

Effect of Dietary Supplemented with Mulberry Leaf Powder on Growth Performance, Serum Metabolites, Antioxidant Property and Intestinal Health of Weaned Piglets

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Mulberry Leaf Powder Product
2.2. Diets and Animal Desgin
2.3. Performance and Diarrhea Rate
2.4. Slaughtering and Collecting
2.5. Serological Analysis and Enzyme Activities
2.6. Intestinal Morphometry
2.7. Real-Time PCR
2.8. 16s rRNA Sequencing
2.9. Statistical Analysis
3. Results
3.1. Performance
3.2. Serum Indicators
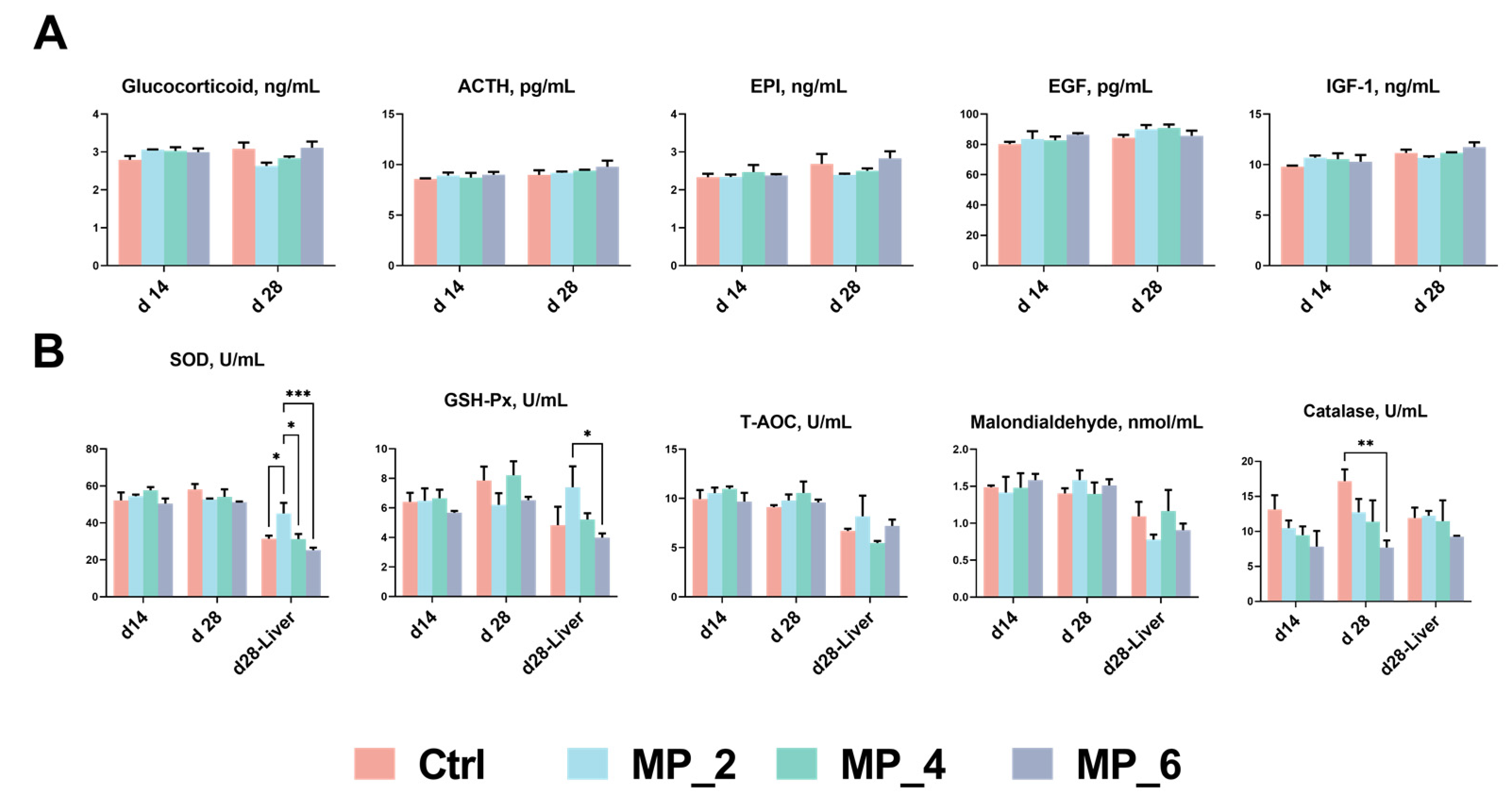
3.3. Serum Hormones and Antioxidant Characteristics
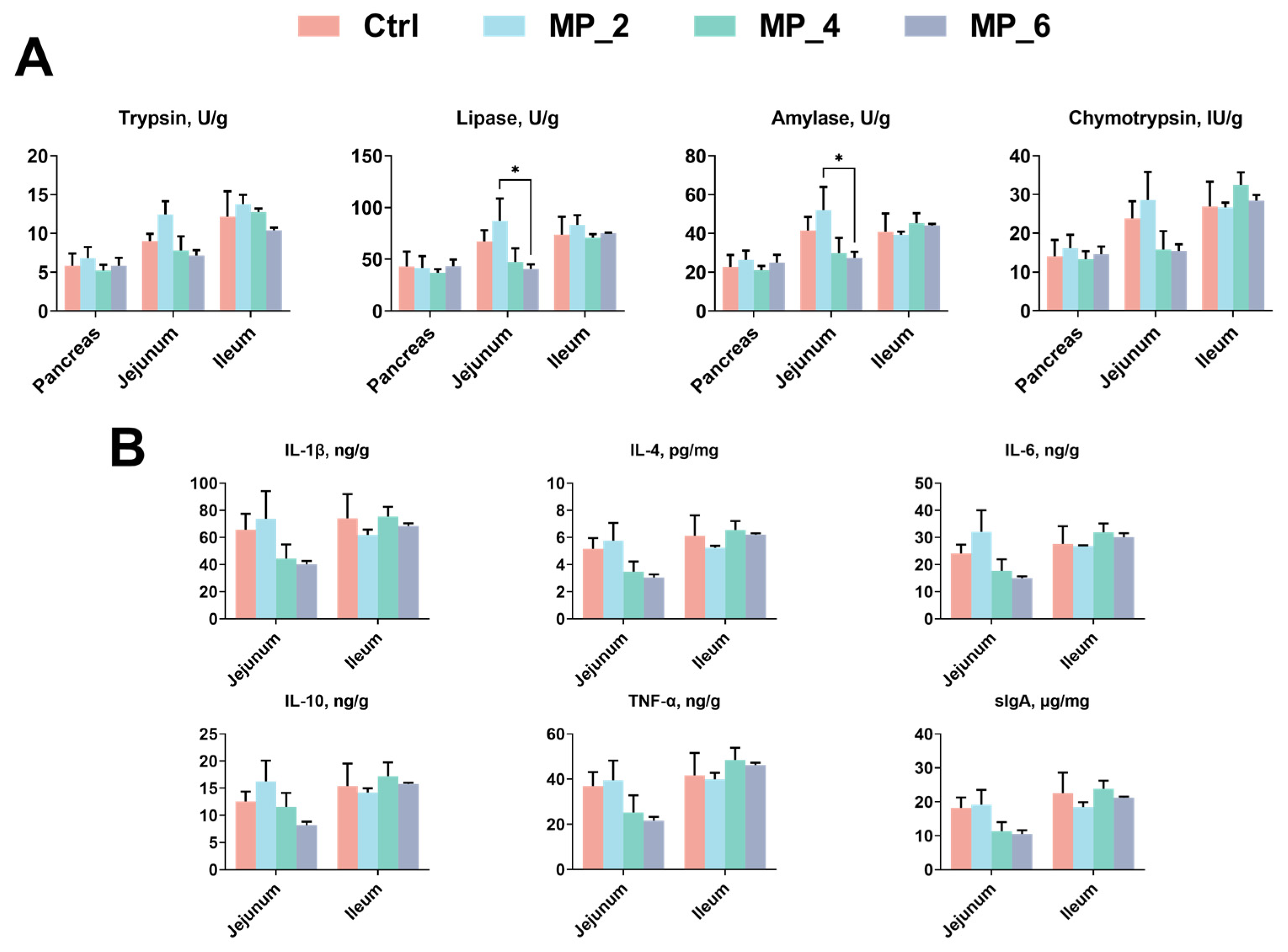
3.4. Digestive Enzyme Activity and Immunity of Intestine
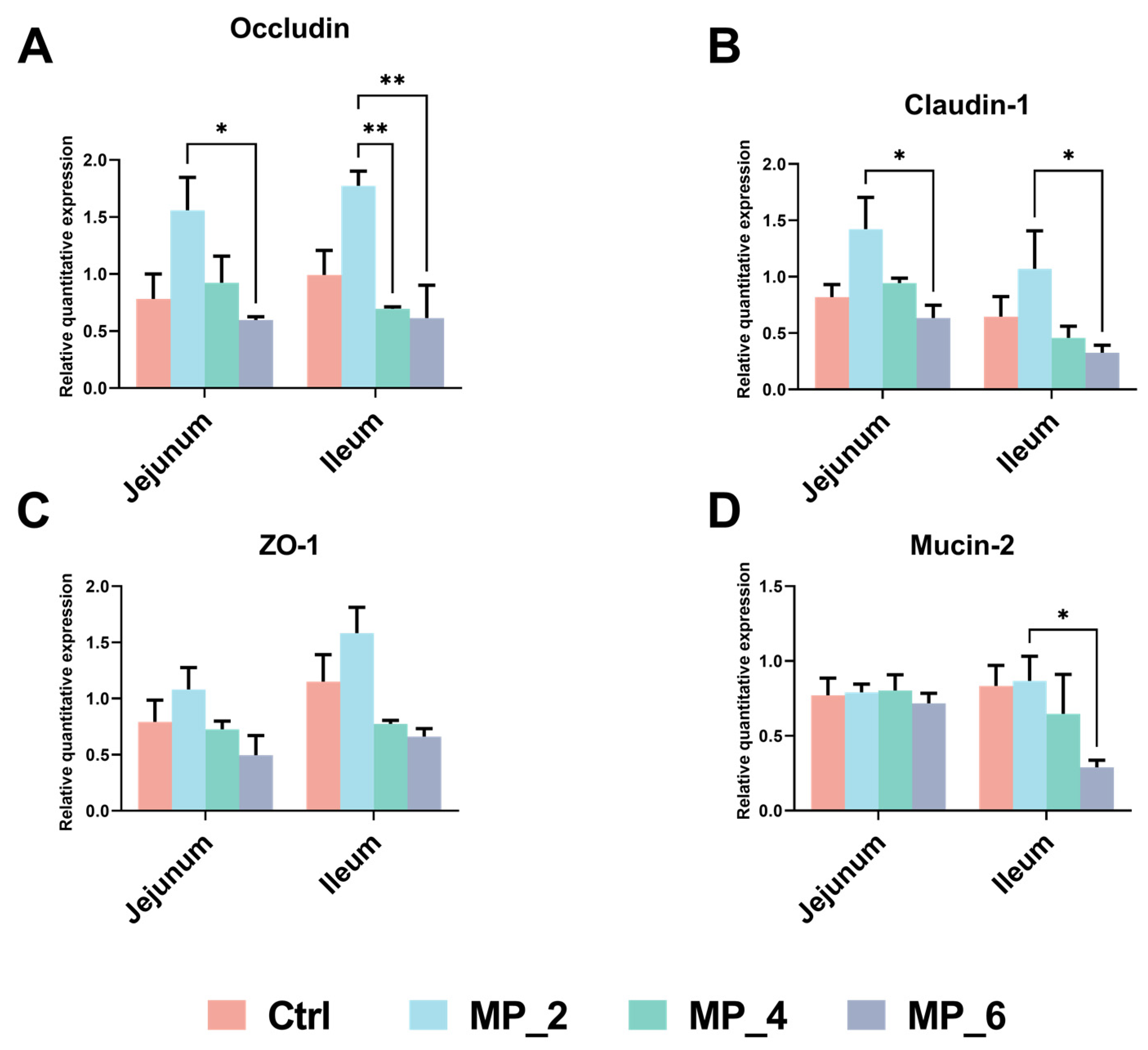
3.5. Morphology and Barrier Function of Intestine
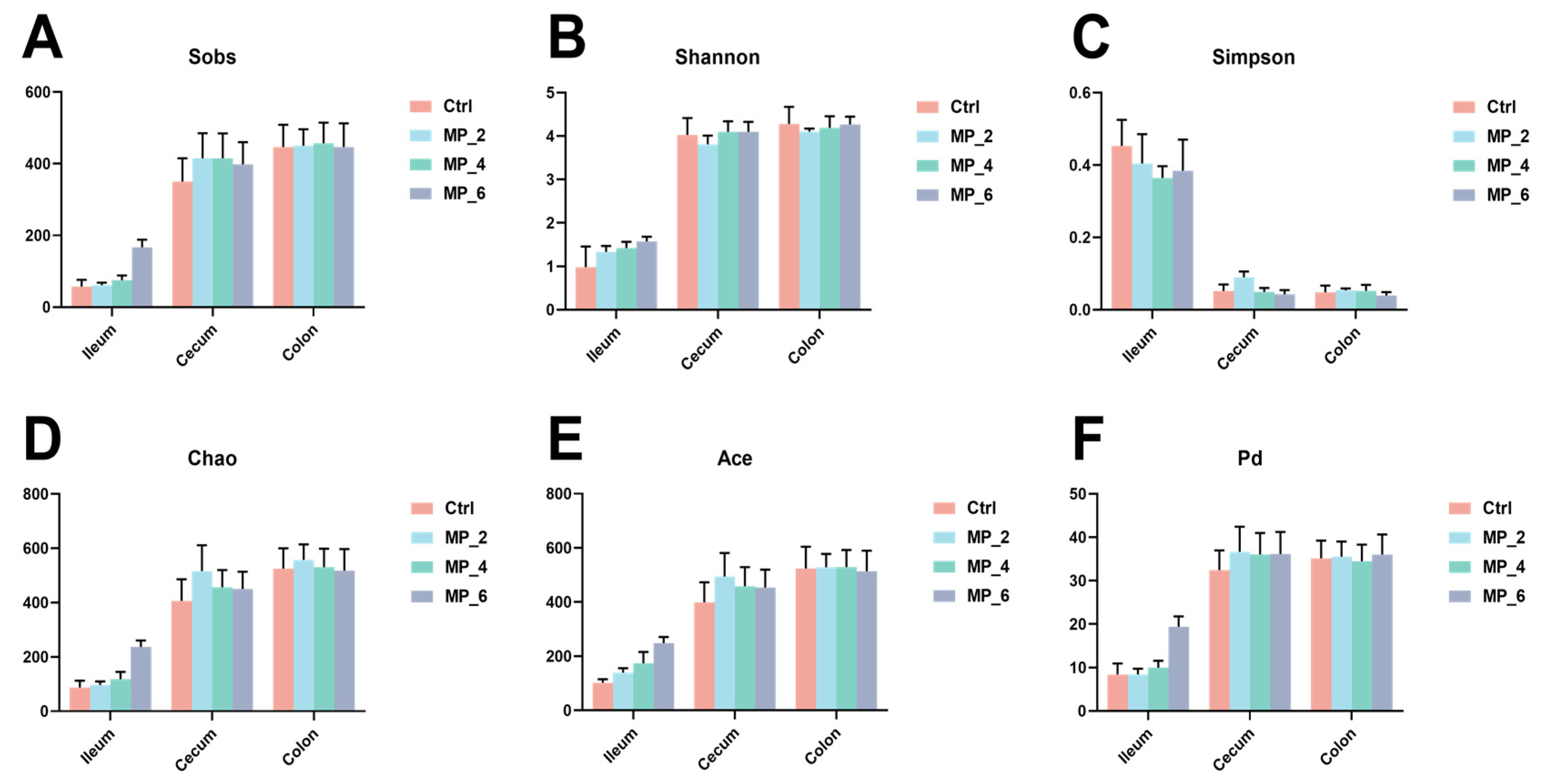
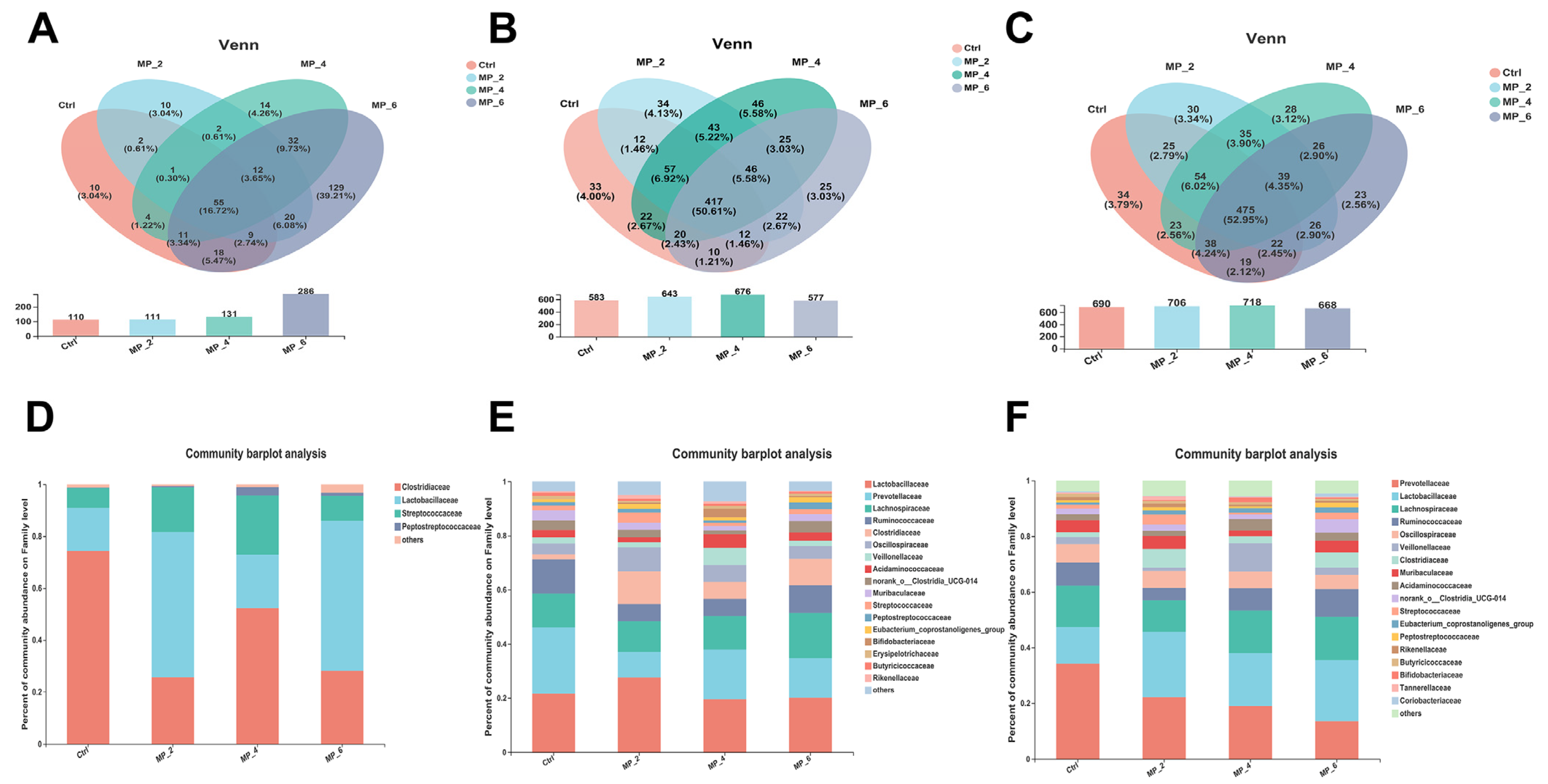
3.6. Bacterial Sequencing and α-Diversity in Ileum, Cecum and Colon of Piglets
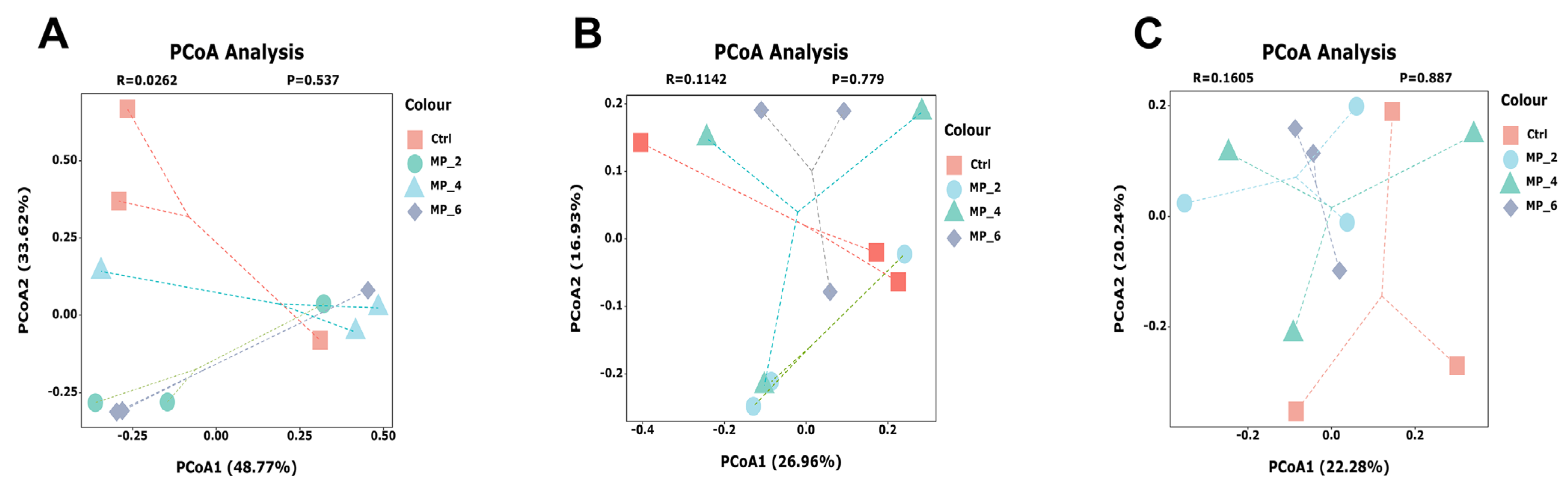
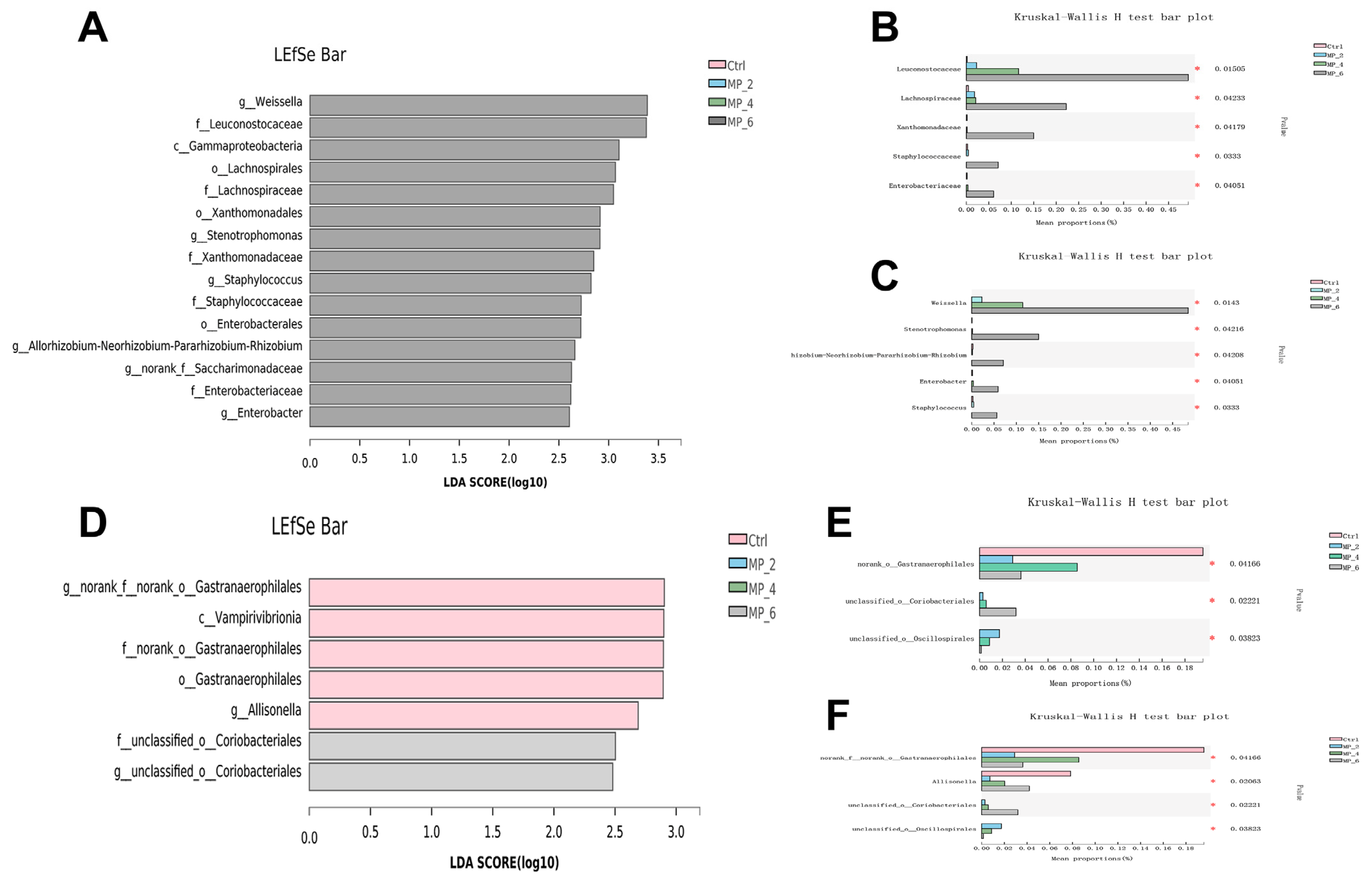
3.7. Bacterial Composition and β-Diversity Analysis in Ileum, Cecum and Colon of Piglets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Li, Y.; Guo, Y.; Wen, Z.S.; Jiang, X.M.; Ma, X.; Han, X.Y. Weaning stress perturbs gut microbiome and its metabolic profile in piglets. Sci. Rep. 2018, 8, 18068. [Google Scholar] [CrossRef] [PubMed]
