
Plasma CoQ10 Status in Patients with Propionic Acidaemia and Possible Benefit of Treatment with Ubiquinol

 , , , ,
, , , ,
Abstract
:1. Introduction
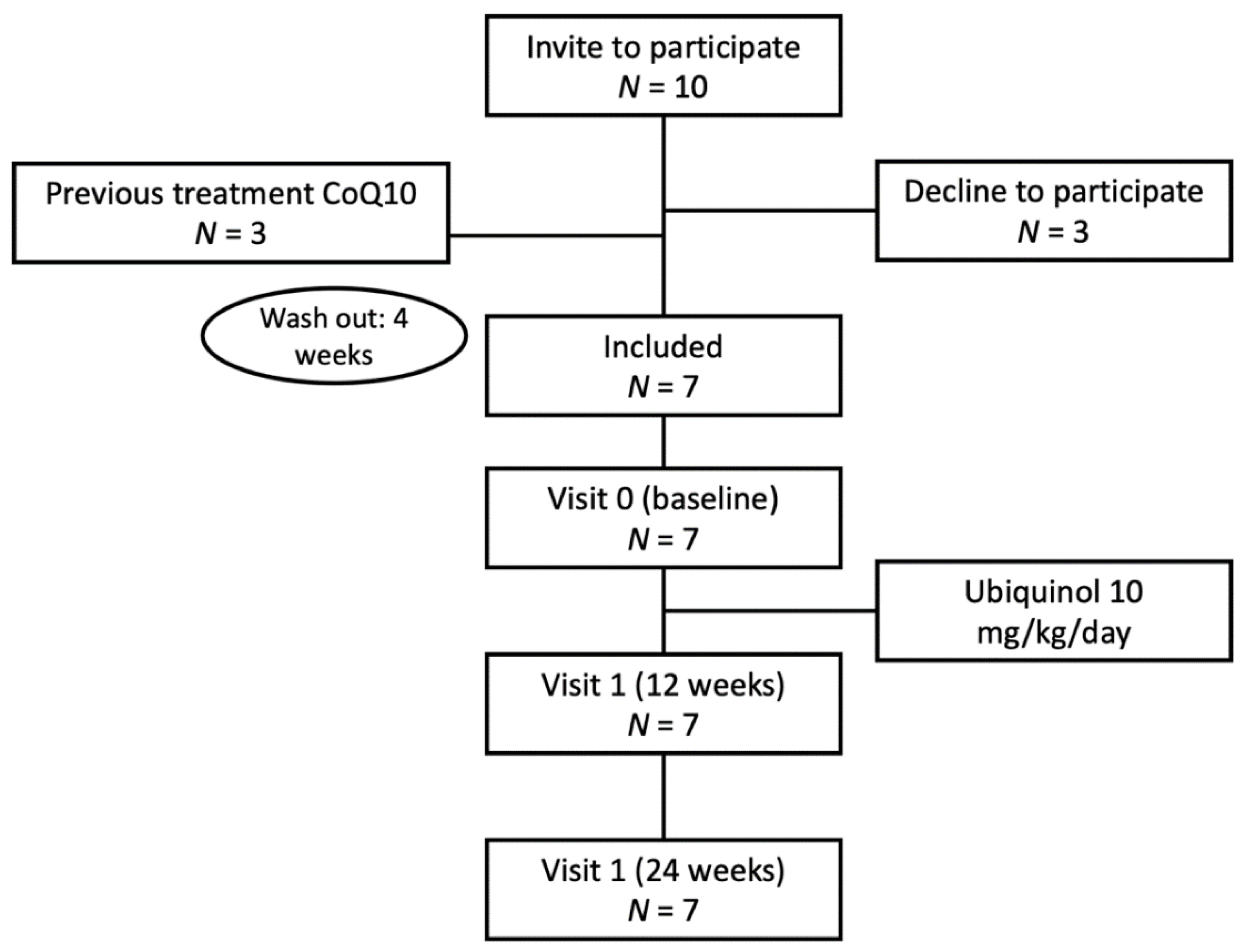
2. Materials and Methods
2.1. Patients and Analyses
2.2. Statistical Analysis
2.3. Ethics Statement
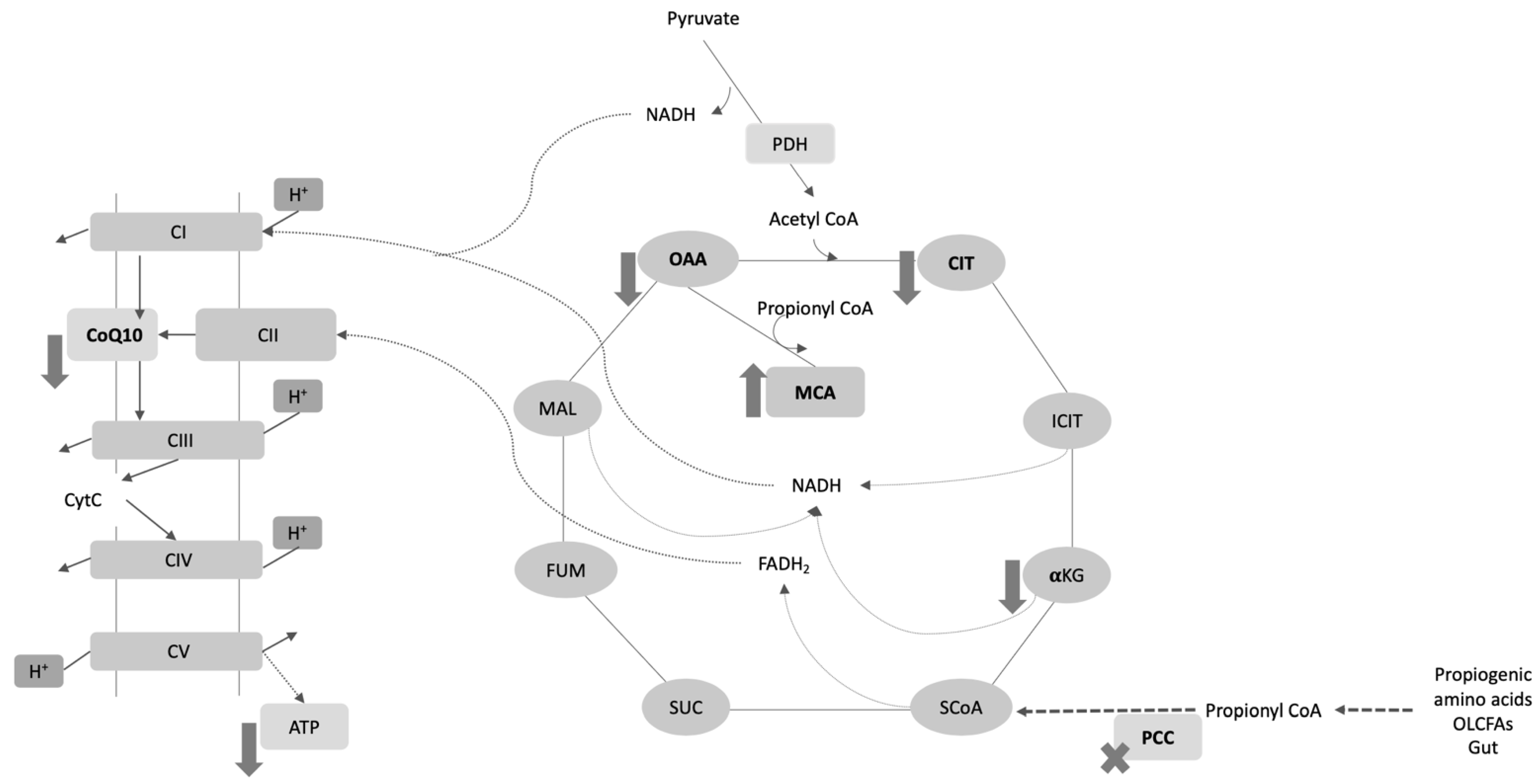
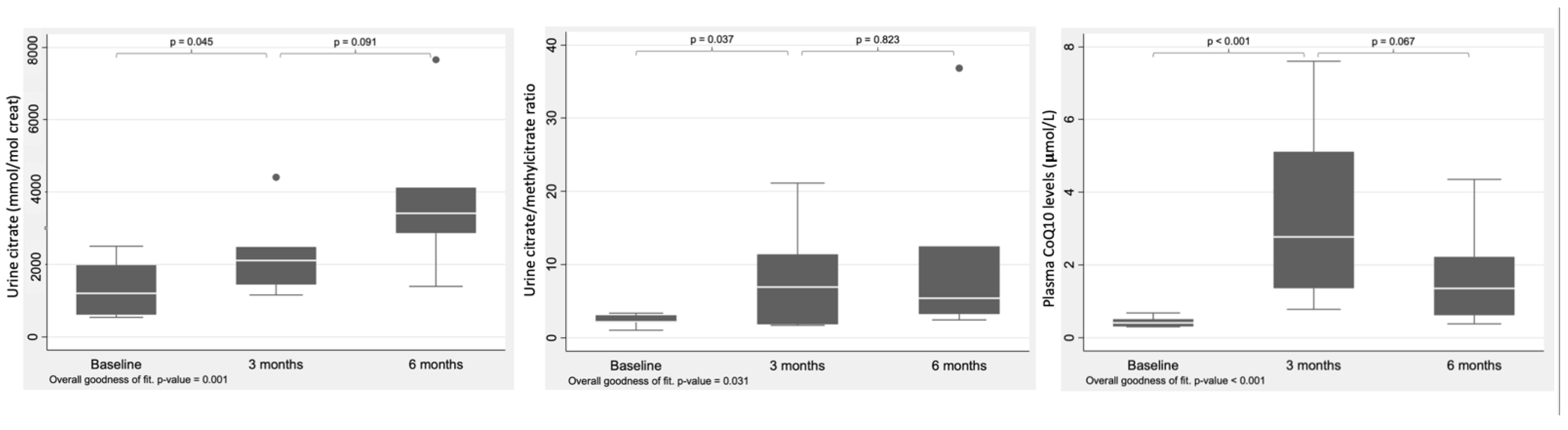
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wongkittichote, P.; Ah Mew, N.; Chapman, K.A. Propionyl-CoA carboxylase—A review. Mol. Genet. Metab. 2017, 122, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Haijes, H.A.; van Hasselt, P.M.; Jans, J.J.M.; Verhoeven-Duif, N.M. Pathophysiology of propionic and methylmalonic acidemias. Part 2: Treatment strategies. J. Inherit. Metab. Dis. 2019, 42, 745–761. [Google Scholar] [CrossRef] [PubMed]
- Stanescu, S.; Belanger-Quintana, A.; Fernandez-Felix, B.M.; Arrieta, F.; Quintero, V.; Maldonado, M.S.; Alcaide, P.; Martínez-Pardo, M. Severe anemia in patients with Propionic acidemia is associated with branched-chain amino acid imbalance. Orphanet J. Rare Dis. 2021, 16, 226. [Google Scholar] [CrossRef] [PubMed]
- Molema, F.; Gleich, F.; Burgard, P.; van der Ploeg, A.T.; Summar, M.L.; Chapman, K.A.; Lund, A.M.; Rizopoulos, D.; Kölker, S.; Williams, M. Decreased plasma l-arginine levels in organic acidurias (MMA and PA) and decreased plasma branched-chain amino acid levels in urea cycle disorders as a potential cause of growth retardation: Options for treatment. Mol. Genet. Metab. 2019, 126, 397–405. [Google Scholar] [CrossRef]
- Chemelli, A.P.; Schocke, M.; Sperl, W.; Trieb, T.; Aichner, F.; Felber, S. Magnetic resonance spectroscopy (MRS) in five patients with treated propionic acidemia. J. Magn. Reson. Imaging 2000, 11, 596–600. [Google Scholar] [CrossRef]
- Shoffner, J. Oxidative phosphorylation diseases. In The Metabolic and Molecular Bases of Inherited Disease; Scriver, C.R., Beaudet, A.L., Sly, W.S., Valle, D., Eds.; McGraw-Hill: New York, NY, USA, 2001; pp. 2367–2423. [Google Scholar]
- Martinez Alvarez, L.; Jameson, E.; Parry, N.R.; Lloyd, C.; Ashworth, J.L. Optic neuropathy in methylmalonic acidemia and propionic acidemia. Br. J. Ophthalmol. 2016, 100, 98–104. [Google Scholar] [CrossRef]
