
An In Vitro and In Silico Perspective Study of Seed Priming with Zinc on the Phytotoxicity and Accumulation Pattern of Arsenic in Rice Seedlings

 ,
,  ,
,  ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Growth Responses and Total Chlorophyll Content
2.3. Acid Digestion for Determination of Total As—Content
2.4. Determination of Hydrogen Peroxide, Superoxide Radical and Lipid Peroxidation
2.5. Enzymatic Antioxidants
2.6. Non-Enzymatic Antioxidants
2.7. Computational Modelling
2.8. Molecular Docking
2.9. Ligands
2.10. Analysis of the Interactions of the Ligands
2.11. Statistics
3. Results
3.1. Zn2+ Supplementation through Seed Priming Effect on Rice Seedlings Grown under As Stress
3.2. Zn2+ Supplementation through the Seed Priming Effect on As-Accumulation in Rice Seedlings
3.3. Zn2+ Supplementation through the Seed Priming Effect on As-Induced Oxidative Stress in Rice Seedlings
3.4. Zn2+ Supplementation through the Seed Priming Effect on Antioxidants Metabolism in Rice Seedlings
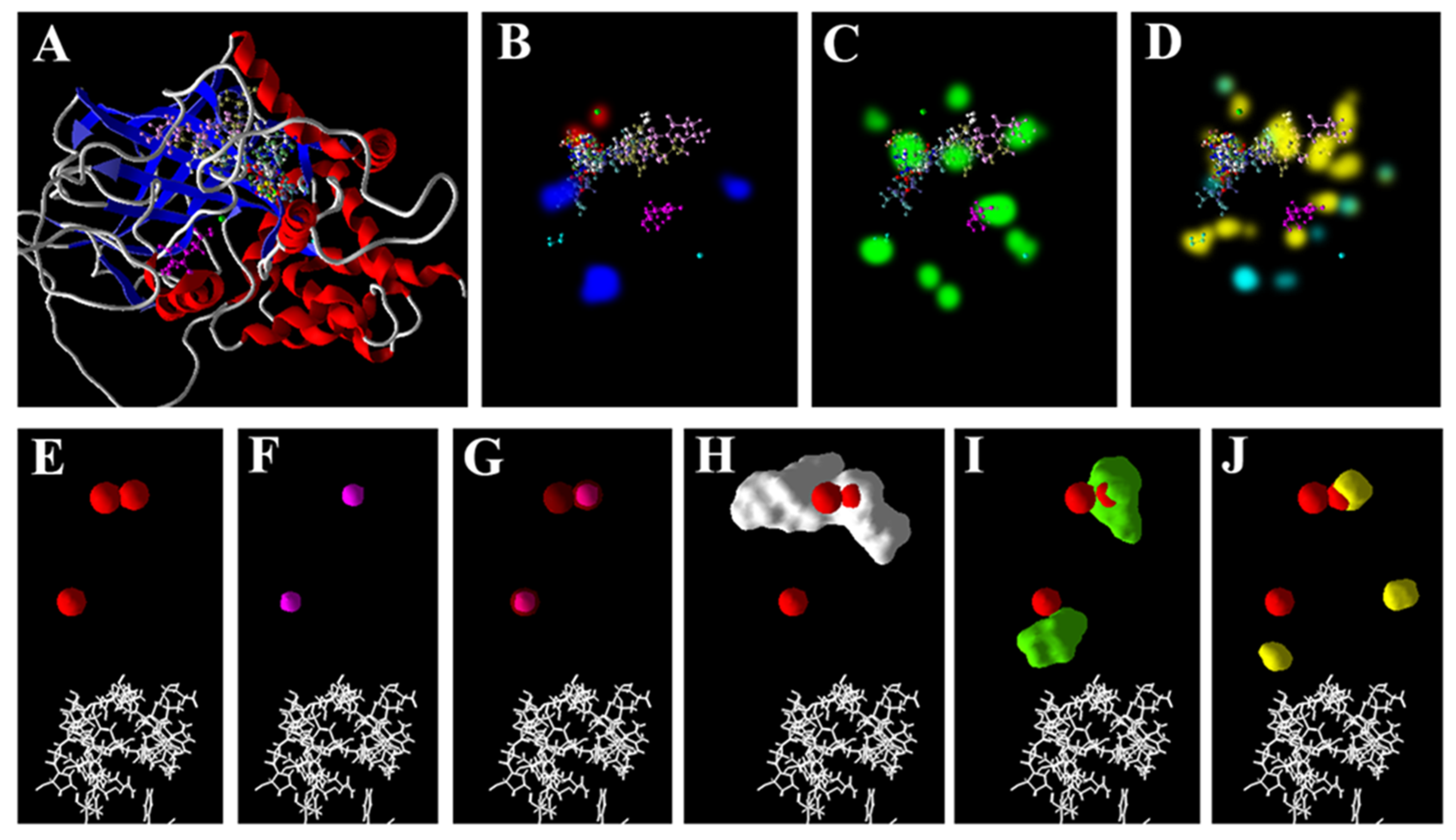
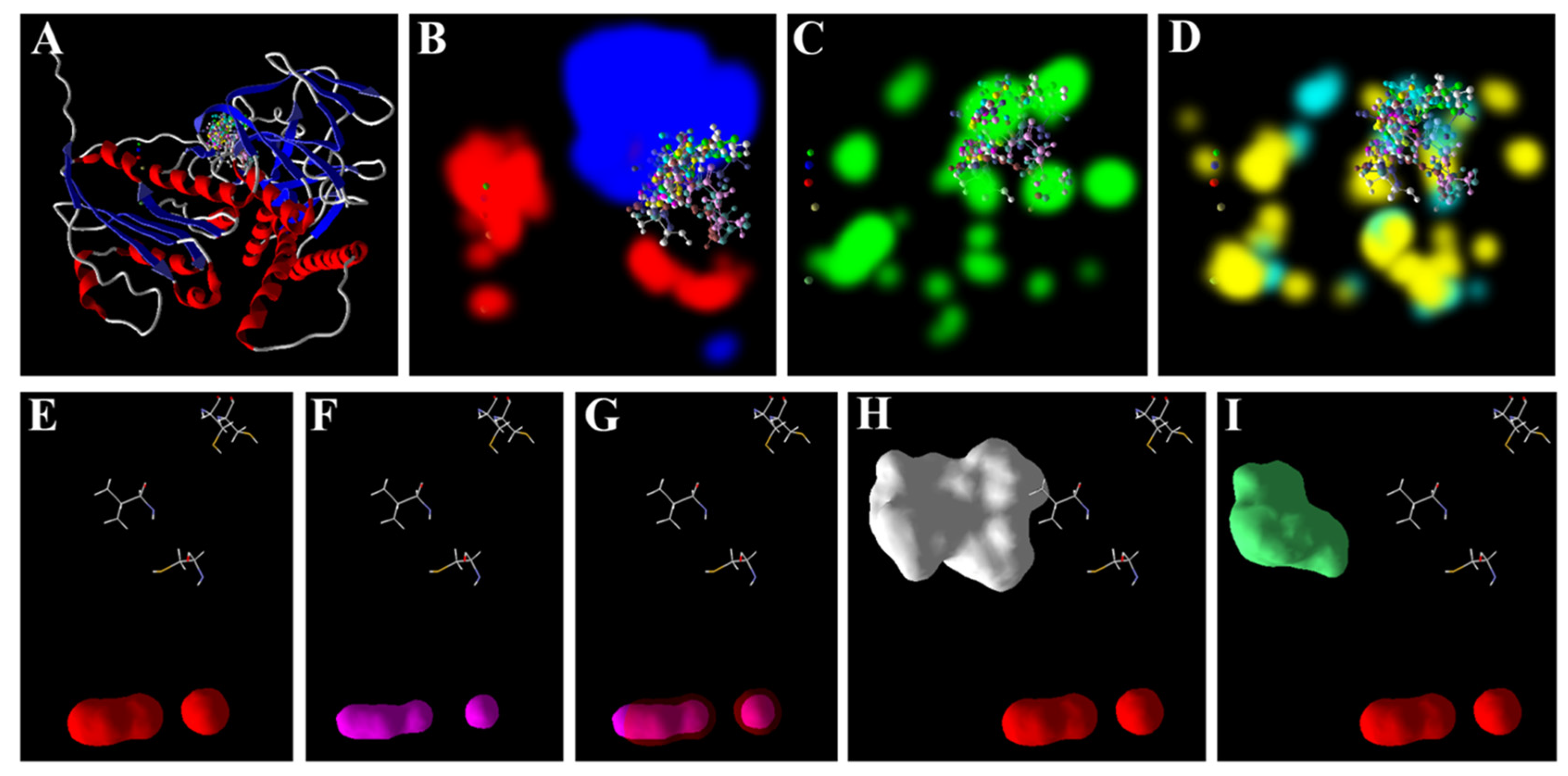
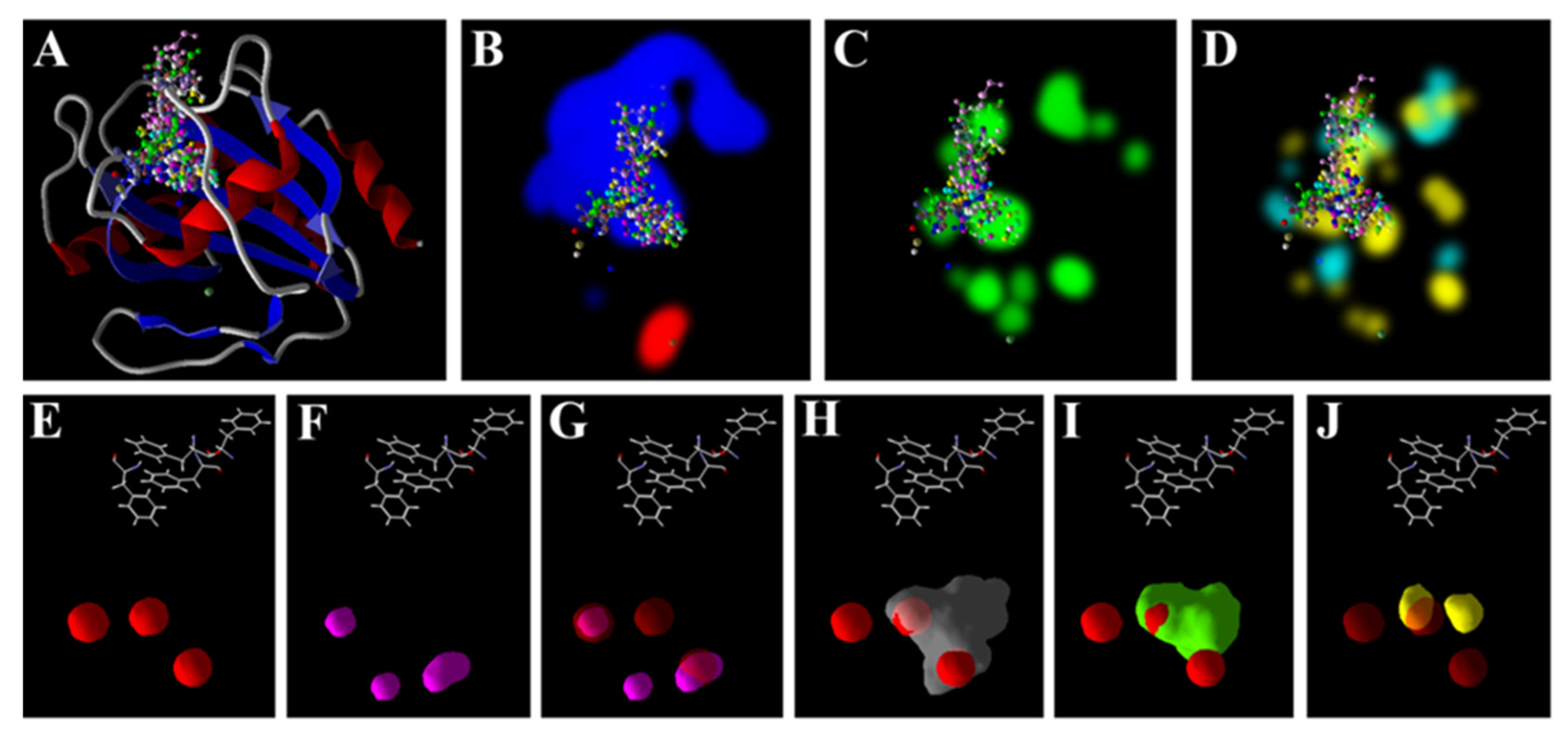
3.5. Zn2+ Supplementation through the Seed Priming Effect on Docking Scores and Their Interactions
4. Discussion
4.1. Effect of Zn2+ Supplementation on As-Induced Morpho-Physiological Responses and As Accumulation
4.2. Effect of Zn2+ Supplementation on As-Induced ROS Accumulation, Oxidative Stress and Antioxidant Metabolism in Rice Seedlings
4.3. Molecular Docking Analysis of Zn × As Interaction by Targeting Antioxidant Enzymes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moulick, D.; Samantha, S.; Sarkar, S.; Mukherjee, A.; Patnayak, B.; Saha, S.; Awasthi, J.P.; Bhoumick, S.; Ghosh, D.; Shyman, A.C.; et al. Arsenic contamination, impact and mitigation strategies in rice agro-ecosystem: An inclusive insight. Sci. Total Environ. 2021, 800, 149477. [Google Scholar] [CrossRef] [PubMed]