- Moeser, A.J.; Pohl, C.S.; Rajput, M. Weaning stress and gastrointestinal barrier development: Implications for lifelong gut health in pigs. Anim. Nutr. 2017, 3, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.H.; Wu, X.Y.; Pan, Y.; Wang, L.; Cui, C.B.; Guo, Y.W.; Zhu, L.L.; Peng, J.; Wei, H.K. Early-life intervention using fecal microbiota combined with probiotics promotes gut microbiota maturation, regulates immune system development, and alleviates weaning stress in piglets. Int. J. Mol. Sci. 2020, 21, 503. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Kulus, M.; Jankowski, M.; Dompe, C.; Bryl, R.; Petitte, J.N.; Kempisty, B.; Mozdziak, P. COVID-19 pandemic is a call to search for alternative protein sources as food and feed: A review of possibilities. Nutrients 2021, 13, 150. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Liu, Y.Y.; Li, F.N.; Sun, A.; Lin, Q.; Huang, X.G.; Yin, Y.L. Effects of dietary ramie powder at various levels on growth performance, antioxidative capacity and fatty acid profile of finishing pigs. J. Anim. Physiol. Anim. Nutr. 2019, 103, 564–573. [Google Scholar] [CrossRef]
- Stein, H.H.; Shurson, G.C. Board-invited review: The use and application of distillers dried grains with solubles in swine diets. J. Anim. Sci. 2009, 87, 1292–1303. [Google Scholar] [CrossRef]
- Wang, X.Q.; Feng, Y.; Shu, G.; Jiang, Q.Y.; Yang, J.P.; Zhang, Z.F. Effect of dietary supplementation with hydrolyzed wheat gluten on growth performance, cell immunity and serum biochemical indices of weaned piglets (sus scrofa). Agric. Sci. China 2011, 10, 938–945. [Google Scholar] [CrossRef]