- Williams, Z.R.; Hurley, P.E.; Altiparmak, U.E.; Feldon, S.E.; Arnold, G.L.; Eggenberger, E.; Mejico, L.J. Late onset optic neuropathy in methylmalonic and propionic acidemia. Am. J. Ophthalmol. 2009, 147, 929–933. [Google Scholar] [CrossRef]
- Dimitrov, B.; Molema, F.; Williams, M.; Schmiesing, J.; Mühlhausen, C.; Baumgartner, M.R.; Schumann, A.; Kölker, S. Organic acidurias: Major gaps, new challenges, and a yet unfulfilled promise. J. Inherit. Metab. Dis. 2021, 44, 9–21. [Google Scholar] [CrossRef]
- de Keyzer, Y.; Valayannopoulos, V.; Benoist, J.F.; Batteux, F.; Lacaille, F.; Hubert, L.; Chrétien, D.; Chadefeaux-Vekemans, B.; Niaudet, P.; Touati, G.; et al. Multiple OXPHOS deficiency in the liver, kidney, heart, and skeletal muscle of patients with methylmalonic aciduria and propionic aciduria. Pediatr. Res. 2009, 66, 91–95. [Google Scholar] [CrossRef]
- Schwab, M.A.; Sauer, S.W.; Okun, J.G.; Nijtmans, L.G.; Rodenburg, R.J.; van den Heuvel, L.P.; Dröse, S.; Brandt, U.; Hoffmann, G.F.; Ter Laak, H.; et al. Secondary mitochondrial dysfunction in propionic aciduria: A pathogenic role for endogenous mitochondrial toxins. Biochem. J. 2006, 398, 107–112. [Google Scholar] [CrossRef]
- Gallego-Villar, L.; Rivera-Barahona, A.; Cuevas-Martín, C.; Guenzel, A.; Pérez, B.; Barry, M.A.; Murphy, M.P.; Logan, A.; Gonzalez-Quintana, A.; Martín, M.A.; et al. In Vivo evidence of mitochondrial dysfunction and altered redox homeostasis in a genetic mouse model of propionic acidemia: Implications for the pathophysiology of this disorder. Free Radic. Biol. Med. 2016, 96, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mardach, R.; Verity, M.A.; Cederbaum, S.D. Clinical, pathological, and biochemical studies in a patient with propionic acidemia and fatal cardiomyopathy. Mol. Genet. Metab. 