently, an integrative study investigating the transcriptional response and microbiota modulation in the intestine in response to a diet enriched with carvacrol and thymol essential oils was conducted using a gilthead seabream (sparus aurata) animal model [124]. Several remarkable changes in inflammation and the expression of immunity genes were observed. The activation of the NF-kB pathway, which could impair the maintenance of intestinal epithelial integrity and immune homeostasis, was inhibited. The fish supplemented with EOs also had downregulated expression of NF-kB mediated proinflammatory cytokines such as interleukin-1 beta (il-1β) [124]. Using chicken and mice models, CVL supplementation prevented Campylobacter (a common foodborne pathogen) infection and colonization by reducing the expression of a vast number of genes involved in motility, adhesion, growth, metabolism, and anaerobic respiration [82]. In a model using human intestinal epithelial cells Caco-2, CVL treatment resulted in a significant reduction of C. jejuni adhesion, invasion, and translocation to Caco-2 cells, as well as a reduction in C. jejuni motility, toxin production and pathogenic gene expression [79]. In another cellular study, Mooyotto et al., showed that CVL reduced C. difficile toxin production using an in vitro bacterial culture method and reduced C. difficile cytotoxicity using Vero cells [120] CVL was also able to reduce the invasive ability of Salmonella typhimurium in intestinal epithelial cells IPEC-J2 and inhibit bacterial viability and motility in vitro [121].
3.2.3. Rosmarinic Acid
Rosmarinic acid (RA) is a phenolic acid commonly found in the Lamiaceae plant species. RA is the most abundant phenolic acid present in O. syriacum and T. spicata. Alongside antioxidant and anti-inflammatory effects in several diseases, RA demonstrated a protective effect within in vitro and in vivo models of gastric mucosal injury [73]. RA exhibited cytoprotective, antioxidant, anti-apoptotic, and wound healing properties in the gastric mucosal epithelial cell line RGM-1 insulted with 5% ethanol. The results were further validated in vivo with ethanol-induced gastric mucosal lesions in mice, where pre-treatment with extract rich in RA promoted gastric mucosal healing by decreasing oxidative stress, inflammatory response, proapoptotic protein expression, and gastric mucosal damage. In addition, the group treated with extract rich in RA showed higher microbial diversity when compared to the control group. In particular, an increase in Muribaculaceae and Ruminococcaceae and a decrease in Prevotellaceae was observed in mice treated with RA-rich extract. Additionally, levels of SCFAs such as acetic acid and propionic acid were slightly increased, and butyric acid was notably increased in RA-treated mice compared to control mice [73]. In another study, the prebiotic effects of RA on gut microbiota were demonstrated by an increase in the population of diabetes-resistant bacteria and a decrease in diabetes-sensitive bacteria in diabetic rats [74].
3.2.4. Oleanolic and Ursolic Acid
Oleanolic acid (OA) and ursolic acid (UA) are pentacyclic terpenoids with pharmacologic properties found in olive oil and medicinal herbs such as O. syracum [125]. OA and UA show antibacterial and antiparasitic activity in the gastrointestinal tract [126]. OA and UA inhibited E. coli enterotoxin-induced diarrhea in mice by blocking the binding of enterotoxins to the surface of intestinal epithelial cells [127]. Oral administration of OA in a mice model of colitis significantly inhibited colon shortening and myeloperoxidase activity, displayed potent anti-inflammatory properties by reducing the activation of TNF-α, IL-1β, IL-17, NF-κB, and MAPK, and increasing the expression of IL-10. The study also showed that OA protected intestinal integrity by increasing the expression of tight junction proteins ZO-1, occludin, and claudin-1 in the colon [84]. Similarly, UA exerted anti-inflammatory and antioxidant effects in mice with ulcerative colitis [85,86], and regulated intestinal microbiota composition and inflammatory cell infiltration [87]. UA also displayed protective effects on the intestinal mucosal barrier by attenuating intestinal injury an ileal epithelial cells apoptosis, decreasing LPS, procalcitonin, and intestinal malondialdehyde levels, and increasing the expression of the tight junction proteins claudin 1 and occludin in the ileum of rats [128]. In a mice model of liver fibrosis, UA reduced intestinal damage by inhibiting TNF-α, increasing the expression of tight junction proteins ZO-1 and occludin, as well as intestinal antimicrobial peptide angiogenin-1 to protect the gut barrier. Considerable effects of UA were observed in an antibiotic-resistant mice model. Treatment with UA protected the intestinal barrier by increasing the height of jejunal villi, decreasing jejunal crypt depth, and upregulating the expression of tight junction proteins ZO-1, claudin-1 and occludin. Additionally, UA decreased serum LPS and diamine oxidase levels and downregulated the expression of pro-inflammatory TNF-α and IL-6 cytokines. Supplementation with UA had beneficial effects on gut microbiota composition, resulting in a decrease in Firmicutes and Ruminococcaceae, an increase in Bacteroidetes, and enhanced the growth of SCFA-producing bacteria including the Rikenellaceae and Bifidobacteriaceae families [88]. Additionally, UA treatment improved gut dysbiosis by increasing Lactobacillus and Bifidobacterium populations [89,90].
UA modified the gut microbiota by promoting the growth of Lactobacillus, inhibiting the proliferation of harmful bacteria such as Burkholderiales, Alphaproteobacteria, Betaproteobacteria, and Gammaproteobacteria, as well as downregulating the expression of antibiotic resistance genes [91].
OA displayed a direct effect on intestinal tight junctions and inflammation in a mice model with Salmonella typhimurium-induced diarrhea. OA alleviated intestinal damage and maintained gut barrier integrity by enhancing the expression and localization of occludin, claudin-1, and ZO-1. OA displayed its anti-inflammatory potential by reducing the levels of standard inflammatory markers such as COX-2, iNOS, pro-inflammatory cytokines IL-1β, IL-6, and TNF-α, as well as the physphorylation and degradation of IκB, nuclear translocation of p65, TLR4, and the activation of the MAPK pathway [92]. OA prevented gut atrophy induced by parenteral nutrition in pigs. OA is an agonist of the bile acid-activated G protein-coupled receptor TGR5. The activation of TGR5 by OA ameliorated gut atrophy [93]. Recently, the role of OA in the gut-liver axis was investigated by Xue et al. in HFD rats with interesting results. In addition to combatting obesity and hepatic steatosis, OA supplementation attenuated HFD-induced metabolic endotoxemia and intestinal barrier damage by decreasing LPS and pro-inflammatory cytokines. OA also displayed an intestinal protective effect by increasing the expression of intestinal tight junction proteins ZO-1 and occludin, decreasing serum diamine oxidase (DAO) activity and d-lactate concentrations, and reducing intestinal inflammation by inhibiting the TLR4/NF-κB pathway [94]. The authors found that OA could modulate gut microbiota and improve gut immunity by enhancing microbial diversity, reducing the ratio of Firmicutes to Bacteroidetes, and increasing the abundance of butyrate-producing bacteria [94,95]. In addition, oleanolic acid glycosides (3-O-glycoside moiety) accelerated gastrointestinal transit and prevented ileus in mice [129]. UA-modulated sphingomyelinase (SMase) activity, is associated with several inflammatory diseases and tumors including inflammatory bowel disease and colon cancer [130,131].
3.2.5. Rhus coriaria (Sumac)
Plants belonging to the Rhus genus have been commonly used in traditional medicine to treat gastrointestinal disorders [27,51]. Several Rhus species that share various phytochemicals with R. coriaria have demonstrated beneficial effects in intestinal disorders in animal and cellular models of gastrointestinal inflammation [132,133], diarrhea [134,135], ulcerative colitis [136], and Vibrio vulnificus infection [137]. In streptozotocin-induced diabetic rats with intestinal oxidative damage, supplementation with sumac extract enhanced the antioxidant defense system by increasing the level of GSH, GST, GR, CAT, GPx, and SOD in the small intestine, and decreasing MDA levels and α-glucosidase activity [97]. In a necrotizing enterocolitis model using newborn rat pups, Isik et al. evaluated the protective effects of sumac supplementation on intestinal injury. Sumac ameliorated histopathologic and biochemical markers in the ileum and proximal colon, and also showed anti-apoptotic and anti-inflammatory effects in the rat colon. Additionally, sumac reduced oxidative stress by reducing lipid peroxidation and DNA and protein oxidation [98]. The inhibitory effect of sumac on the enzyme urease was tested in vitro. Urease is essential for the colonization of the gastric mucosa by Helicobacter pylori, a bacterium that induces gastrointestinal diseases such as gastritis and peptic ulcer disease, potentially leading to gastric cancer. Sumac showed a potent inhibitory effect against Jack bean urease activity [138]. The effects of sumac on microbiota were evaluated by using a metabolomic approach in an in vitro model of gut microbiota. Farag et al. investigated the response of the gut microbiota to sumac by analyzing changes in metabolites in bacterial cultures. Overall, a decrease in amino acid levels, nitrogenous compounds, and sugar levels was observed in samples treated with sumac. These changes were accompanied by an increase in levels of SCFA and nucleic acids [96].
Gallic acid (GA), the most abundant phenolic compound in sumac, and plant extract rich in gallic acid showed potential prebiotic effects associated with a decrease in intestinal inflammation and the promotion of intestinal integrity [139]. A recent study reported that GA may have a protective effect against colon toxicity. Colon toxicity induced by 1,2-dimethylhydrazine in female Waster rats was characterized by increased inflammation, apoptosis, oxidative stress, goblet cell disintegration, and mucin depletion. Treatment with GA reduced all these parameters and increased glutathione content and the activity of the detoxifying enzymes GPx, GR, GST, and CAT [101]. Several studies have reported that GA exerted inhibitory effects on intestinal pathogenic bacteria, such as Clostridium histolyticum, Clostridium difficile, and Bacteroides spp., which have been associated with several intestinal diseases [140]. Penta-O-galloyl-β-D-glucose—a gallotannin present in the Rhus plant family and sumac-inhibited NF-κB and MAPK activation in stimulated peritoneal and colonic macrophages and suppressed IL-1β, TNF-α, and IL-6 in LPS-stimulated peritoneal macrophages, while increasing the expression of the anti-inflammatory cytokine IL-10 in vitro. In a mice model of colitis, supplementation with penta-O-galloyl-β-D-glucose inhibited colon shortening and myeloperoxidase activity, reduced the activation of NF-κB and levels of IL-1β, TNF-α, and IL-6, but increased IL-10 levels [99].