- Barringer, J.L.; Reilly, P.A. Arsenic in ground water: A summary of sources and the biogeochemical and hydrogeologic factors affecting arsenic occurrence and mobility. In Current Perspective in Contaminant Hydrology and Water Resources Sustainability; Bradley, P.M., Ed.; Intech Open: Singapore, 2013; p. 83. [Google Scholar]
- Bhattacharya, P.; Adhikari, S.; Samal, A.C.; Das, R.; Dey, D.; Deb, A. Health risk assessment of co-occurrence of toxic fluoride and arsenic in ground water of Dharmanagar region of North Tripura (India). Sustain. Dev. 2020, 11, 100430. [Google Scholar]
- Wang, X.; Peng, B.; Tan, C.; Ma, L.; Rathinasabapathi, D. Recent advances in arsenic bioavailability, transport and speciation in rice. Environ. Sci. Pollut. Res. Int. 2015, 22, 5742–5750. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, J.; Song, W.Y. Arsenic uptake and translocation in plants. Plant Cell Physiol. 2016, 57, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Wailes, E.J. Rice: Global trade, protectionist’s policies and the impact of trade liberalization. In Global Agricultural Trade and Developing Countries; Aksoy, M.A., Beghin, J.C., Eds.; The World Bank: Washington, DC, USA, 2005. [Google Scholar]
- Dittmar, J.; Voegelin, A.; Maurer, F.; Roberts, L.C.; Hug, S.J.; Saha, G.C.; Ali, M.A.; Badruzzaman, A.B.M.; Kretzschmar, R. Arsenic in soil and irrigation water affects arsenic uptake in rice: Complementary insight from field and pot studies. Environ. Sci. Technol. 2010, 44, 8842–8848. [Google Scholar] [CrossRef]