2005, 85, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Fragaki, K.; Cano, A.; Benoist, J.F.; Rigal, O.; Chaussenot, A.; Rouzier, C.; Bannwarth, S.; Caruba, C.; Chabrol, B.; Paquis-Flucklinger, V. Fatal heart failure associated with CoQ10 and multiple OXPHOS deficiency in a child with propionic acidemia. Mitochondrion 2011, 11, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Baruteau, J.; Hargreaves, I.; Krywawych, S.; Chalasani, A.; Land, J.M.; Davison, J.E.; Kwok, M.K.; Christov, G.; Karimova, A.; Ashworth, M.; et al. Successful reversal of propionic acidaemia associated cardiomyopathy: Evidence for low myocardial coenzyme Q10 status and secondary mitochondrial dysfunction as an underlying pathophysiological mechanism. Mitochondrion 2014, 17, 150–156. [Google Scholar] [CrossRef]
- Stanescu, S.; Belanger-Quintana, A.; Fernandez-Felix, B.M.; Ruiz-Sala, P.; Del Valle, M.; Garcia, F.; Arrieta, F.; Martinez-Pardo, M. Interorgan amino acid interchange in propionic acidemia: The missing key to understanding its physiopathology. Amino Acids 2022. Epub ahead of print. [Google Scholar] [CrossRef]
- Longo, N.; Price, L.B.; Gappmaier, E.; Cantor, N.L.; Ernst, S.L.; Bailey, C.; Pasquali, M. Anaplerotic therapy in propionic acidemia. Mol. Genet. Metab. 2017, 122, 51–59. [Google Scholar] [CrossRef]
- Brunengraber, H.; Roe, C.R. Anaplerotic molecules: Current and future. J. Inherit. Metab. Dis. 2006, 29, 327–331. [Google Scholar] [CrossRef]
- Gallego-Villar, L.; Pérez-Cerdá, C.; Pérez, B.; Abia, D.; Ugarte, M.; Richard, E.; Desviat, L.R. Functional characterization of novel genotypes and cellular oxidative stress studies in propionic acidemia. J. Inherit. Metab. Dis. 2013, 36, 731–740. [Google Scholar] [CrossRef]
- Gallego-Villar, L.; Pérez, B.; Ugarte, M.; Desviat, L.R.; Richard, E. Antioxidants successfully reduce ROS production in propionic acidemia fibroblasts. Biochem. Biophys. Res. Commun. 2014, 452, 457–461. [Google Scholar] [CrossRef]
- Bentinger, M.; Tekle, M.; Dallner, G. Coenzyme Q--biosynthesis and functions. Biochem. Biophys. Res. Commun. 2010, 396, 74–79. [Google Scholar] [CrossRef]
- Gómez-Díaz, C.; Rodríguez-Aguilera, J.C.; Barroso, M.P.; Villalba, J.M.; Navarro, F.; Crane, F.L.; Navas, P. Antioxidant ascorbate is stabilized by NADH-coenzyme Q10 reductase in the plasma membrane. J. Bioenerg. Biomembr. 1997, 29, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Papucci, L.; Schiavone, N.; Witort, E.; Donnini, M.; Lapucci, A.; Tempestini, A.; Formigli, L.; Zecchi-Orlandini, S.; Orlandini, G.; Carella, G.; et al. Coenzyme q10 prevents apoptosis by inhibiting mitochondrial depolarization independently of its free radical scavenging property. J. Biol. Chem. 2003, 278, 28220–28228. [Google Scholar] [CrossRef] [PubMed]