In addition, GA showed a protective effect against ulcerative colitis mediated by gut microbiota g modulation in the rat. Treatment with GA increased the growth of probiotic bacteria, such as Lactobacillaceae and Prevotellaceae, and decreased the number of several pathogenic species, such as Firmicutes and Proteobacteria families. GA also induced metabolic changes by increasing carbohydrate bile acid metabolism and decreasing amino acid metabolism [100]. A mixture of GA and anthocyanins, which are compounds abundant in sumac, significantly enhanced the growth of Bifidobacterium spp. and Lactobacillus spp. [141]. Anthocyanins are primarily transformed into GA by the gut microbiota, contributing to their anti-inflammatory effects [142].
To summarize, the potential bioactivity of GA, GA-rich extracts, and GA metabolites from gallotannins and anthocyanins against inflammatory disorders and intestinal disease, as well as their potential for microbiota modulation, have been recently extensively reviewed [139,143,144]. The evidence available so far suggests that along with its direct antioxidant and anti-inflammatory effects, GA may modulate the gastrointestinal immune system by modifying the composition of gut microbiota.
3.2.6. Sesamum indicum (Sesame)
Sesame sauce displayed anti-inflammatory and anti-cancerogenic effects in a mouse colon carcinogenesis model. Specifically, supplementation with sesame sauce decreased the expression and serum levels of tumor necrosis factor-α, interferon-γ, interleukin (IL)-6, IL-17α, inducible nitric oxide synthase, and cyclooxygenase-2 in mice colon mucosa [102]. Sesamol, a lignan present in sesame oil with antioxidant and anti-inflammatory properties, suppressed cyclooxygenase-2 transcriptional activity in colon cancer cells and modified the development of intestinal polyps in mice [145]. Sesamol protected gut barrier integrity and reduced the release of LPS in aging mice and HFD models. Treatment with sesamol increased the length of intestinal villi and muscularis mucosa thickness and elevated mRNA expression of tight junction complex claudin-1 in the colon. Moreover, sesamol was able to attenuate HDF-induced inflammation in the colon by reducing the expression of IL-1β and TNF-α [103,104]. Sesaminol glucosides alleviated premalignant lesions of the rat colon [146]. These findings indicate that sesame may possess chemopreventive properties.
Cecal ligation and puncture is a well-established model for abdominal sepsis. In septic rats and mice sesame oil significantly decreased markers of oxidative stress such as lipid peroxidation and serum nitrite levels, consequently attenuating hepatic injury. Moreover, supplementation with sesame oil increased the survival rate and levels of anti-inflammatory cytokine IL-10, as well as significantly reduced xanthine oxidase activity [147,148,149,150]. Rezaeipour et al. observed that diets containing sesame meal increased villus height and villus height to crypt depth ratio in the jejunum [151]. In a rat model of chemically-induced acute colitis, sesame oil displayed potential healing effects as shown by a significant decrease in the levels of inflammatory markers (mast cells and CD68+ cells), fibrosis (collagen, laminin), and modulation of colon mucins by a decrease in acidic mucin and increase in neutral mucin [106]. Similar results were also observed in colitic mice, with a fermented sauce containing sesame displaying significant protective and anti-inflammatory effects in the colon [152]. In a rat model of acetic acid-induced inflammatory bowel disease, the level of myeloperoxidase, lipid peroxidation, and nitrite in the colon was reduced following sesamol treatment [107]. Sesamol showed a protective effect against radiation-induced gastrointestinal injury in mice, with pre-treatment with a single dose of sesamol partially decreasing radiation-induced mortality. Additionally, pre-treatment with sesamol decreased lipid peroxidation and the translocation of gut bacteria to the spleen and liver and enhanced the regeneration of crypt cells in the gastrointestinal tract [108]. The effect of sesame constituents on gut microbiota is well observed. A significant increase in microbiome diversity was observed in the group supplemented with sesamol, with higher numbers of Bifidobacterium and Akkermansia and lower numbers of Clostridium bacteria, when compared to aging mice [104]. In HFD mice, sesamol significantly improved the relative abundance of Bacillales, Fusobacterium, and Lactococus, while decreasing the number of Bilophila. In addition, sesamol significantly increased the level of SCFAs such as acetate, propionate, and butyrate [103]. Likewise, supplementation with sesamol in an Alzheimer mice model altered gut microbiota by significantly decreasing the relative abundance of Helicobacter hepaticus and Clostridium spp. and increasing the relative abundance of Rikenellaceae and Bifidobacterium [105]. The authors showed that sesamol also has a protective effect on gut barrier integrity, decreasing LPS leakage into the serum and increasing levels of SCFAs, including acetate, propionate, isobutyrate, butyrate, and valerate [105]. These findings indicate that sesamol can decrease systemic inflammation and improve brain disease by protecting the integrity of the gut barrier, altering the gut microbiota, reducing LPS levels, and increasing the level of SCFAs in the blood.
3.3. Clinical Studies (Table 3)
The effect of a diet rich in polyphenols on increased intestinal permeability was investigated by measuring serum zonulin levels in a randomized, controlled, cross-over clinical trial [153]. Polyphenol intake significantly decreased serum zonulin levels and blood pressure. This effect was accompanied by a significant increase in fiber-fermenting and butyrate-producing bacteria such as Ruminococcaceae and Faecalibacterium. Notably, the effect of a polyphenol-rich diet was greater in subjects with metabolic syndrome, suggesting an association between intestinal permeability and metabolic syndrome [153]. The polyphenol-rich diet used in the study shares a similar profile with the polyphenolic compounds present in the Za’atar mixture. Za’atar contains high concentrations of polyphenols and ursolic and oleanolic acids, which have also been associated with the improvement of several components of metabolic syndrome in humans such as the reduction of inflammatory cytokines, plasma lipid and cholesterol levels [154,155]. Gallotannins and gallic acid-rich extract (which are also found in high amounts in sumac and Za’atar mixture) improved chronic constipation, increased levels of gastrin and valeric acid, and decreased levels of endotoxins and interleukin 6 [156]. After consuming Gallotannins and gallic acid-rich mango extract for six weeks, lean and obese individuals had increased levels of tannase-producing Lactococcus lactis and decreased levels of Clostridium leptum and Bacteroides thetaiotaomicron, which have been shown to be associated with obesity. In addition, an increase in fecal SCFAs, such as butyric and valeric acid, and a decrease in endotoxin levels were observed after the consumption of polyphenols [157]. Polyphenols have also demonstrated anti-inflammatory and gut microbiota-modifying effects in IBD patients. Gallotannins and gallic acid-rich mango pulp intake (200–400 g) for eight weeks reduced plasma levels of pro-inflammatory cytokines such as interleukin 8 (IL-8), growth-regulated oncogene and granulocyte-macrophage colony-stimulating factor. The consumption of polyphenols modulates the composition of gut microbiota by increasing the abundance of Lactobacillus plantarum, Lactobacillus reuteri, and Lactobacillus lactis, accompanied by increased fecal butyric acid production [139].
A randomized, placebo-controlled, crossover study showed that sesame consumption improved blood lipids and exerted antioxidant effects [158]. In addition, in subjects with partial adhesive small bowel obstruction, treatment with sesame oil had a positive effect on clinical parameters including a decreased need for surgical intervention, and a shorter convalescence and hospital stay compared to the control group [159].
Table 3.
Clinical studies assessing the effects of a polyphenol-rich diet on gut microbiota and intestinal permeability in gastrointestinal disorders.
Table 3.
Clinical studies assessing the effects of a polyphenol-rich diet on gut microbiota and intestinal permeability in gastrointestinal disorders.
| Authors (Year) | Sample Size | Gender M/F (Age) | Participants | Format, Dose | Duration of Study | Main Findings |
|---|---|---|---|---|---|---|
| [153] | 51 | 27/39 (≥60 years) | Subjects with increased IP | PR-diet 724 mg/day | 8 weeks | ↓ Serum zonulin ↓ Blood pressure ↑ Fiber-fermenting bacteria, butyrate-producing bacteria |
| [156] | 36 | 11/25 (8–65 years) | Subjects with chronic constipation | Mango consumption, 300 g/day | 4 weeks | ↑ Stool frequency, consistency ↑ Gastrin levels, SCFA (valeric acid) ↓ Endotoxin and IL-6 |
| [157] | 32 | 22/10 (18–50 years) | healthy lean and obese individuals | Mango consumption, 400 g/day | 6 weeks | ↑ SCFA in lean individuals |
| [139] | 10 | 5/5 (18–75 years) | IBD Subjects | Mango pulp intake, 200–400 g/day | 8 weeks | ↑ SCCAI ↓ IL-8 ↓ GRO and GM-CSF ↑ Fecal butyric acid ↓ Inflammation biomarkers ↑ Lactobacillus plantarum, Lactobacillus reuteri, Lactobacillus lactis |
| [159] | 64 | 33/31 (19–79) | Subjects with symptoms and signs of SBO | Sesame oil in nasogastric tube, 150 mL/day | 1 day | ↓ SBO resolution time ↓ Hospital stays ↓ Relaparotomy rate |
Abbreviations: PR-diet—polyphenol-rich diet; IP—intestinal permeability; SCFAs—short-chain fatty acids; IL—interleukin; GRO—growth-regulated oncogene, GM-CSF—colony-stimulating factor; SBO—small bowel obstruction; SSCAI—squamous cell carcinoma antigen-1; IBD—Inflammatory bowel diseases; ↑—increased; ↓—decreased.
4. Summary
A growing number of studies have reported potential gut microbiota-mediated effects of plant-based functional foods rich in polyphenols on human health and disease. Moreover, plant-derived polyphenols have attracted much attention in regard to the regulation of intestinal barrier function [160]. In humans, the bioactivation of polyphenols happens through intestinal transformation and is mediated partly by digestive enzymes, but fundamentally by gut microbiota. Once the active “smaller” phenolic compounds and metabolites are absorbed into the portal vein, they travel to various tissues and organs to enact their beneficial role [161]. However, polyphenols also exert local cytoprotective and antioxidant effects, especially on intestinal cells. The plant-based Za’atar mixture is widely used as a food and ingredient in Lebanon and the Eastern part of the Mediterranean basin. The mixture is rich in dietary fiber and phenolic compounds, as well as minerals and vitamins. Za’atar mixture is mainly a blend of O. syriacum (known as the Za’atar plant in Lebanon) and Thymbra spicata (wild Za’atar), R. coriaria (sumac), and sesame. Despite its richness in bioactive compounds and its nutraceutical potential, studies on the whole Za’atar mixture are still missing, however, most of the studies have focused on the single components or single (poly)phenolic compounds. Here we reviewed the literature available on chemical and phytochemical profile of Za’atar, and its combined effects on the gastrointestinal tract.