- Cheong, S.H.; Kim, K.H.; Jeon, B.T.; Park, P.J.; Hwang, I.H.; Choi, N.J.; Kim, E.T.; Hong, S.K.; Park, J.H.; Sung, S.H.; et al. Effect of mulberry silage supplementation during late fattening stage of Hanwoo (Bos taurus coreanae) steer on antioxidative enzyme activity within the longissimus muscle. Anim. Prod. Sci. 2012, 52, 240–247. [Google Scholar] [CrossRef]
- Cai, M.; Mu, L.; Wang, Z.L.; Liu, J.Y.; Liu, T.L.; Wanapat, M.; Huang, B.Z. Assessment of mulberry leaf as a potential feed supplement for animal feeding in PR China. Asian Australas. J. Anim. Sci. 2019, 32, 1145–1152. [Google Scholar] [CrossRef]
- Ma, G.Q.; Chai, X.Y.; Hou, G.G.; Zhao, F.L.; Meng, Q.G. Phytochemistry, bioactivities and future prospects of mulberry leaves: A review. Food Chem. 2022, 372, 131335. [Google Scholar] [CrossRef]
- Al-Kirshi, R.A.; Alimon, A.R.; Zulkifli, R.; Sazili, I.; Zahari, A.Q.W. The chemical composition and nutritive value of mulberry leaf meal as a protein source in poultry diets. Int. Semin. Anim. Ind. 2009, 98–102. Available online: http://repository.ipb.ac.id/handle/123456789/33831. (accessed on 12 November 2022).
- Chen, X.Y.; Zhang, T.; Wang, X.; Hamann, M.T.; Kang, J.; Yu, D.Q.; Chen, R.Y. A chemical investigation of the leaves of Morus alba L. Molecules 2018, 23, 1018. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kang, H.J.; Kim, S.Z.; Kwon, T.O.; Jeong, S.I.; Jang, S.I. Antioxidant effect of astragalin isolated from the leaves of Morus alba L. against free radical-induced oxidative hemolysis of human red blood cells. Arch. Pharmacal Res. 2013, 36, 912–917. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Maehara, R.; Kobuchi, S.; Tanaka, R.; Ohkita, M.; Matsumura, Y. Beneficial effects of gamma-aminobutyric acid on right ventricular pressure and pulmonary vascular remodeling in experimental pulmonary hypertension. Life Sci. 2012, 91, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, I.; Buhroo, Z.; Nagoo, S.A. Mulberry (Morus spp.): A versatile tree with inherent bioactive compounds of promising pharmaceutical and nutraceutical properties. J. Pharmacogn. Phytochem. 2019, 8, 731–738. [Google Scholar]
- Bhattacharjya, D.; Sadat, A.; Dam, P.; Buccini, D.F.; Mondal, R.; Biswas, T.; Biswas, K.; Sarkar, H.; Bhuimali, A.; Kati, A.; et al. Current concepts and prospects of mulberry fruits for nutraceutical and medicinal benefits. Curr. Opin. Food Sci. 2021, 40, 121–135. [Google Scholar] [CrossRef]
- Datta, R.K. Mulberry Cultivation and Utilization in India. FAO Electronic Conference on Mulberry for Animal Production (Morus-L). 2002. Available online: https://www.fao.org/3/X9895E/x9895e04.htm. (accessed on 15 October 2022).
- Ma, J.Y.; Ma, H.; Liu, S.J.; Wang, J.; Wang, H.L.; Zang, J.J.; Long, S.F.; Piao, X.S. Effect of mulberry leaf powder of varying levels on growth performance, immuno-antioxidant status, meat quality and intestinal health in finishing pigs. Antioxidants 2022, 11, 2243. [Google Scholar] [CrossRef] [PubMed]
- Doran, M.P.; Laca, E.A.; Sainz, R.D. Total tract and rumen digestibility of mulberry foliage (Morus alba), alfalfa hay and oat hay in sheep. Anim. Feed. Sci. Technol. 2007, 138, 239–253. [Google Scholar] [CrossRef]