- Schmelzer, C.; Lindner, I.; Vock, C.; Fujii, K.; Döring, F. Functional connections and pathways of coenzyme Q10-inducible genes: An in-silico study. IUBMB Life 2007, 59, 628–633. [Google Scholar] [PubMed]
- Thomas, S.R.; Neuzil, J.; Stocker, R. Cosupplementation with coenzyme Q prevents the prooxidant effect of alpha-tocopherol and increases the resistance of LDL to transition metal-dependent oxidation initiation. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 687–696. [Google Scholar] [CrossRef]
- Hamilton, S.J.; Chew, G.T.; Watts, G.F. Therapeutic regulation of endothelial dysfunction in type 2 diabetes mellitus. Diab. Vasc. Dis. Res. 2007, 4, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Gutiérrez, A.; González-García, P.; Díaz-Casado, M.E.; Barriocanal-Casado, E.; López-Herrador, S.; Quinzii, C.M.; López, L.C. Metabolic Targets of Coenzyme Q10 in Mitochondria. Antioxidants 2021, 10, 520. [Google Scholar] [CrossRef]
- Longo, N.; Sass, J.O.; Jurecka, A.; Vockley, J. Biomarkers for drug development in propionic and methylmalonic acidemias. J. Inherit. Metab. Dis. Epub ahead of print. [CrossRef]
- Al-Dirbashi, O.Y.; Alfadhel, M.; Al-Thihli, K.; Al Dhahouri, N.; Langhans, C.D.; Al Hammadi, Z.; Al-Shamsi, A.; Hertecant, J.; Okun, J.G.; Hoffmann, G.F.; et al. Assessment of methylcitrate and methylcitrate to citrate ratio in dried blood spots as biomarkers for inborn errors of propionate metabolism. Sci. Rep. 2019, 9, 12366. [Google Scholar] [CrossRef]
- Sadun, A.A. Mitochondrial optic neuropathies. J. Neurol. Neurosurg. Psychiatry 2002, 72, 423–425. [Google Scholar]
- Pinar-Sueiro, S.; Martínez-Fernández, R.; Lage-Medina, S.; Aldamiz-Echevarria, L.; Vecino, E. Optic neuropathy in methylmalonic acidemia: The role of neuroprotection. J. Inherit. Metab. Dis. 2010, 33, S199–S203. [Google Scholar] [CrossRef]