Studies have shown that even at low doses the components present in Za’atar are able to prevent and reduce chemically induced acute inflammation in animal models. In addition, these compounds have exhibited protective and therapeutic effects in pathogen and chemical-induced colitis. Furthermore, polyphenols can attenuate the mucosal activation of NF-κB, the production of cytokines, intestinal barrier dysfunction, and apoptosis (Table 2).
Gut dysbiosis is closely related to increased intestinal permeability. Therefore, the modulation of gut microbiota can have a positive effect on intestinal permeability and vice versa. Diet can play a modulatory role, although the exact molecular mechanisms involved in this interplay remain unclear, especially in humans [162].
As shown in Table 2 and Table 3, several experimental studies in animals and humans have reported that Za’atar components may exert prebiotic-like effects on gut microbiota by enhancing the growth of beneficial bacteria (e.g., Lactobacillus spp., Bifidobacterium spp., fiber-fermenting, and butyrate-producing bacteria) and reducing the relative number of potentially harmful and pathogenic bacteria (e.g., Clostridium spp.). Additionally, Za’atar components may be beneficial in the treatment of intestinal inflammatory diseases and leaky gut syndrome. Numerous studies have reported the potential antioxidant and anti-inflammatory effects of Za’atar compounds in experimental models of intestinal disease in animals and humans (Table 2 and Table 3). Based on a review of the available studies, we show that Za’atar compounds may significantly reduce intestinal permeability by increasing the level of tight junction proteins and by maintaining gut epithelial integrity. The overall possible effects of Za’atar on gastrointestinal tract are depicted in the Figure 3.
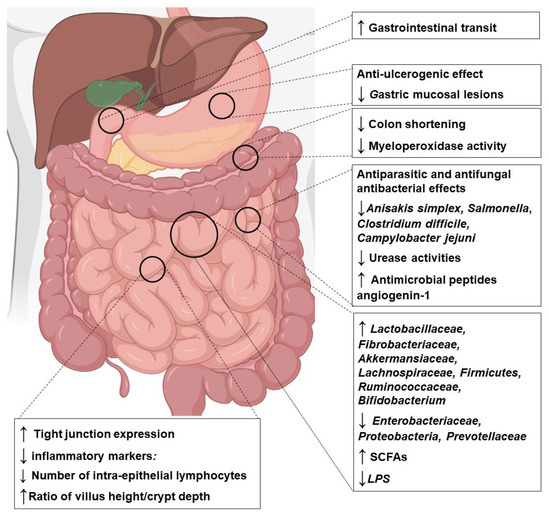
Figure 3.
Summary of the combined effects of polyphenols found in Za’atar components on the gastrointestinal tract. Abbreviations: SCFA—short-chain fatty acids; LPS—lipopolysaccharide; ↑—increased; ↓—decreased. The figure is developed using https://biorender.com/ (accessed on 12 December 2022).
5. Clinical Perspectives and Future Applications
The use of natural compounds in the prevention and treatment of disease has garnered increasing clinical and medical interest over the past decades.
The idea of natural compounds is appealing due to their ease of use as dietary supplements, lower associated costs, and a more favorable side effect profile when compared to synthetic drugs. Another attractive aspect is the potential use of natural compounds in the treatment of functional gastrointestinal disorders (e.g., dyspepsia, irritable bowel syndrome, constipation) as an adjunct to dietary intervention and lifestyle modifications. In fact, individuals affected by functional gastrointestinal disorders often seek out alternative methods to improve their symptoms when the limited number of currently available guideline-based treatment options have failed or provided insufficient improvement of symptoms [163]. Factors implicated in functional gastrointestinal disorders include gastrointestinal dysmotility, altered intestinal barrier and immune function, dysbiosis, dysregulation of neurologic and signaling networks, and stress [164]. The effects of Za’atar components reported so far are related to several of these factors, raising questions as to whether Za’atar and its components might have beneficial clinical effects.
Although the potential clinical applications of natural compounds such as Za’atar are attractive, research on this mixture and its components is still in its early stages before evidence-based conclusions and recommendations can be made. We cover the advantages and limitations of Za’atar use in the following paragraphs.
Advantages: Za’atar is a low-cost and environmentally friendly plant-based food consisting of locally available herbs. Active compounds found in Za’atar have gained attention due to their potential role in the prevention and treatment of cardiovascular, metabolic, and gastrointestinal diseases with no clinically significant side effects reported in studies undertaken so far. Based on our previous discussion on the properties of its compounds, we raise the following questions on whether Za’atar could be applied as: (i) a prebiotic-like agent, (ii) an adjunct to dietary modifications used in the prevention or treatment of metabolic and gastrointestinal diseases; (iii) a supplement and nutraceutical.
Limitations: Despite the health-promoting effects of the components of Za’atar described previously, an understanding of the biological effects of the Za’atar mixture as a whole is missing. More animal and human studies are necessary to further elucidate the translational aspects of the effects of Za’atar and to fully characterize the mixture as a preventive and therapeutic tool to ascertain its viability as a plant-based functional food. In fact, there is no defined therapeutic dose for the Za’atar mixture, which could partially explain why there is a lack of clinical trials.
Suggestions concerning and covering the general quality and safety aspects of traditional herbal medicine or food products are depicted in the following points:
- Well-designed human studies evaluating drug interaction with herbal supplements are required.
- Controlled clinical trials evaluating the acceptance, tolerance, and safety of herbal preparation are highly needed.
- Standardized preparations of herbal products or traditional foods should be adjusted to an exact content of substances with well-known nutraceutical properties.
Regarding the possible effects on the gastrointestinal tract, future clinical trials investigating Za’atar mixture should consider the following: (i) consider the interaction of Za’atar with gut microbiota and levels of short-chain fatty acids (SCFAs); (ii) take into account the effect of Za’atar on intestinal barrier function and selectivity by assessing levels of oxidative stress, inflammatory markers, and tight junction proteins; (iii) interpret the effects observed in the context of dietary factors as well as an adjunct to the appropriate dietary intervention.
In this context, our future molecular and clinical studies on gastrointestinal motility, intestinal permeability, and microbiota composition and metabolites will address the effects of the Za’atar mixture whilst testing its clinical efficacy in a selected group of subjects, with the aim of evaluating the potential health-promoting effects of Za’atar.
6. Conclusions
Several studies on cellular and animal models, as well as in humans, have demonstrated the beneficial effects of individual plant-based bioactive compounds on the gastrointestinal tract. The combination of well-known medicinal and health-promoting herbs in Za’atar could provide synergistic pharmacological and nutraceutical effects, especially pertaining to the gastrointestinal tract and gut microbiota. In conclusion, a Za’atar mixture rich in polyphenols, essential oils, and fiber could provide beneficial effects in the modulation of gut microbiota and intestinal permeability. Its richness in bioactive compounds, safety, and prebiotic properties make mixed Za’atar a potential promising adjunct to existing treatments used in gastrointestinal and metabolic diseases. However, studies of the effects of the whole Za’atar mixture are required to explore the combined effects and safety of Za’atar, in addition, well-controlled human studies are necessary to improve our understanding of the molecular mechanisms and effects on microbiota and intestinal permeability before any conclusions can be made on the clinical application of Za’atar.
Author Contributions
M.K.: Methodology, Writing—Original Draft; H.A.: Data curation; D.R.-E.: Methodology; M.C. and M.D.A.: Data Analysis; P.P.: Project administration, Supervision, Funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by FOIE GRAS project, which has received funding from the European Union’s Horizon 2020 Research and Innovation framework, under the Marie Skłodowska-Curie Grant Agreement No. 722619 (P.P.) and the mitoFOIE GRAS funded by H2020-MSCA-RISE-2016/H2020-MSCA-RISE-2016 Grant Agreement No. 734719 (P.P.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No data were generated for this publication.
Acknowledgments
The authors are indebted to Carlo Di Benedetta and Marina Ricci for helpful discussion involving the Consortium of Mediterranean Universities (CMU), Bari (www.cmungo.eu).