- Lin, W.C.; Lee, M.T.; Chang, S.C.; Chang, Y.L.; Shih, C.H.; Yu, B.; Lee, T.T. Effects of mulberry leaves on production performance and the potential modulation of antioxidative status in laying hens. Poult. Sci. 2017, 96, 1191–1203. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Swine, 11th ed.; National Academy Press: Washington, DC, USA, 2012. [Google Scholar]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from bacterial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Goebel, B.M. A place for dna-dna reassociation and 16s ribosomal-rna sequence-analysis in the present species definition in bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef]
- Wang, Q.B.; Liu, C.Q.; Zhao, Y.F.; Kitsos, A.; Cannella, M.; Wang, S.K.; Han, L. Impacts of the COVID-19 pandemic on the dairy industry: Lessons from China and the United States and policy implications. J. Integr. Agric. 2020, 19, 2903–2915. [Google Scholar] [CrossRef]
- Woyengo, T.A.; Weihrauch, D.; Nyachoti, C.M. Effect of dietary phytic acid on performance and nutrient uptake in the small intestine of piglets. J. Anim. Sci. 2012, 90, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Emire, S.A.; Jha, Y.K.; Mekam, F. Role of anti-nutritional factors in food industry. Beverage Food World 2013, 2, 23–28. Available online: https://www.researchgate.net/publication/280722236_Role_of_Anti-nutritional_Factors_in_Food_Industry. (accessed on 24 October 2022).