- Traber, G.; Baumgartner, M.R.; Schwarz, U.; Pangalu, A.; Donath, M.Y.; Landau, K. Subacute bilateral visual loss in methylmalonic acidemia. J. Neuroophthalmol. 2011, 31, 344–346. [Google Scholar] [CrossRef] [PubMed]
- Montero, R.; Yubero, D.; Salgado, M.C.; González, M.J.; Campistol, J.; O’Callaghan, M.D.M.; Pineda, M.; Delgadillo, V.; Maynou, J.; Fernandez, G.; et al. Plasma coenzyme Q10 status is impaired in selected genetic conditions. Sci. Rep. 2019, 9, 793. [Google Scholar] [CrossRef] [PubMed]
- García-Corzo, L.; Luna-Sánchez, M.; Doerrier, C.; Ortiz, F.; Escames, G.; Acuña-Castroviejo, D.; López, L.C. Ubiquinol-10 ameliorates mitochondrial encephalopathy associated with CoQ deficiency. Biochim. Biophys. Acta 2014, 1842, 893–901. [Google Scholar] [CrossRef]
- Salviati, L.; Trevisson, E.; Rodriguez Hernandez, M.A.; Casarin, A.; Pertegato, V.; Doimo, M.; Cassina, M.; Agosto, C.; Desbats, M.A.; Sartori, G.; et al. Haploinsufficiency of COQ4 causes coenzyme Q10 deficiency. J. Med. Genet. 2012, 49, 187–191. [Google Scholar] [CrossRef]
- López-Lluch, G.; Del Pozo-Cruz, J.; Sánchez-Cuesta, A.; Cortés-Rodríguez, A.B.; Navas, P. Bioavailability of coenzyme Q10 supplements depends on carrier lipids and solubilization. Nutrition 2019, 57, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Buján, N.; Arias, A.; Montero, R.; García-Villoria, J.; Lissens, W.; Seneca, S.; Espinós, C.; Navas, P.; De Meirleir, L.; Artuch, R.; et al. Characterization of CoQ10 biosynthesis in fibroblasts of patients with primary and secondary CoQ10 deficiency. J. Inherit. Metab. Dis. 2014, 37, 53–62. [Google Scholar] [CrossRef]
- Yubero, D.; Montero, R.; Martín, M.A.; Montoya, J.; Ribes, A.; Grazina, M.; Trevisson, E.; Rodriguez-Aguilera, J.C.; Hargreaves, I.P.; Salviati, L.; et al. Secondary coenzyme Q10 deficiencies in oxidative phosphorylation (OXPHOS) and non-OXPHOS disorders. Mitochondrion 2016, 30, 51–58. [Google Scholar] [CrossRef]
- Tuncel, A.T.; Boy, N.; Morath, M.A.; Hörster, F.; Mütze, U.; Kölker, S. Organic acidurias in adults: Late complications and management. J. Inherit. Metab. Dis. 2018, 41, 765–776. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Sex | Age at the Beginning of the Study | Age at Diagnosis | Genetics | Clinical Course during the Study | Clinical Course (Long-Term Complications) |