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Portincasa, P.; Bonfrate, L.; Khalil, M.; Angelis, M.D.; Calabrese, F.M.; D’Amato, M.; Wang, D.Q.-H.; Di Ciaula, A. Intestinal Barrier and Permeability in Health, Obesity and NAFLD. Biomedicines 2022, 10, 83. [Google Scholar] [CrossRef]
- Scaldaferri, F.; Pizzoferrato, M.; Gerardi, V.; Lopetuso, L.; Gasbarrini, A. The gut barrier: New acquisitions and therapeutic approaches. J. Clin. Gastroenterol. 2012, 46, S12–S17. [Google Scholar] [CrossRef]
- Konturek, P.C.; Brzozowski, T.; Konturek, S.J. Stress and the gut: Pathophysiology, clinical consequences, diagnostic approach and treatment options. J. Physiol. Pharmacol. 2011, 62, 591–599. [Google Scholar]
- Portincasa, P.; Bonfrate, L.; Vacca, M.; De Angelis, M.; Farella, I.; Lanza, E.; Khalil, M.; Wang, D.Q.-H.; Sperandio, M.; Di Ciaula, A. Gut Microbiota and Short Chain Fatty Acids: Implications in Glucose Homeostasis. Int. J. Mol. Sci. 2022, 23, 1105. [Google Scholar] [CrossRef]
- Kallus, S.J.; Brandt, L.J. The intestinal microbiota and obesity. J. Clin. Gastroenterol. 2012, 46, 16–24. [Google Scholar] [CrossRef]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; FitzGerald, M.G.; Fulton, R.S.; et al. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Guarner, F.; Malagelada, J.R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Biesalski, H.K. Nutrition meets the microbiome: Micronutrients and the microbiota. Ann. N. Y. Acad. Sci. 2016, 1372, 53–64. [Google Scholar] [CrossRef]
- Gao, K.; Mu, C.-L.; Farzi, A.; Zhu, W.-Y. Tryptophan Metabolism: A Link Between the Gut Microbiota and Brain. Adv. Nutr. 2019, 11, 709–723. [Google Scholar] [CrossRef]
- De Angelis, M.; Garruti, G.; Minervini, F.; Bonfrate, L.; Portincasa, P.; Gobbetti, M. The Food-gut Human Axis: The Effects of Diet on Gut Microbiota and Metabolome. Curr. Med. Chem. 2019, 26, 3567–3583. [Google Scholar] [CrossRef]
- France, M.M.; Turner, J.R. The mucosal barrier at a glance. J. Cell Sci. 2017, 130, 307–314. [Google Scholar] [CrossRef]
- Salvo Romero, E.; Alonso Cotoner, C.; Pardo Camacho, C.; Casado Bedmar, M.; Vicario, M. The intestinal barrier function and its involvement in digestive disease. Rev. Esp. Enferm. Dig. 2015, 107, 686–696. [Google Scholar] [CrossRef]
- Camilleri, M.; Madsen, K.; Spiller, R.; Van Meerveld, B.G.; Verne, G.N. Intestinal barrier function in health and gastrointestinal disease. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2012, 24, 503–512. [Google Scholar] [CrossRef]
- Blikslager, A.T.; Moeser, A.J.; Gookin, J.L.; Jones, S.L.; Odle, J. Restoration of Barrier Function in Injured Intestinal Mucosa. Physiol. Rev. 2007, 87, 545–564. [Google Scholar] [CrossRef]
- Sonnenburg, J.L.; Backhed, F. Diet-microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef]
- Belancic, A. Gut microbiome dysbiosis and endotoxemia—Additional pathophysiological explanation for increased COVID-19 severity in obesity. Obes. Med. 2020, 20, 100302. [Google Scholar] [CrossRef]
- Salguero, M.V.; Al-Obaide, M.A.I.; Singh, R.; Siepmann, T.; Vasylyeva, T.L. Dysbiosis of Gram-negative gut microbiota and the associated serum lipopolysaccharide exacerbates inflammation in type 2 diabetic patients with chronic kidney disease. Exp. Ther. Med. 2019, 18, 3461–3469. [Google Scholar] [CrossRef]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef]
- Kimera, F.; Sewilam, H.; Fouad, W.M.; Suloma, A. Sustainable production of Origanum syriacum L. using fish effluents improved plant growth, yield, and essential oil composition. Heliyon 2021, 7, e06423. [Google Scholar] [CrossRef]
- Alwafa, R.A.; Mudalal, S.; Shraim, F.; Mauriello, G. Comparison between Quality Traits of Solar-Dried and Freeze-Dried Origanum syriacum L. (Za’atar). Plants 2022, 11, 1110. [Google Scholar] [CrossRef]
- Mudalal, S.; Kanan, D.; Anabtawi, O.; Irshaid, A.; Sabbah, M.; Shtaya, M.; Shraim, F.; Mauriello, G. Application of the Hurdle Technology Concept to the Fresh Za’atar (Origanum syriacum) Preservation. Foods 2022, 11, 3002. [Google Scholar] [CrossRef]
- Alwafa, R.A.; Mudalal, S.; Mauriello, G. Origanum syriacum L. (Za’atar), from Raw to Go: A Review. Plants 2021, 10, 1001. [Google Scholar] [CrossRef]
- Khalil, M.; Caponio, G.R.; Diab, F.; Shanmugam, H.; Di Ciaula, A.; Khalifeh, H.; Vergani, L.; Calasso, M.; De Angelis, M.; Portincasa, P. Unraveling the beneficial effects of herbal Lebanese mixture “Za’atar”. History, studies, and properties of a potential healthy food ingredient. J. Funct. Foods 2022, 90, 104993. [Google Scholar] [CrossRef]
- Khalil, M.; Khalifeh, H.; Baldini, F.; Serale, N.; Parodi, A.; Voci, A.; Vergani, L.; Daher, A. Antitumor Activity of Ethanolic Extract from Thymbra spicata L. aerial Parts: Effects on Cell Viability and Proliferation, Apoptosis Induction, STAT3, and NF-kB Signaling. Nutr. Cancer 2020, 73, 1193–1206. [Google Scholar] [CrossRef]
- Khoury, M.; Stien, D.; Eparvier, V.; Ouaini, N.; El Beyrouthy, M. Report on the Medicinal Use of Eleven Lamiaceae Species in Lebanon and Rationalization of Their Antimicrobial Potential by Examination of the Chemical Composition and Antimicrobial Activity of Their Essential Oils. Evid. Based Complement. Altern. Med. 2016, 2016, 2547169. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Petrelli, R.; Cappellacci, L.; Bartolucci, F.; Canale, A.; Maggi, F. Origanum syriacum subsp. syriacum: From an ingredient of Lebanese ‘manoushe’ to a source of effective and eco-friendly botanical insecticides. Ind. Crop. Prod. 2019, 134, 26–32. [Google Scholar] [CrossRef]
- Khalil, M.; Hayek, S.; Khalil, N.; Serale, N.; Vergani, L.; Calasso, M.; De Angelis, M.; Portincasa, P. Role of Sumac (Rhus coriaria L.) in the management of metabolic syndrome and related disorders: Focus on NAFLD-atherosclerosis interplay. J. Funct. Foods 2021, 87, 104811. [Google Scholar] [CrossRef]
- Toker, E.B.; Yeşilbağ, K. In vitro antiviral activity of Thymbra spicata L. extract on bovine respiratory viruses (BCoV, BPIV-3, BRSV, BVDV and BoHV-1). J. Appl. Microbiol. 2022, 132, 2625–2632. [Google Scholar] [CrossRef]
- Mesmar, J.; Abdallah, R.; Badran, A.; Maresca, M.; Baydoun, E. Origanum syriacum Phytochemistry and Pharmacological Properties: A Comprehensive Review. Molecules 2022, 27, 4272. [Google Scholar] [CrossRef]
- AlKahlout, A.; Fardoun, M.; Mesmar, J.; Abdallah, R.; Badran, A.; Nasser, S.A.; Baydoun, S.; Kobeissy, F.; Shaito, A.; Iratni, R.; et al. Origanum syriacum L. Attenuates the Malignant Phenotype of MDA-MB231 Breast Cancer Cells. Front. Oncol. 2022, 12, 922196. [Google Scholar] [CrossRef]
- Kerem, S.; Koşar, N.; Tekin, F.; Güreser, A.S.; Özbek, Ö. Investigation of antimicrobial activities and molecular characterization of the species belong to Origanum, Thymus and Thymbra genera by ISSR. Mol. Biol. Rep. 2023, 50, 289–298. [Google Scholar] [CrossRef]
- Diab, F.; Khalil, M.; Lupidi, G.; Zbeeb, H.; Salis, A.; Damonte, G.; Bramucci, M.; Portincasa, P.; Vergani, L. Influence of Simulated In Vitro Gastrointestinal Digestion on the Phenolic Profile, Antioxidant, and Biological Activity of Thymbra spicata L. Extracts. Antioxidants 2022, 11, 1778. [Google Scholar] [CrossRef]
- Alonazi, M.A.; Jemel, I.; Moubayed, N.; Alwhibi, M.; El-Sayed, N.N.E.; Ben Bacha, A. Evaluation of the in vitro anti-inflammatory and cytotoxic potential of ethanolic and aqueous extracts of Origanum syriacum and Salvia lanigera leaves. Environ. Sci. Pollut. Res. Int. 2021, 28, 19890–19900. [Google Scholar] [CrossRef]
- Sedeek, M.S.; Afifi, S.M.; Mansour, M.K.; Hassan, M.; Mehaya, F.M.; Naguib, I.A.; Abourehab, M.A.S.; Farag, M.A. Unveiling Antimicrobial and Antioxidant Compositional Differences between Dukkah and Za’atar via SPME-GCMS and HPLC-DAD. Molecules 2022, 27, 6471. [Google Scholar] [CrossRef]
- Shen, D.; Pan, M.H.; Wu, Q.L.; Park, C.H.; Juliani, H.R.; Ho, C.T.; Simon, J.E. LC-MS method for the simultaneous quantitation of the anti-inflammatory constituents in oregano (Origanum species). J. Agric. Food Chem. 2010, 58, 7119–7125. [Google Scholar] [CrossRef]
- Al Hafi, M.; El Beyrouthy, M.; Ouaini, N.; Stien, D.; Rutledge, D.; Chaillou, S. Chemical Composition and Antimicrobial Activity of Satureja, Thymus, and Thymbra Species Grown in Lebanon. Chem. Biodivers. 2017, 14, e1600236. [Google Scholar] [CrossRef]