- Stanogias, G.; Pearce, G.R. The digestion of fiber by pigs.1. The effects of amount and type of fiber on apparent digestibility, nitrogen-balance and rate of passage. Br. J. Nutr. 1985, 53, 513–530. [Google Scholar] [CrossRef]
- Ma, Y.D.; Lv, Q.F.; Zhao, D.D.; Wang, J.J.; Fu, Y.; Li, C.; Wu, G.F.; Liu, M.; Hu, J.M.; Lin, S.M.; et al. Intervention Effect of Taurine on lps-Induced Intestinal Mechanical Barrier Injury in Piglets. In Taurine 12: A Conditionally Essential Amino Acid; Schaffer, S.W., El Idrissi, A., Murakami, S., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2022; pp. 73–80. [Google Scholar] [CrossRef]
- Klein, R.; Nagy, O.; Tóthová, C.; Chovanová, F. Clinical and Diagnostic Significance of Lactate Dehydrogenase and Its Isoenzymes in Animals. Vet. Med. Int. 2020, 2020, 5346483. [Google Scholar] [CrossRef]
- Li, W.; Li, B.; Lv, J.Q.; Dong, L.; Zhang, L.L.; Wang, T. Choline supplementation improves the lipid metabolism of intrauterine-growth-restricted pigs. Asian Australas. J. Anim. Sci. 2018, 31, 686–695. [Google Scholar] [CrossRef]
- Bowden, D.M. Non-esterified fatty acids and ketone bodies in blood as indicators of nutritional status in ruminants: A review. Can. J. Anim. Sci. 1971, 51, 1–13. [Google Scholar] [CrossRef]
- Zhao, Y.W.; Niu, Y.; He, J.T.; Gan, Z.D.; Ji, S.L.; Zhang, L.L.; Wang, C.; Wang, T. Effects of dietary dihydroartemisinin supplementation on growth performance, hepatic inflammation, and lipid metabolism in weaned piglets with intrauterine growth retardation. Anim. Sci. J. 2020, 91, e13363. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Min, B.J.; Chen, Y.J.; Yoo, J.S.; Wang, Q.; Kimi, J.D.; Kim, I.H. Evaluation of FSP (fermented soy protein) to replace soybean meal in weaned pigs: Growth performance, blood urea nitrogen and total protein concentrations in serum and nutrient digestibility. Asian Australas. J. Anim. Sci. 2007, 20, 1874–1879. [Google Scholar] [CrossRef]
- Novais, A.K.; Deschene, K.; Martel-Kennes, Y.; Roy, C.; Laforest, J.P.; Lessard, M.; Matte, J.J.; Lapointe, J. Weaning differentially affects mitochondrial function, oxidative stress, inflammation and apoptosis in normal and low birth weight piglets. PLoS ONE 2021, 16, e0247188. [Google Scholar] [CrossRef]
- Lei, X.G.; Zhu, J.H.; Cheng, W.H.; Bao, Y.P.; Ho, Y.S.; Reddi, A.R.; Holmgren, A.; Arner, E.S.J. Paradoxical roles of antioxidant enzymes: Basic mechanisms and health implications. Physiol. Rev. 2016, 96, 307–364. [Google Scholar] [CrossRef]
- Zhao, X.J.; Li, L.; Luo, Q.L.; Ye, M.Q.; Luo, G.Q.; Kuang, Z.S. Effects of mulberry (Morus alba L.) leaf polysaccharides on growth performance, diarrhea, blood parameters, and gut microbiota of early-weanling pigs. Livest. Sci. 2015, 177, 88–94. [Google Scholar] [CrossRef]
- Kim, D.S.; Kim, S.H.; Lee, G.J.; Kim, H.K. Antioxidant activities and polyphenol content of Morus alba leaf extracts collected from varying regions. Planta Med. 2014, 80, 1476–1477. [Google Scholar] [CrossRef]
- Radojkovic, M.M.; Zekovic, Z.P.; Vidovic, S.S.; Kocar, D.D.; Maskovic, P.Z. Free radical scavenging activity and total phenolic and flavonoid contents of mulberry (Morus spp. L., Moraceae) extracts. Hem. Ind. 2012, 66, 545–550. [Google Scholar] [CrossRef]
- Naowaboot, J.; Pannangpetch, P.; Kukongviriyapan, V.; Kongyingyoes, B.; Kukongviriyapan, U. Antihyperglycemic, antioxidant and antiglycation activities of mulberry leaf extract in streptozotocin-induced chronic diabetic rats. Plant Foods Hum. Nutr. 2009, 64, 116–121. [Google Scholar] [CrossRef]
- Moeser, A.J.; Borst, L.B.; Overman, B.L.; Pittman, J.S. Defects in small intestinal epithelial barrier function and morphology associated with peri-weaning failure to thrive syndrome (PFTS) in swine. Res. Vet. Sci. 2012, 93, 975–982. [Google Scholar] [CrossRef]
- Moeser, A.J.; Vander Klok, C.; Ryan, K.A.; Wooten, J.G.; Little, D.; Cook, V.L.; Blikslager, A.T. Stress signaling pathways activated by weaning mediate intestinal dysfunction in the pig. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G173–G181. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.Y.; Long, S.F.; Wang, J.; Gao, J.; Piao, X.S. Microencapsulated essential oils combined with organic acids improves immune antioxidant capacity and intestinal barrier function as well as modulates the hindgut bacterial community in piglets. J. Anim. Sci. Biotechnol. 2022, 13, 16. [Google Scholar] [CrossRef]
- Ma, J.Y.; Piao, X.S.; Mahfuz, S.; Long, S.F.; Wang, J. The interaction among gut microbes, the intestinal barrier and short chain fatty acids. Anim. Nutr. 2022, 9, 159–174. [Google Scholar] [CrossRef]