- Duan, G.L.; Liu, W.J.; Chen, X.P.; Hua, Y.; Zhu, Y.G. Association of arsenic with nutrient elements in rice plants. Metalomics 2013, 5, 784–792. [Google Scholar] [CrossRef]
- Meharg, A.A. Arsenic in rice-understanding a new disaster for South—East Asia. Trend Plant Sci. 2004, 9, 415–417. [Google Scholar] [CrossRef]
- Neumann, R.B.; Ashfaque, K.N.; Badruzzaman, A.B.M.; Ali, M.A.; Shoemaker, J.K.; Harvey, C.F. Anthropogenic influences on groundwater arsenic concentrations in Bangladesh. Nat. Geosci. 2011, 3, 46–52. [Google Scholar] [CrossRef]
- Moulick, D.; Ghosh, D.; Santra, S.C. Evaluation of effectiveness of seed priming with selenium in rice during germination under arsenic stress. Plant Physiol. Biochem. 2016, 109, 571–578. [Google Scholar] [CrossRef]
- Moulick, D.; Santra, S.C.; Ghosh, D. Consequences of paddy cultivation in arsenic-contaminated paddy fields of lower Indo-Gangetic Plain on arsenic accumulation pattern and selected grain quality traits: A preliminary assessment. In Mechanisms of Arsenic Toxicity and Tolerance in Plants; Hasanuzzaman, M., Nahar, K., Fujita, M., Eds.; Springer: Singapore, 2018; pp. 49–78. [Google Scholar]
- Finnegan, P.M.; Chen, W. Arsenic toxicity: The effects on plant metabolism. Fornt. Physiol. 2012, 3, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, I. Arsenic induced oxidative stress in plants. Biologia 2012, 67, 447–453. [Google Scholar] [CrossRef]
- Nath, S.; Panda, P.; Mishra, S.; Dey, M.; Choudhury, S.; Sahoo, L.; Panda, S.K. Arsenic stress in rice: Oxidative stress and redox regulation by iron. Plant Physiol. Biochem. 2014, 80, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Singh, S.; Parihar, P.; Singh, V.P.; Prasad, S.M. Arsenic contamination, consequences and remediation techniques: A review. Ecotoxicol. Environ. Saf. 2015, 112, 247–270. [Google Scholar] [CrossRef]