- Gedikoğlu, A.; Sökmen, M.; Çivit, A. Evaluation of Thymus vulgaris and Thymbra spicata essential oils and plant extracts for chemical composition, antioxidant, and antimicrobial properties. Food Sci. Nutr. 2019, 7, 1704–1714. [Google Scholar] [CrossRef]
- Dorman, H.J.; Bachmayer, O.; Kosar, M.; Hiltunen, R. Antioxidant properties of aqueous extracts from selected lamiaceae species grown in Turkey. J. Agric. Food Chem. 2004, 52, 762–770. [Google Scholar] [CrossRef]
- Khalil, M.; Khalifeh, H.; Baldini, F.; Salis, A.; Damonte, G.; Daher, A.; Voci, A.; Vergani, L. Antisteatotic and antioxidant activities of Thymbra spicata L. extracts in hepatic and endothelial cells as in vitro models of non-alcoholic fatty liver disease. J. Ethnopharmacol. 2019, 239, 111919. [Google Scholar] [CrossRef]
- Prasad Mn, N.; Kr, S.; Prasad, D.S. A Review on Nutritional and Nutraceutical Properties of Sesame. J. Nutr. Food Sci. 2012, 2, 1–6. [Google Scholar] [CrossRef]
- Elleuch, M.; Besbes, S.; Roiseux, O.; Blecker, C.; Attia, H. Quality characteristics of sesame seeds and by-products. Food Chem. 2007, 103, 641–650. [Google Scholar] [CrossRef]
- Saydut, A.; Duz, M.; Kaya, C.; Kafadar, A.; Hamamci, C. Transesterified sesame (Sesamum indicum L.) seed oil as a biodiesel fuel. Bioresour. Technol. 2008, 99, 6656–6660. [Google Scholar] [CrossRef]
- Majdalawieh, A.F.; Mansour, Z.R. Sesamol, a major lignan in sesame seeds (Sesamum indicum): Anti-cancer properties and mechanisms of action. Eur. J. Pharmacol. 2019, 855, 75–89. [Google Scholar] [CrossRef]
- Regazzoni, L.; Arlandini, E.; Garzon, D.; Santagati, N.A.; Beretta, G.; Facino, R.M. A rapid profiling of gallotannins and flavonoids of the aqueous extract of Rhus coriaria L. by flow injection analysis with high-resolution mass spectrometry assisted with database searching. J. Pharm. Biomed. Anal. 2013, 72, 202–207. [Google Scholar] [CrossRef]
- Gök, H.N.; Deliorman Orhan, D.; Gürbüz, İ.; Aslan, M. Activity-guided isolation of α-amylase, α-glucosidase, and pancreatic lipase inhibitory compounds from Rhus coriaria L. J. Food Sci. 2020, 85, 3220–3228. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Ali-Shtayeh, M.S.; Jamous, R.M.; Arráez-Román, D.; Segura-Carretero, A. HPLC-DAD-ESI-MS/MS screening of bioactive components from Rhus coriaria L. (Sumac) fruits. Food Chem. 2015, 166, 179–191. [Google Scholar] [CrossRef]
- Mehrdad, M.; Zebardast, M.; Abedi, G.; Koupaei, M.N.; Rasouli, H.; Talebi, M. Validated high-throughput HPLC method for the analysis of flavonol aglycones myricetin, quercetin, and kaempferol in Rhus coriaria L. using a monolithic column. J. AOAC Int. 2009, 92, 1035–1043. [Google Scholar]
- Hashim, A.M.; Alharbi, B.M.; Abdulmajeed, A.M.; Elkelish, A.; Hozzein, W.N.; Hassan, H.M. Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content. Plants 2020, 9, 869. [Google Scholar] [CrossRef]
- Shamseddine, L.; Chidiac, J.J. Composition’s effect of Origanum syriacum essential oils in the antimicrobial activities for the treatment of denture stomatitis. Odontology 2021, 109, 327–335. [Google Scholar] [CrossRef]
- Al Hafi, M.; El Beyrouthy, M.; Ouaini, N.; Stien, D.; Rutledge, D.; Chaillou, S. Chemical Composition and Antimicrobial Activity of Origanum libanoticum, Origanum ehrenbergii, and Origanum syriacum Growing Wild in Lebanon. Chem. Biodivers. 2016, 13, 555–560. [Google Scholar] [CrossRef]
- Khalil, M.; Bazzi, A.; Zeineddine, D.; Jomaa, W.; Daher, A.; Awada, R. Repressive effect of Rhus coriaria L. fruit extracts on microglial cells-mediated inflammatory and oxidative stress responses. J. Ethnopharmacol. 2021, 269, 113748. [Google Scholar] [CrossRef]
- Sakhr, K.; El Khatib, S. Physiochemical properties and medicinal, nutritional and industrial applications of Lebanese Sumac (Syrian Sumac—Rhus coriaria): A review. Heliyon 2020, 6, e03207. [Google Scholar] [CrossRef]
- Kizil, S.; Turk, M. Microelement contents and fatty acid compositions of Rhus coriaria L. and Pistacia terebinthus L. fruits spread commonly in the south eastern Anatolia region of Turkey. Nat. Prod. Res. 2010, 24, 92–98. [Google Scholar] [CrossRef]
- Reidel, R.V.B.; Cioni, P.L.; Majo, L.; Pistelli, L. Evolution of Volatile Emission in Rhus coriaria Organs During Different Stages of Growth and Evaluation of the Essential Oil Composition. Chem. Biodivers. 2017, 14, e1700270. [Google Scholar] [CrossRef]
- Namiki, M. Nutraceutical functions of sesame: A review. Crit. Rev. Food Sci. Nutr. 2007, 47, 651–673. [Google Scholar] [CrossRef]
- Kermani, S.G.; Saeidi, G.; Sabzalian, M.R.; Gianinetti, A. Drought stress influenced sesamin and sesamolin content and polyphenolic components in sesame (Sesamum indicum L.) populations with contrasting seed coat colors. Food Chem. 2019, 289, 360–368. [Google Scholar] [CrossRef]
- Esmaeilzadeh Kenari, R.; Mohsenzadeh, F.; Amiri, Z.R. Antioxidant activity and total phenolic compounds of Dezful sesame cake extracts obtained by classical and ultrasound-assisted extraction methods. Food Sci. Nutr. 2014, 2, 426–435. [Google Scholar] [CrossRef]
- Makarewicz, M.; Drożdż, I.; Tarko, T.; Duda-Chodak, A. The Interactions between Polyphenols and Microorganisms, Especially Gut Microbiota. Antioxidants 2021, 10, 188. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Co, V.A.; El-Nezami, H. Dietary polyphenol impact on gut health and microbiota. Crit. Rev. Food Sci. Nutr. 2021, 61, 690–711. [Google Scholar] [CrossRef]
- Burdulis, D.; Sarkinas, A.; Jasutiené, I.; Stackevicené, E.; Nikolajevas, L.; Janulis, V. Comparative study of anthocyanin composition, antimicrobial and antioxidant activity in bilberry (Vaccinium myrtillus L.) and blueberry (Vaccinium corymbosum L.) fruits. Acta Pol. Pharm. 2009, 66, 399–408. [Google Scholar]
- Sourabh, A.; Kanwar, S.S.; Sud, R.G.; Ghabru, A.; Sharma, O.P. Influence of phenolic compounds of Kangra tea [Camellia sinensis (L) O Kuntze] on bacterial pathogens and indigenous bacterial probiotics of Western Himalayas. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2013, 44, 709–715. [Google Scholar] [CrossRef]
- Chan, E.W.; Soh, E.Y.; Tie, P.P.; Law, Y.P. Antioxidant and antibacterial properties of green, black, and herbal teas of Camellia sinensis. Pharmacogn. Res. 2011, 3, 266–272. [Google Scholar] [CrossRef]
- Shen, X.; Sun, X.; Xie, Q.; Liu, H.; Zhao, Y.; Pan, Y.; Hwang, C.-A.; Wu, V.C.H. Antimicrobial effect of blueberry (Vaccinium corymbosum L.) extracts against the growth of Listeria monocytogenes and Salmonella enteritidis. Food Control 2014, 35, 159–165. [Google Scholar] [CrossRef]
- Vitetta, L.; Vitetta, G.; Hall, S. Immunological Tolerance and Function: Associations Between Intestinal Bacteria, Probiotics, Prebiotics, and Phages. Front. Immunol. 2018, 9, 2240. [Google Scholar] [CrossRef]
- Larrosa, M.; Luceri, C.; Vivoli, E.; Pagliuca, C.; Lodovici, M.; Moneti, G.; Dolara, P. Polyphenol metabolites from colonic microbiota exert anti-inflammatory activity on different inflammation models. Mol. Nutr. Food Res. 2009, 53, 1044–1054. [Google Scholar] [CrossRef]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Prebiotic evaluation of cocoa-derived flavanols in healthy humans by using a randomized, controlled, double-blind, crossover intervention study. Am. J. Clin. Nutr. 2010, 93, 62–72. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Sánchez-Alcoholado, L.; Pérez-Martínez, P.; Andrés-Lacueva, C.; Cardona, F.; Tinahones, F.; Queipo-Ortuño, M.I. Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients. Food Funct. 2016, 7, 1775–1787. [Google Scholar] [CrossRef]
- López, V.; Pavela, R.; Gómez-Rincón, C.; Les, F.; Bartolucci, F.; Galiffa, V.; Petrelli, R.; Cappellacci, L.; Maggi, F.; Canale, A.; et al. Efficacy of Origanum syriacum Essential Oil against the Mosquito Vector Culex quinquefasciatus and the Gastrointestinal Parasite Anisakis simplex, with Insights on Acetylcholinesterase Inhibition. Molecules 2019, 24, 2563. [Google Scholar] [CrossRef]
- El-Meligy, R.M.; Awaad, A.S.; Soliman, G.A.; Kenawy, S.A.; Alqasoumi, S.I. Prophylactic and curative anti-ulcerogenic activity and the possible mechanisms of action of some desert plants. Saudi Pharm. J. 2017, 25, 387–396. [Google Scholar] [CrossRef]