|---|---|---|---|---|---|
| M | 7 | neonatal | PCCB gene p.Gly407Argfs*14/p.Arg410Trp | - | Severe neuromotor delay, leukopenia Severe persistent anemia |
| M | 3 | neonatal | PCCA gene p.Leu470Arg/p.Leu470Arg | Bacteriemia related to vascular central catheter | Severe neuromotor delay Choreoathetosis, basal ganglia involvement, leukopenia, frequent infections, dilated cardiomyopathy, pancreatitis Severe persistent anemia |
| F | 10 | 4 months | PCCA gene p.Gly477fs*9/p.Cys616_Val633del | Mild pancreatitis | Pancreatitis |
| M | 5 | Neonatal screening | PCCB p.Asn536Asp/p.Asn536Asp | - | Autism |
| F | 22 | 4 months | PCCB gene p.Gly407Argfs*14/p.Glu168Lys | - | - |
| M | 7 | 6 months | PCCA gene p.Gly477fs*9/p.Cys616_Val633del | - | Sever neuromotor delay |
| F | 13 | 6 months | PCCB gene p.Arg512Cys/p.Gly255Ser | Mild hyperammonemia due to dietary transgression | Neuromotor delay, epilepsy, pancreatitis, myositis |
| Plasma CoQ10 (NV: 0.91 ± 0.35 μmol/L) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Time | 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| Patient | ||||||||
| Baseline | 0.43 | 0.68 | 0.51 | 0.3 | 0.31 | 0.41 | 0.41 | |
| 3 months | 7.6 | 1.38 | 1.36 | 4.29 | 5.11 | 0.78 | 2.77 | |
| 6 months | 4.35 | 0.85 | 0.62 | 0.38 | 1.86 | 2.22 | ||
| N Median (25th Percentile; 75th Percentile) | |||
|---|---|---|---|
| Baseline | 3 Months | 6 Months | |
| CoQ10 (μmol/L) NV: 0.91 ± 0.35 | N = 7 0.41 (0.36; 0.47) | N = 7 2.77 (1.37; 4.7) | N = 6 1.35 (0.67; 2.13) |
| Glutamine (μmol/L) NV: 440 ± 99 | N = 7 387 (320; 415) | N = 6 439 (339; 570) | N = 7 381 (327; 413) |
| Alanine (μmol/L) NV: 283 ± 88 | N = 7 326 (285; 350) | N = 6 332 (262; 376) | N = 7 334 (282; 445) |
| Lysine (μmol/L) NV: 132 ± 32 | N = 7 118 (101.5; 155) | N = 6 162 (143; 179) | N = 7 144 (124.5; 159) |
| Glycine (μmol/L) NV: 209 ± 66 | N = 7 744 (353.5; 925.5) | N = 6 525 (280; 1061.5) | N = 7 746 (404; 859) |
| Lactate (mmol/mol creat) NV: 1–113 | N = 6 31.5 (21.5; 47) | N = 6 25 (15; 30.5) | N = 7 31 (23; 89) |