- Horng, C.T.; Liu, Z.H.; Huang, Y.T.; Lee, H.J.; Wang, C.J. Extract from mulberry (Morus australis) leaf decelerate acetaminophen induced hepatic inflammation involving downregulation of myeloid differentiation factor 88 (MyD88) signals. J. Food Drug Anal. 2017, 25, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Xu, R.Y.; Zhang, H.; Su, Y.; Zhu, W.Y. Swine gut microbiota and its interaction with host nutrient metabolism. Anim. Nutr. 2020, 6, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.V.; Frassetto, A.; Kowalik, E.J.; Nawrocki, A.R.; Lu, M.F.M.; Kosinski, J.R.; Hubert, J.A.; Szeto, D.; Yao, X.R.; Forrest, G.; et al. Butyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanisms. PLoS ONE 2012, 7, e35240. [Google Scholar] [CrossRef] [PubMed]
- Woodhouse, C.A.; Patel, V.C.; Singanayagam, A.; Shawcross, D.L. Review article: The gut microbiome as a therapeutic target in the pathogenesis and treatment of chronic liver disease. Aliment. Pharmacol. Ther. 2018, 47, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Abbott, I.J.; Slavin, M.A.; Turnidge, J.D.; Thursky, K.A.; Worth, L.J. Stenotrophomonas maltophilia: Emerging disease patterns and challenges for treatment. Expert Rev. Anti Infect. Ther. 2011, 9, 471–488. [Google Scholar] [CrossRef]
- Aranaz, P.; Ramos-Lopez, O.; Cuevas-Sierra, A.; Martinez, J.A.; Milagro, F.I.; Riezu-Boj, J.I. A predictive regression model of the obesity-related inflammatory status based on gut microbiota composition. Int. J. Obes. 2021, 45, 2261–2268. [Google Scholar] [CrossRef]
- Wong, V.W.S.; Tse, C.H.; Lam, T.T.Y.; Wong, G.L.H.; Chim, A.M.L.; Chu, W.C.W.; Yeung, D.K.W.; Law, P.T.W.; Kwan, H.S.; Yu, J.; et al. Molecular characterization of the fecal microbiota in patients with nonalcoholic steatohepatitis—A longitudinal study. PLoS ONE 2013, 8, e62885. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Mulberry Leaf Powder | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 2% | 4% | 6% | ANOVA | Linear | Quadratic | ||
| D 0 to 14 | ||||||||
| ADG, kg/d | 394 a | 389 a | 358 ab | 328 b | 13.00 | 0.04 | 0.01 | 0.37 |
| ADFI, kg/d | 558 a | 553 a | 495 ab | 451 b | 17.24 | 0.01 | 0.01 | 0.30 |
| FCR | 1.41 | 1.42 | 1.38 | 1.38 | 0.02 | 0.17 | 0.06 | 0.59 |
| Diarrhea rate, % | 3.27 | 2.68 | 1.19 | 5.06 | 1.56 | 0.44 | 0.60 | 0.20 |
| D 15 to 28 | ||||||||
| ADG, kg/d | 585 | 590 | 555 | 533 | 19.48 | 0.23 | 0.07 | 0.51 |
| ADFI, kg/d | 928 | 912 | 858 | 813 | 32.89 | 0.15 | 0.03 | 0.68 |
| FCR | 1.59 | 1.54 | 1.55 | 1.51 | 0.02 | 0.10 | 0.03 | 0.72 |
| Diarrhea rate, % | 1.79 | 0.59 | 1.57 | 1.79 | 0.47 | 0.31 | 0.65 | 0.18 |
| D 0 to 28 | ||||||||
| ADG, kg/d | 490 a | 490 a | 456 ab | 430 b | 8.44 | 0.01 | 0.01 | 0.17 |
| ADFI, kg/d | 743 a | 732 a | 677 b | 632 b | 13.15 | 0.01 | 0.01 | 0.25 |
| FCR | 1.51 | 1.50 | 1.48 | 1.46 | 0.01 | 0.14 | 0.03 | 0.91 |
| Diarrhea rate, % | 2.53 | 1.64 | 1.40 | 3.42 | 0.69 | 0.25 | 0.46 | 0.08 |
| Item | Mulberry Leaf Powder | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 2% | 4% | 6% | ANOVA | Linear | Quadratic | ||
| D 14 | ||||||||
| IgA, μg/mL | 12.74 | 13.55 | 14.79 | 13.85 | 0.65 | 0.27 | 0.17 | 0.23 |
| IgG, mg/mL | 6.76 | 6.33 | 6.79 | 6.97 | 0.25 | 0.39 | 0.36 | 0.27 |
| IgM, μg/mL | 5.84 | 6.00 | 5.85 | 5.95 | 0.27 | 0.96 | 0.86 | 0.91 |
| IL-1β, ng/L | 103 | 104 | 104 | 103 | 3.45 | 0.99 | 0.93 | 0.85 |
| IL-6, ng/L | 33.62 | 31.54 | 32.49 | 33.46 | 1.27 | 0.65 | 0.93 | 0.28 |
| IL-8, ng/L | 64.32 | 68.66 | 64.46 | 65.95 | 2.28 | 0.54 | 0.95 | 0.55 |
| IL-10, ng/L | 18.86 | 19.27 | 19.37 | 18.93 | 0.90 | 0.97 | 0.94 | 0.65 |
| TNF-α, ng/L | 49.97 | 51.52 | 52.13 | 52.04 | 1.49 | 0.73 | 0.34 | 0.60 |
| IFN-γ, pg/mL | 143 | 145 | 149 | 149 | 4.22 | 0.67 | 0.29 | 0.78 |
| D 28 | ||||||||
| IgA, μg/mL | 14.02 | 14.93 | 15.80 | 14.71 | 0.58 | 0.28 | 0.30 | 0.13 |
| IgG, mg/mL | 6.93 | 7.27 | 7.42 | 8.12 | 0.29 | 0.12 | 0.06 | 0.55 |
| IgM, μg/mL | 6.38 | 5.79 | 5.48 | 6.20 | 0.24 | 0.14 | 0.47 | 0.04 |
| IL-1β, ng/L | 102 | 109 | 105 | 109 | 4.83 | 0.67 | 0.46 | 0.66 |
| IL-6, ng/L | 32.05 | 35.88 | 36.35 | 35.24 | 0.93 | 0.06 | 0.05 | 0.04 |
| IL-8, ng/L | 69.70 | 69.10 | 65.25 | 75.23 | 3.49 | 0.34 | 0.45 | 0.18 |
| IL-10, ng/L | 19.69 | 20.37 | 20.25 | 20.13 | 0.62 | 0.87 | 0.68 | 0.54 |
| TNF-α, ng/L | 54.93 | 54.31 | 55.11 | 55.05 | 2.63 | 1.00 | 0.92 | 0.92 |
| IFN-γ, pg/mL | 165 | 156 | 153 | 170 | 5.57 | 0.22 | 0.65 | 0.06 |
| Item | Mulberry Leaf Powder | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 2% | 4% | 6% | ANOVA | Linear | Quadratic | ||