- Li, Y.; Fu, X.; Ma, X.; Geng, S.; Jiang, X.; Huang, Q.; Hu, C.; Han, X. Intestinal Microbiome-Metabolome Responses to Essential Oils in Piglets. Front. Microbiol. 2018, 9, 1988. [Google Scholar] [CrossRef]
- Hall, H.N.; Wilkinson, D.J.; Le Bon, M. Oregano essential oil improves piglet health and performance through maternal feeding and is associated with changes in the gut microbiota. Anim. Microbiome 2021, 3, 2. [Google Scholar] [CrossRef]
- Abouelezz, K.; Abou-Hadied, M.; Yuan, J.; Elokil, A.A.; Wang, G.; Wang, S.; Wang, J.; Bian, G. Nutritional impacts of dietary oregano and Enviva essential oils on the performance, gut microbiota and blood biochemicals of growing ducks. Animal 2019, 13, 2216–2222. [Google Scholar] [CrossRef]
- Wang, G.Y.; Chen, S.Y.; Chen, Y.Y.; Hong, C.J.; Hsu, Y.H.; Yen, G.C. Protective effect of rosmarinic acid-rich trichodesma khasianum clarke leaves against ethanol-induced gastric mucosal injury in vitro and in vivo. Phytomedicine 2021, 80, 153382. [Google Scholar] [CrossRef]
- Ou, J.; Huang, J.; Zhao, D.; Du, B.; Wang, M. Protective effect of rosmarinic acid and carnosic acid against streptozotocin-induced oxidation, glycation, inflammation and microbiota imbalance in diabetic rats. Food Funct. 2018, 9, 851–860. [Google Scholar] [CrossRef]
- Mooyottu, S.; Flock, G.; Upadhyay, A.; Upadhyaya, I.; Maas, K.; Venkitanarayanan, K. Protective Effect of Carvacrol against Gut Dysbiosis and Clostridium difficile Associated Disease in a Mouse Model. Front. Microbiol. 2017, 8, 625. [Google Scholar] [CrossRef]
- Du, E.; Wang, W.; Gan, L.; Li, Z.; Guo, S.; Guo, Y. Effects of thymol and carvacrol supplementation on intestinal integrity and immune responses of broiler chickens challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2016, 7, 19. [Google Scholar] [CrossRef]
- Yin, D.; Du, E.; Yuan, J.; Gao, J.; Wang, Y.; Aggrey, S.E.; Guo, Y. Supplemental thymol and carvacrol increases ileum Lactobacillus population and reduces effect of necrotic enteritis caused by Clostridium perfringes in chickens. Sci. Rep. 2017, 7, 7334. [Google Scholar] [CrossRef]
- Ran, C.; Hu, J.; Liu, W.; Liu, Z.; He, S.; Dan, B.C.; Diem, N.N.; Ooi, E.L.; Zhou, Z. Thymol and Carvacrol Affect Hybrid Tilapia through the Combination of Direct Stimulation and an Intestinal Microbiota-Mediated Effect: Insights from a Germ-Free Zebrafish Model. J. Nutr. 2016, 146, 1132–1140. [Google Scholar] [CrossRef]
- Upadhyay, A.; Arsi, K.; Wagle, B.R.; Upadhyaya, I.; Shrestha, S.; Donoghue, A.M.; Donoghue, D.J. Trans-Cinnamaldehyde, Carvacrol, and Eugenol Reduce Campylobacter jejuni Colonization Factors and Expression of Virulence Genes in Vitro. Front. Microbiol. 2017, 8, 713. [Google Scholar] [CrossRef]
- Kelly, C.; Gundogdu, O.; Pircalabioru, G.; Cean, A.; Scates, P.; Linton, M.; Pinkerton, L.; Magowan, E.; Stef, L.; Simiz, E.; et al. The In Vitro and In Vivo Effect of Carvacrol in Preventing Campylobacter Infection, Colonization and in Improving Productivity of Chicken Broilers. Foodborne Pathog. Dis. 2017, 14, 341–349. [Google Scholar] [CrossRef]
- Mousavi, S.; Schmidt, A.M.; Escher, U.; Kittler, S.; Kehrenberg, C.; Thunhorst, E.; Bereswill, S.; Heimesaat, M.M. Carvacrol ameliorates acute campylobacteriosis in a clinical murine infection model. Gut Pathog. 2020, 12, 2. [Google Scholar] [CrossRef]
- Wagle, B.R.; Donoghue, A.M.; Shrestha, S.; Upadhyaya, I.; Arsi, K.; Gupta, A.; Liyanage, R.; Rath, N.C.; Donoghue, D.J.; Upadhyay, A. Carvacrol attenuates Campylobacter jejuni colonization factors and proteome critical for persistence in the chicken gut. Poult. Sci. 2020, 99, 4566–4577. [Google Scholar] [CrossRef]
- Wei, H.K.; Xue, H.X.; Zhou, Z.X.; Peng, J. A carvacrol-thymol blend decreased intestinal oxidative stress and influenced selected microbes without changing the messenger RNA levels of tight junction proteins in jejunal mucosa of weaning piglets. Animal 2017, 11, 193–201. [Google Scholar] [CrossRef]
- Kang, G.D.; Lim, S.; Kim, D.H. Oleanolic acid ameliorates dextran sodium sulfate-induced colitis in mice by restoring the balance of Th17/Treg cells and inhibiting NF-κB signaling pathway. Int. Immunopharmacol. 2015, 29, 393–400. [Google Scholar] [CrossRef]
- Chun, J.; Lee, C.; Hwang, S.W.; Im, J.P.; Kim, J.S. Ursolic acid inhibits nuclear factor-κB signaling in intestinal epithelial cells and macrophages, and attenuates experimental colitis in mice. Life Sci. 2014, 110, 23–34. [Google Scholar] [CrossRef]
- Liu, B.; Piao, X.; Guo, L.; Liu, S.; Chai, F.; Gao, L. Ursolic acid protects against ulcerative colitis via anti-inflammatory and antioxidant effects in mice. Mol. Med. Rep. 2016, 13, 4779–4785. [Google Scholar] [CrossRef]
- Sheng, Q.; Li, F.; Chen, G.; Li, J.; Li, J.; Wang, Y.; Lu, Y.; Li, Q.; Li, M.; Chai, K. Ursolic Acid Regulates Intestinal Microbiota and Inflammatory Cell Infiltration to Prevent Ulcerative Colitis. J. Immunol. Res. 2021, 2021, 6679316. [Google Scholar] [CrossRef]
- Hao, W.; Kwek, E.; He, Z.; Zhu, H.; Liu, J.; Zhao, Y.; Ma, K.Y.; He, W.S.; Chen, Z.Y. Ursolic acid alleviates hypercholesterolemia and modulates the gut microbiota in hamsters. Food Funct. 2020, 11, 6091–6103. [Google Scholar] [CrossRef]
- Wan, S.Z.; Liu, C.; Huang, C.K.; Luo, F.Y.; Zhu, X. Ursolic Acid Improves Intestinal Damage and Bacterial Dysbiosis in Liver Fibrosis Mice. Front. Pharmacol. 2019, 10, 1321. [Google Scholar] [CrossRef]
- Wan, S.; Huang, C.; Wang, A.; Zhu, X. Ursolic acid improves the bacterial community mapping of the intestinal tract in liver fibrosis mice. PeerJ 2020, 8, e9050. [Google Scholar] [CrossRef]
- Peng, F.; Zhang, H.; He, X.; Song, Z. Effects of Ursolic Acid on Intestinal Health and Gut Bacteria Antibiotic Resistance in Mice. Front. Physiol. 2021, 12, 650190. [Google Scholar] [CrossRef]
- Dong, N.; Xue, C.; Zhang, L.; Zhang, T.; Wang, C.; Bi, C.; Shan, A. Oleanolic acid enhances tight junctions and ameliorates inflammation in Salmonella typhimurium-induced diarrhea in mice via the TLR4/NF-κB and MAPK pathway. Food Funct. 2020, 11, 1122–1132. [Google Scholar] [CrossRef]
- Jain, A.K.; Wen, J.X.; Blomenkamp, K.S.; Arora, S.; Blaufuss, T.A.; Rodrigues, J.; Long, J.P.; Neuschwander-Tetri, B.A.; Teckman, J.H. Oleanolic Acid Improves Gut Atrophy Induced by Parenteral Nutrition. JPEN J. Parenter. Enter. Nutr. 2016, 40, 67–72. [Google Scholar] [CrossRef]
- Xue, C.; Li, Y.; Lv, H.; Zhang, L.; Bi, C.; Dong, N.; Shan, A.; Wang, J. Oleanolic Acid Targets the Gut-Liver Axis to Alleviate Metabolic Disorders and Hepatic Steatosis. J. Agric. Food Chem. 2021, 69, 7884–7897. [Google Scholar] [CrossRef]
- Xue, C.; Lv, H.; Li, Y.; Dong, N.; Wang, Y.; Zhou, J.; Shi, B.; Shan, A. Oleanolic acid reshapes the gut microbiota and alters immune-related gene expression of intestinal epithelial cells. J. Sci. Food Agric. 2021, 102, 764–773. [Google Scholar] [CrossRef]
- Farag, M.A.; Abdelwareth, A.; Sallam, I.E.; El Shorbagi, M.; Jehmlich, N.; Fritz-Wallace, K.; Schape, S.S.; Rolle-Kampczyk, U.; Ehrlich, A.; Wessjohann, L.A.; et al. Metabolomics reveals impact of seven functional foods on metabolic pathways in a gut microbiota model. J. Adv. Res. 2020, 23, 47–59. [Google Scholar] [CrossRef]
- Doğan, A.; Çelik, İ. Healing effects of sumac (Rhus coriaria) in streptozotocin-induced diabetic rats. Pharm. Biol. 2016, 54, 2092–2102. [Google Scholar] [CrossRef]
- Isik, S.; Tayman, C.; Cakir, U.; Koyuncu, I.; Taskin Turkmenoglu, T.; Cakir, E. Sumac (Rhus coriaria) for the prevention and treatment of necrotizing enterocolitis. J. Food Biochem. 2019, 43, e13068. [Google Scholar] [CrossRef]
- Jang, S.E.; Hyam, S.R.; Jeong, J.J.; Han, M.J.; Kim, D.H. Penta-O-galloyl-β-D-glucose ameliorates inflammation by inhibiting MyD88/NF-κB and MyD88/MAPK signalling pathways. Br. J. Pharmacol. 2013, 170, 1078–1091. [Google Scholar] [CrossRef]
- Li, Y.; Xie, Z.; Gao, T.; Li, L.; Chen, Y.; Xiao, D.; Liu, W.; Zou, B.; Lu, B.; Tian, X.; et al. A holistic view of gallic acid-induced attenuation in colitis based on microbiome-metabolomics analysis. Food Funct. 2019, 10, 4046–4061. [Google Scholar] [CrossRef]
- Shree, A.; Islam, J.; Vafa, A.; Mohammad Afzal, S.; Sultana, S. Gallic acid prevents 1, 2-Dimethylhydrazine induced colon inflammation, toxicity, mucin depletion, and goblet cell disintegration. Environ. Toxicol. 2020, 35, 652–664. [Google Scholar] [CrossRef]