| 3-OH-Propionic acid (mmol/mol creat) NV: 5–27 | N = 6 168.5 (87; 646) | N = 6 181.5 (114; 396) | N = 7 337 (210; 578.5) |
| α-Ketoglutarate (mmol/mol creat) NV: 17–492 | N = 6 35.5 (32; 186) | N = 6 60.5 (45.5; 220) | N = 7 90 (60.5; 137.5) |
| Citrate (mmol/mol creat) NV: 205–1735 | N = 6 1204.5 (739; 1809) | N = 6 2110.5 (1522.5; 2479.5) | N = 7 3412 (3042; 3859) |
| Methylcitrate (mmol/mol creat) NV: 1–13 | N = 5 516 (509; 811) | N = 5 635 (127; 676) | N = 7 530 (270; 906.5) |
| Citrate/methylcitrate | N = 5 2.2 (2.1; 3) | N = 5 6.9 (1.8; 11.4) | N = 7 5.4 (3.8; 9.5) |
| Fumarate (mmol/mol creat) NV: 0–17 | N = 6 8 (6; 50) | N = 6 6 (1.5; 20) | N = 7 25 (9; 56) |
| Malate (mmol/mol creat) NV: 0–35 | N = 6 7.5 (5.2; 81) | N = 6 15 (8; 20) | N = 7 29 (11; 72) |
| Succinate (mmol/mol creat) NV: 3–80 | N = 5 35 (16; 39) | N = 6 50 (35; 62) | N = 6 49 (49; 50) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanescu, S.; Belanger-Quintana, A.; Fernández-Felix, B.M.; Ruiz-Sala, P.; Alcaide, P.; Arrieta, F.; Martínez-Pardo, M. Plasma CoQ10 Status in Patients with Propionic Acidaemia and Possible Benefit of Treatment with Ubiquinol. Antioxidants 2022, 11, 1588. https://doi.org/10.3390/antiox11081588
Stanescu S, Belanger-Quintana A, Fernández-Felix BM, Ruiz-Sala P, Alcaide P, Arrieta F, Martínez-Pardo M. Plasma CoQ10 Status in Patients with Propionic Acidaemia and Possible Benefit of Treatment with Ubiquinol. Antioxidants. 2022; 11(8):1588. https://doi.org/10.3390/antiox11081588
Chicago/Turabian StyleStanescu, Sinziana, Amaya Belanger-Quintana, Borja Manuel Fernández-Felix, Pedro Ruiz-Sala, Patricia Alcaide, Francisco Arrieta, and Mercedes Martínez-Pardo. 2022. "Plasma CoQ10 Status in Patients with Propionic Acidaemia and Possible Benefit of Treatment with Ubiquinol" Antioxidants 11, no. 8: 1588. https://doi.org/10.3390/antiox11081588
APA StyleStanescu, S., Belanger-Quintana, A., Fernández-Felix, B. M., Ruiz-Sala, P., Alcaide, P., Arrieta, F., & Martínez-Pardo, M. (2022). Plasma CoQ10 Status in Patients with Propionic Acidaemia and Possible Benefit of Treatment with Ubiquinol. Antioxidants, 11(8), 1588. https://doi.org/10.3390/antiox11081588