- Moulick, D.; Santra, S.C.; Ghosh, D.; Panda, S.K. An assessment of efficiency of zinc priming in Rice (cv. MTU-7029) during germination and early seedling growth. In Priming and Pre-Treatment of Seeds and Seedlings; Hasanuzzaman, M., Fotopoulos, V., Eds.; Springer: Singapore, 2019; pp. 495–507. [Google Scholar]
- Wenzel, W.; Brandstetter, A.; Wutte, H.; Lombi, E.; Prohaska, T.; Stingeder, G.; Adriano, D. Arsenic in field collected soil solutions and extracts of contaminated soils and its implication to soil standards. J. Plant Nutr. Soil Sci. 2002, 165, 221–228. [Google Scholar] [CrossRef]
- Takahashi, Y.; Keiko, R.M.; Kurishima, K.; Kihou, N.; Yuita, K. Arsenic behavior in paddy fields during the cycle of flooded and non-flooded period. Environ. Sci. Technol. 2004, 38, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Xu, X.-Y.; Su, Y.-H.; McGrath, S.P.; Zhao, F.-J. Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proc. Natl. Acad. Sci. USA 2008, 105, 9931–9935. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.H.; McGrath, S.P.; Zhao, F.J. Rice is more efficient in arsenite uptake and translocation than wheat and barley. Plant Soil 2010, 328, 27–34. [Google Scholar] [CrossRef]
- Tripathi, R.D.; Srivastava, S.; Mishra, S.; Singh, N.; Tuli, R.; Gupta, D.K.; Maathuis, F.J.M. Arsenic hazard: Strategies for tolerance and remediation by plants. Trends Biotechnol. 2007, 25, 158–165. [Google Scholar] [CrossRef]
- Maity, J.P.; Kar, S.; Liu, J.-H.; Jean, J.-S.; Chen, C.-Y.; Bundschuh, J.; Santra, S.C.; Liu, C.