| D 14 | ||||||||
| GLU, mmol/L | 5.95 | 5.47 | 5.87 | 5.11 | 0.47 | 0.59 | 0.35 | 0.78 |
| TC, mmol/L | 1.69 | 1.61 | 2.25 | 1.99 | 0.32 | 0.53 | 0.33 | 0.79 |
| TG, mmol/L | 0.50 | 0.48 | 0.77 | 0.50 | 0.11 | 0.29 | 0.59 | 0.29 |
| HDL, mmol/L | 0.45 | 0.55 | 0.68 | 0.64 | 0.07 | 0.22 | 0.07 | 0.36 |
| LDL, mmol/L | 1.40 | 1.26 | 1.69 | 1.56 | 0.17 | 0.40 | 0.29 | 0.98 |
| ALT, U/L | 65.86 | 48.29 | 43.05 | 62.09 | 11.37 | 0.49 | 0.76 | 0.16 |
| AST, U/L | 53.10 | 54.45 | 51.55 | 45.90 | 6.07 | 0.77 | 0.40 | 0.58 |
| TP, g/L | 47.09 | 48.05 | 50.52 | 49.88 | 2.45 | 0.74 | 0.36 | 0.75 |
| ALB, g/L | 20.68 | 23.01 | 23.78 | 22.65 | 1.88 | 0.70 | 0.46 | 0.39 |
| GLB, g/L | 28.26 | 24.80 | 27.11 | 27.39 | 2.36 | 0.76 | 0.98 | 0.46 |
| ALP, U/L | 238 | 242 | 218 | 217 | 45.85 | 0.97 | 0.69 | 0.97 |
| LDH, U/L | 586 | 602 | 496 | 488 | 38.33 | 0.17 | 0.06 | 0.76 |
| UA, μmol/L | 35.39 | 32.67 | 34.11 | 35.04 | 1.34 | 0.52 | 0.95 | 0.22 |
| BUN, mmol/L | 1.93 | 1.48 | 2.45 | 1.51 | 0.57 | 0.61 | 0.91 | 0.68 |
| DLA, μmol/L | 6.68 a | 6.11 b | 6.54 a | 6.64 a | 0.11 | 0.04 | 0.58 | 0.02 |
| NEFA, μmol/L | 157 | 152 | 154 | 154 | 6.80 | 0.97 | 0.88 | 0.70 |
| D 28 | ||||||||
| GLU, mmol/L | 6.40 | 6.17 | 5.67 | 6.44 | 0.45 | 0.62 | 0.86 | 0.31 |
| TC, mmol/L | 1.54 | 1.71 | 2.32 | 2.04 | 0.27 | 0.28 | 0.13 | 0.44 |
| TG, mmol/L | 0.38 | 0.39 | 0.62 | 0.58 | 0.06 | 0.08 | 0.03 | 0.71 |
| HDL, mmol/L | 0.49 | 0.61 | 0.76 | 0.60 | 0.10 | 0.38 | 0.33 | 0.21 |
| LDL, mmol/L | 1.25 | 1.38 | 1.69 | 1.52 | 0.19 | 0.45 | 0.23 | 0.46 |
| ALT, U/L | 56.05 | 48.12 | 48.85 | 45.51 | 5.37 | 0.58 | 0.25 | 0.68 |
| AST, U/L | 75.47 | 48.98 | 69.09 | 42.29 | 9.10 | 0.12 | 0.10 | 0.99 |
| TP, g/L | 51.28 | 48.32 | 52.59 | 52.12 | 2.02 | 0.49 | 0.48 | 0.56 |
| ALB, g/L | 23.98 | 21.70 | 23.73 | 23.37 | 1.80 | 0.81 | 0.98 | 0.61 |
| GLB, g/L | 26.34 | 26.16 | 27.18 | 26.58 | 1.48 | 0.96 | 0.80 | 0.89 |
| ALP, U/L | 316 | 285 | 286 | 226 | 30.93 | 0.31 | 0.10 | 0.66 |
| LDH, U/L | 636 a | 390 b | 547 ab | 429 ab | 50.04 | 0.04 | 0.08 | 0.25 |
| UA, μmol/L | 26.79 | 35.19 | 28.84 | 27.44 | 3.65 | 0.42 | 0.80 | 0.23 |
| BUN, mmol/L | 2.84 | 1.69 | 1.61 | 1.51 | 0.31 | 0.07 | 0.03 | 0.15 |
| DLA, μmol/L | 6.59 | 7.12 | 6.88 | 7.00 | 0.18 | 0.28 | 0.26 | 0.29 |
| NEFA, μmol/L | 161 b | 165 ab | 170 ab | 186 a | 4.83 | 0.04 | 0.02 | 0.27 |
| Item | Mulberry Leaf Powder | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 2% | 4% | 6% | ANOVA | Linear | Quadratic | ||
| Duodenum | ||||||||
| Villus height | 370 | 363 | 305 | 308 | 44.60 | 0.64 | 0.89 | 0.23 |
| Crypt depth | 312 | 339 | 420 | 369 | 47.23 | 0.47 | 0.28 | 0.44 |
| VH/CD | 1.22 | 1.07 | 0.77 | 0.84 | 0.18 | 0.43 | 0.48 | 0.16 |
| Jejunum | ||||||||
| Villus height | 376 | 460 | 420 | 543 | 62.61 | 0.36 | 0.15 | 0.76 |
| Crypt depth | 324 | 253 | 305 | 282 | 24.11 | 0.28 | 0.52 | 0.35 |
| VH/CD | 1.19 | 1.85 | 1.38 | 2.03 | 0.26 | 0.18 | 0.13 | 0.98 |
| Ileum | ||||||||
| Villus height | 213 | 372 | 293 | 313 | 33.13 | 0.07 | 0.06 | 0.76 |
| Crypt depth | 191 | 213 | 267 | 221 | 23.67 | 0.25 | 0.23 | 0.20 |
| VH/CD | 1.12 | 1.75 | 1.12 | 1.42 | 0.29 | 0.40 | 0.29 | 0.70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Wang, J.; Jin, X.; Liu, S.; Tang, S.; Zhang, Z.; Long, S.; Piao, X. Effect of Dietary Supplemented with Mulberry Leaf Powder on Growth Performance, Serum Metabolites, Antioxidant Property and Intestinal Health of Weaned Piglets. Antioxidants 2023, 12, 307. https://doi.org/10.3390/antiox12020307
Ma J, Wang J, Jin X, Liu S, Tang S, Zhang Z, Long S, Piao X. Effect of Dietary Supplemented with Mulberry Leaf Powder on Growth Performance, Serum Metabolites, Antioxidant Property and Intestinal Health of Weaned Piglets. Antioxidants. 2023; 12(2):307. https://doi.org/10.3390/antiox12020307
Chicago/Turabian StyleMa, Jiayu, Jian Wang, Xiaoyun Jin, Sujie Liu, Shaofeng Tang, Zhenghua Zhang, Shenfei Long, and Xiangshu Piao. 2023. "Effect of Dietary Supplemented with Mulberry Leaf Powder on Growth Performance, Serum Metabolites, Antioxidant Property and Intestinal Health of Weaned Piglets" Antioxidants 12, no. 2: 307. https://doi.org/10.3390/antiox12020307
APA StyleMa, J., Wang, J., Jin, X., Liu, S., Tang, S., Zhang, Z., Long, S., & Piao, X. (2023). Effect of Dietary Supplemented with Mulberry Leaf Powder on Growth Performance, Serum Metabolites, Antioxidant Property and Intestinal Health of Weaned Piglets. Antioxidants, 12(2), 307. https://doi.org/10.3390/antiox12020307