- Song, J.L.; Choi, J.H.; Seo, J.H.; Park, K.Y. Fermented Ganjangs (Soy Sauce and Sesame Sauce) Attenuates Colonic Carcinogenesis in Azoxymethane/Dextran Sodium Sulfate-Treated C57BL/6J Mice. J. Med. Food 2018, 21, 905–914. [Google Scholar] [CrossRef]
- Yuan, T.; Chu, C.; Shi, R.; Cui, T.; Zhang, X.; Zhao, Y.; Shi, X.; Hui, Y.; Pan, J.; Qian, R.; et al. ApoE-Dependent Protective Effects of Sesamol on High-Fat Diet-Induced Behavioral Disorders: Regulation of the Microbiome-Gut-Brain Axis. J. Agric. Food Chem. 2019, 67, 6190–6201. [Google Scholar] [CrossRef]
- Ren, B.; Yuan, T.; Zhang, X.; Wang, L.; Pan, J.; Liu, Y.; Zhao, B.; Zhao, W.; Liu, Z.; Liu, X. Protective Effects of Sesamol on Systemic Inflammation and Cognitive Impairment in Aging Mice. J. Agric. Food Chem. 2020, 68, 3099–3111. [Google Scholar] [CrossRef]
- Liu, Q.; Xie, T.; Xi, Y.; Li, L.; Mo, F.; Liu, X.; Liu, Z.; Gao, J.M.; Yuan, T. Sesamol Attenuates Amyloid Peptide Accumulation and Cognitive Deficits in APP/PS1 Mice: The Mediating Role of the Gut-Brain Axis. J. Agric. Food Chem. 2021, 69, 12717–12729. [Google Scholar] [CrossRef]
- Periasamy, S.; Hsu, D.Z.; Chandrasekaran, V.R.; Liu, M.Y. Sesame oil accelerates healing of 2,4,6-trinitrobenzenesulfonic acid-induced acute colitis by attenuating inflammation and fibrosis. JPEN J. Parenter. Enter. Nutr. 2013, 37, 674–682. [Google Scholar] [CrossRef]
- Kondamudi, P.K.; Kovelamudi, H.; Mathew, G.; Nayak, P.G.; Rao, M.C.; Shenoy, R.R. Investigation of sesamol on myeloperoxidase and colon morphology in acetic acid-induced inflammatory bowel disorder in albino rats. Sci. World J. 2014, 2014, 802701. [Google Scholar] [CrossRef]
- Khan, S.; Kumar, A.; Adhikari, J.S.; Rizvi, M.A.; Chaudhury, N.K. Protective effect of sesamol against ⁶⁰Co γ-ray-induced hematopoietic and gastrointestinal injury in C57BL/6 male mice. Free Radic. Res. 2015, 49, 1344–1361. [Google Scholar] [CrossRef]
- Dafni, A.; Böck, B. Medicinal plants of the Bible-revisited. J. Ethnobiol. Ethnomed. 2019, 15, 57. [Google Scholar] [CrossRef]
- Carballeda-Sangiao, N.; Sánchez-Alonso, I.; Navas, A.; Arcos, S.C.; de Palencia, P.F.; Careche, M.; González-Muñoz, M. Anisakis simplex products impair intestinal epithelial barrier function and occludin and zonula occludens-1 localisation in differentiated Caco-2 cells. PLoS Negl. Trop. Dis. 2020, 14, e0008462. [Google Scholar] [CrossRef]
- Polimeno, L.; Loiacono, M.; Pesetti, B.; Mallamaci, R.; Mastrodonato, M.; Azzarone, A.; Annoscia, E.; Gatti, F.; Amoruso, A.; Ventura, M.T. Anisakiasis, an underestimated infection: Effect on intestinal permeability of Anisakis simplex-sensitized patients. Foodborne Pathog. Dis. 2010, 7, 809–814. [Google Scholar] [CrossRef]
- Daouk, R.K.; Dagher, S.M.; Sattout, E.J. Antifungal Activity of the Essential Oil of Origanum syriacum L. J. Food Prot. 1995, 58, 1147–1149. [Google Scholar] [CrossRef]
- Kiliç, T. Analysis of essential oil composition of Thymbra spicata var. spicata: Antifungal, antibacterial and antimycobacterial activities. Z. Nat. C J. Biosci. 2006, 61, 324–328. [Google Scholar] [CrossRef]
- Karimi, M.; Naserifar, R.; Abbasi, N.; Abdi, J.; Maspi, N. In vitro antileishmanial activity of hydroalcoholic Thymbra spicata extract on Leishmania major promastigotes. Ann. Parasitol. 2021, 67, 237–241. [Google Scholar] [CrossRef]
- Haroun, M.F.; Al-Kayali, R.S. Synergistic effect of Thymbra spicata L. extracts with antibiotics against multidrug-resistant Staphylococcus aureus and Klebsiella pneumoniae strains. Iran. J. Basic Med. Sci. 2016, 19, 1193–1200. [Google Scholar]
- Hawrelak, J.A.; Cattley, T.; Myers, S.P. Essential oils in the treatment of intestinal dysbiosis: A preliminary in vitro study. Altern. Med. Rev. 2009, 14, 380–384. [Google Scholar]
- Thapa, D.; Louis, P.; Losa, R.; Zweifel, B.; Wallace, R.J. Essential oils have different effects on human pathogenic and commensal bacteria in mixed faecal fermentations compared with pure cultures. Microbiology 2015, 161, 441–449. [Google Scholar] [CrossRef]
- Logan, A.C.; Beaulne, T.M. The treatment of small intestinal bacterial overgrowth with enteric-coated peppermint oil: A case report. Altern. Med. Rev. 2002, 7, 410–417. [Google Scholar]
- Ceppa, F.; Faccenda, F.; De Filippo, C.; Albanese, D.; Pindo, M.; Martelli, R.; Marconi, P.; Lunelli, F.; Fava, F.; Parisi, G. Influence of essential oils in diet and life-stage on gut microbiota and fillet quality of rainbow trout (Oncorhynchus mykiss). Int. J. Food Sci. Nutr. 2018, 69, 318–333. [Google Scholar] [CrossRef]
- Mooyottu, S.; Kollanoor-Johny, A.; Flock, G.; Bouillaut, L.; Upadhyay, A.; Sonenshein, A.L.; Venkitanarayanan, K. Carvacrol and trans-cinnamaldehyde reduce Clostridium difficile toxin production and cytotoxicity in vitro. Int. J. Mol. Sci. 2014, 15, 4415–4430. [Google Scholar] [CrossRef]
- Burt, S.A.; Adolfse, S.J.; Ahad, D.S.; Tersteeg-Zijderveld, M.H.; Jongerius-Gortemaker, B.G.; Post, J.A.; Brüggemann, H.; Santos, R.R. Cinnamaldehyde, Carvacrol and Organic Acids Affect Gene Expression of Selected Oxidative Stress and Inflammation Markers in IPEC-J2 Cells Exposed to Salmonella typhimurium. Phytother. Res. PTR 2016, 30, 1988–2000. [Google Scholar] [CrossRef]
- Michiels, J.; Missotten, J.; Van Hoorick, A.; Ovyn, A.; Fremaut, D.; De Smet, S.; Dierick, N. Effects of dose and formulation of carvacrol and thymol on bacteria and some functional traits of the gut in piglets after weaning. Arch. Anim. Nutr. 2010, 64, 136–154. [Google Scholar] [CrossRef]
- Hsu, J.E.; Lo, S.H.; Lin, Y.Y.; Wang, H.T.; Chen, C.Y. Effects of essential oil mixtures on nitrogen metabolism and odor emission via in vitro simulated digestion and in vivo growing pig experiments. J. Sci. Food Agric. 2021, 102, 1939–1947. [Google Scholar] [CrossRef]
- Firmino, J.P.; Vallejos-Vidal, E.; Balebona, M.C.; Ramayo-Caldas, Y.; Cerezo, I.M.; Salomón, R.; Tort, L.; Estevez, A.; Moriñigo, M.; Reyes-López, F.E.; et al. Diet, Immunity, and Microbiota Interactions: An Integrative Analysis of the Intestine Transcriptional Response and Microbiota Modulation in Gilthead Seabream (Sparus aurata) Fed an Essential Oils-Based Functional Diet. Front. Immunol. 2021, 12, 625297. [Google Scholar] [CrossRef]
- Liu, J. Oleanolic acid and ursolic acid: Research perspectives. J. Ethnopharmacol. 2005, 100, 92–94. [Google Scholar] [CrossRef]
- Szakiel, A.; Ruszkowski, D.; Grudniak, A.; Kurek, A.; Wolska, K.I.; Doligalska, M.; Janiszowska, W. Antibacterial and antiparasitic activity of oleanolic acid and its glycosides isolated from marigold (Calendula officinalis). Planta Med. 2008, 74, 1709–1715. [Google Scholar] [CrossRef]
- Chen, J.C.; Chang, Y.S.; Wu, S.L.; Chao, D.C.; Chang, C.S.; Li, C.C.; Ho, T.Y.; Hsiang, C.Y. Inhibition of Escherichia coli heat-labile enterotoxin-induced diarrhea by Chaenomeles speciosa. J. Ethnopharmacol. 2007, 113, 233–239. [Google Scholar] [CrossRef]
- Zhang, W.; Gan, D.; Jian, J.; Huang, C.; Luo, F.; Wan, S.; Jiang, M.; Wan, Y.; Wang, A.; Li, B.; et al. Protective Effect of Ursolic Acid on the Intestinal Mucosal Barrier in a Rat Model of Liver Fibrosis. Front. Physiol. 2019, 10, 956. [Google Scholar] [CrossRef]
- Li, Y.; Matsuda, H.; Yoshikawa, M. Effects of oleanolic acid glycosides on gastrointestinal transit and ileus in mice. Bioorg. Med. Chem. 1999, 7, 1201–1205. [Google Scholar] [CrossRef]
- Andersson, D.; Cheng, Y.; Duan, R.D. Ursolic acid inhibits the formation of aberrant crypt foci and affects colonic sphingomyelin hydrolyzing enzymes in azoxymethane-treated rats. J. Cancer Res. Clin. Oncol. 2008, 134, 101–107. [Google Scholar] [CrossRef]
- Zhang, P.; Cheng, Y.; Duan, R.D. Ursolic acid inhibits acid sphingomyelinase in intestinal cells. Phytother. Res. PTR 2013, 27, 173–178. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, H.G.; Im, H.J.; Lee, J.S.; Lee, S.B.; Kim, W.Y.; Lee, H.W.; Lee, S.K.; Byun, C.K.; Son, C.G. Antiemetic and Myeloprotective Effects of Rhus verniciflua Stoke in a Cisplatin-Induced Rat Model. Evid. Based Complement. Altern. Med. 2017, 2017, 9830342. [Google Scholar] [CrossRef]
- Barka, Z.B.; Grintzalis, K.; Polet, M.; Heude, C.; Sommer, U.; Ben Miled, H.; Rhouma, K.B.; Mohsen, S.; Tebourbi, O.; Schneider, Y.J. A combination of NMR and liquid chromatography to characterize the protective effects of Rhus tripartita extracts on ethanol-induced toxicity and inflammation on intestinal cells. J. Pharm. Biomed. Anal. 2018, 150, 347–354. [Google Scholar] [CrossRef]
- Barka, Z.B.; Aouadhi, C.; Tlili, M.; Alimi, H.; Miled, H.B.; Rhouma, K.B.; Sakly, M.; Ksouri, R.; Schneider, Y.J.; Maaroufi, A.; et al. Evaluation of the anti-diarrheal activity of the hydromethanolic root extract of Rhus tripartita (Ucria) (Anacardiacae). Biomed. Pharmacother. 2016, 83, 827–834. [Google Scholar] [CrossRef]
- Tangpu, V.; Yadav, A.K. Antidiarrhoeal activity of Rhus javanica ripen fruit extract in albino mice. Fitoterapia 2004, 75, 39–44. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, O.; Ma, N.; Yi, J.; Mi, H.; Cai, S. The preventive effect and underlying mechanism of Rhus chinensis Mill. fruits on dextran sulphate sodium-induced ulcerative colitis in mice. Food Funct. 2021, 12, 9965–9978. [Google Scholar] [CrossRef]
- Lee, Y.M.; Park, J.P.; Lim, K.T.; Lee, S.J. Intestinal epithelial cell apoptosis due to a hemolytic toxin from Vibrio vulnificus and protection by a 36 kDa glycoprotein from Rhus verniciflua Stokes. Food Chem. Toxicol. 2019, 125, 46–54. [Google Scholar] [CrossRef]
- Mahernia, S.; Bagherzadeh, K.; Mojab, F.; Amanlou, M. Urease Inhibitory Activities of some Commonly Consumed Herbal Medicines. Iran. J. Pharm. Res. 2015, 14, 943–947. [Google Scholar]
- Kim, H.; Venancio, V.P.; Fang, C.; Dupont, A.W.; Talcott, S.T.; Mertens-Talcott, S.U. Mango (Mangifera indica L.) polyphenols reduce IL-8, GRO, and GM-SCF plasma levels and increase Lactobacillus species in a pilot study in patients with inflammatory bowel disease. Nutr. Res. 2020, 75, 85–94. [Google Scholar] [CrossRef]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.; de Pascual-Teresa, S. Metabolism of anthocyanins by human gut microflora and their influence on gut bacterial growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef]
- Hidalgo, M.; Martin-Santamaria, S.; Recio, I.; Sanchez-Moreno, C.; de Pascual-Teresa, B.; Rimbach, G.; de Pascual-Teresa, S. Potential anti-inflammatory, anti-adhesive, anti/estrogenic, and angiotensin-converting enzyme inhibitory activities of anthocyanins and their gut metabolites. Genes Nutr. 2012, 7, 295–306. [Google Scholar] [CrossRef]
- Kiss, A.K.; Piwowarski, J.P. Ellagitannins, Gallotannins and their Metabolites—The Contribution to the Anti-Inflammatory Effect of Food Products and Medicinal Plants. Curr. Med. Chem. 2018, 25, 4946–4967. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, L.; Liao, P.; Xiao, Z.; Zhang, F.; Sindaye, D.; Xin, Z.; Tan, C.; Deng, J.; Yin, Y.; et al. Impact of Gallic Acid on Gut Health: Focus on the Gut Microbiome, Immune Response, and Mechanisms of Action. Front. Immunol. 2020, 11, 580208. [Google Scholar] [CrossRef]
- Shimizu, S.; Fujii, G.; Takahashi, M.; Nakanishi, R.; Komiya, M.; Shimura, M.; Noma, N.; Onuma, W.; Terasaki, M.; Yano, T.; et al. Sesamol suppresses cyclooxygenase-2 transcriptional activity in colon cancer cells and modifies intestinal polyp development in Apc (Min/+) mice. J. Clin. Biochem. Nutr. 2014, 54, 95–101. [Google Scholar] [CrossRef]
- Sheng, H.; Hirose, Y.; Hata, K.; Zheng, Q.; Kuno, T.; Asano, N.; Yamada, Y.; Hara, A.; Osawa, T.; Mori, H. Modifying effect of dietary sesaminol glucosides on the formation of azoxymethane-induced premalignant lesions of rat colon. Cancer Lett. 2007, 246, 63–68. [Google Scholar] [CrossRef]
- Chavali, S.R.; Utsunomiya, T.; Forse, R.A. Increased survival after cecal ligation and puncture in mice consuming diets enriched with sesame seed oil. Crit. Care Med. 2001, 29, 140–143. [Google Scholar] [CrossRef]
- Hsu, D.Z.; Chen, K.T.; Li, Y.H.; Chuang, Y.C.; Liu, M.Y. Sesamol delays mortality and attenuates hepatic injury after cecal ligation and puncture in rats: Role of oxidative stress. Shock 2006, 25, 528–532. [Google Scholar] [CrossRef]
- Hsu, D.Z.; Li, Y.H.; Chien, S.P.; Liu, M.Y. Effects of sesame oil on oxidative stress and hepatic injury after cecal ligation and puncture in rats. Shock 2004, 21, 466–469. [Google Scholar] [CrossRef]
- Hsu, D.Z.; Chien, S.P.; Li, Y.H.; Chuang, Y.C.; Chang, Y.C.; Liu, M.Y. Sesame oil attenuates hepatic lipid peroxidation by inhibiting nitric oxide and superoxide anion generation in septic rats. JPEN J. Parenter. Enter. Nutr. 2008, 32, 154–159. [Google Scholar] [CrossRef]
- Rezaeipour, V.; Barsalani, A.; Abdullahpour, R. Effects of phytase supplementation on growth performance, jejunum morphology, liver health, and serum metabolites of Japanese quails fed sesame (Sesamum indicum) meal-based diets containing graded levels of protein. Trop. Anim. Health Prod. 2016, 48, 1141–1146. [Google Scholar] [CrossRef]
- Song, J.L.; Choi, J.H.; Seo, J.H.; Lim, Y.I.; Park, K.Y. Anti-colitic effects of kanjangs (fermented soy sauce and sesame sauce) in dextran sulfate sodium-induced colitis in mice. J. Med. Food 2014, 17, 1027–1035. [Google Scholar] [CrossRef]
- Del Bo, C.; Bernardi, S.; Cherubini, A.; Porrini, M.; Gargari, G.; Hidalgo-Liberona, N.; González-Domínguez, R.; Zamora-Ros, R.; Peron, G.; Marino, M.; et al. A polyphenol-rich dietary pattern improves intestinal permeability, evaluated as serum zonulin levels, in older subjects: The MaPLE randomised controlled trial. Clin. Nutr. 2021, 40, 3006–3018. [Google Scholar] [CrossRef]
- Luo, H.Q.; Shen, J.; Chen, C.P.; Ma, X.; Lin, C.; Ouyang, Q.; Xuan, C.X.; Liu, J.; Sun, H.B.; Liu, J. Lipid-lowering effects of oleanolic acid in hyperlipidemic patients. Chin. J. Nat. Med. 2018, 16, 339–346. [Google Scholar] [CrossRef]
- Asghari, E.; Rashidlamir, A.; Hosseini, S.R.A.; Moazzami, M.; Samarghandian, S.; Farkhondeh, T. Synergism Effects of Ursolic Acid Supplementation on the Levels of Irisin, C-reactive Protein, IL-6, and TNF-alpha During High-intensity Resistance Training in Low Activity Men. Cardiovasc. Hematol. Disord. Drug Targets 2020, 20, 138–144. [Google Scholar] [CrossRef]
- Venancio, V.P.; Kim, H.; Sirven, M.A.; Tekwe, C.D.; Honvoh, G.; Talcott, S.T.; Mertens-Talcott, S.U. Polyphenol-rich Mango (Mangifera indica L.) Ameliorate Functional Constipation Symptoms in Humans beyond Equivalent Amount of Fiber. Mol. Nutr. Food Res. 2018, 62, e1701034. [Google Scholar] [CrossRef]
- Barnes, R.C.; Kim, H.; Fang, C.; Bennett, W.; Nemec, M.; Sirven, M.A.; Suchodolski, J.S.; Deutz, N.; Britton, R.A.; Mertens-Talcott, S.U.; et al. Body Mass Index as a Determinant of Systemic Exposure to Gallotannin Metabolites during 6-Week Consumption of Mango (Mangifera indica L.) and Modulation of Intestinal Microbiota in Lean and Obese Individuals. Mol. Nutr. Food Res. 2019, 63, e1800512. [Google Scholar] [CrossRef]
- Wu, W.H.; Kang, Y.P.; Wang, N.H.; Jou, H.J.; Wang, T.A. Sesame ingestion affects sex hormones, antioxidant status, and blood lipids in postmenopausal women. J. Nutr. 2006, 136, 1270–1275. [Google Scholar] [CrossRef]
- Ji, Z.L.; Li, J.S.; Yuan, C.W.; Chen, W.D.; Zhang, Y.N.; Ju, X.T.; Tang, W.H. Therapeutic value of sesame oil in the treatment of adhesive small bowel obstruction. Am. J. Surg. 2010, 199, 160–165. [Google Scholar] [CrossRef]
- Yuan, M.; Chen, X.; Su, T.; Zhou, Y.; Sun, X. Supplementation of Kiwifruit Polyphenol Extract Attenuates High Fat Diet Induced Intestinal Barrier Damage and Inflammation via Reshaping Gut Microbiome. Front. Nutr. 2021, 8, 702157. [Google Scholar] [CrossRef]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: Antimicrobial properties. BioMed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef]
- Peron, G.; Hidalgo-Liberona, N.; González-Domínguez, R.; Garcia-Aloy, M.; Guglielmetti, S.; Bernardi, S.; Kirkup, B.; Kroon, P.A.; Cherubini, A.; Riso, P.; et al. Exploring the Molecular Pathways Behind the Effects of Nutrients and Dietary Polyphenols on Gut Microbiota and Intestinal Permeability: A Perspective on the Potential of Metabolomics and Future Clinical Applications. J. Agric. Food Chem. 2020, 68, 1780–1789. [Google Scholar] [CrossRef]
- Mullin, G.E.; Singh, M.; Parian, A.; Weil, A.T. Integrative Gastroenterology; Oxford University Press: Oxford, UK, 2020. [Google Scholar]
- Drossman, D.A.; Hasler, W.L. Rome IV-Functional GI Disorders: Disorders of Gut-Brain Interaction. Gastroenterology 2016, 150, 1257–1261. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).