-C. The potential reductive mobilization of arsenic [As(V) to As(III)] by OSBH(2) (Pseudomonas stutzeri) and OSBH(5) (Bacillus cereus) in an oil contaminated site. J. Environ. Health Tox. Hazard. Subst. Environ. Eng. 2011, 46, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Dhankher, O.P.; Rosen, B.P.; McKinney, E.C.; Meagher, R.B. Hyperaccumulation of arsenic in shoot of Arabidopsis silenced for arsenate reductase (ACR2). Proc. Natl. Acad. Sci. USA 2006, 103, 5413–5418. [Google Scholar] [CrossRef] [Green Version]
- Requejo, R.; Tena, M. Proteome analysis of maize roots reveals that oxidative stress is a main contributing factor to plant arsenic toxicity. Phytochemistry 2005, 66, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Moulick, D.; Ghosh, D.; Santra, S.C. An assessment of some physicochemical properties and cooking characteristics of milled rice and associated health risk in two rice varieties of arsenic contaminated areas of West Bengal, India. Int. J. Res. Agric. Food Sci. 2016, 6, 44–55. [Google Scholar]
- Hartley-Whitaker, J.; Ainsworth, G.; Vooijs, R.; Bookum, W.T.; Schat, H.; Meharg, A.A. Phytochelatin are involved in differential arsenate tolerance in Holcus lanatus. Plant Physiol. 2001, 126, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abedin, M.J.; Meharg, A.A. Relative toxicity of arsenite and arsenate on germination and seedling growth of rice (Oryza sativa L.). Plant Soil 2002, 243, 57–66. [Google Scholar] [CrossRef]
- Garg, N.; Singla, P. Arsenic toxicity in crop plants: Physiological effects and tolerance mechanisms. Environ. Chem. Lett. 2011, 9, 303–321. [Google Scholar] [CrossRef]
- Awasthi, S.; Chauhan, R.; Srivastava, S.; Tripathi, R.D. The journey of arsenic from soil to grain in rice. Front. Plant Sci. 2017, 8, 1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meharg, A.A.; Williams, P.N.; Adomako, E.; Lawgali, Y.Y.; Deacon, D.; Villada, A.; Cambell, R.C.J.; Sun, G.; Zhu, Y.G.; Feldmann, J.; et al. Geographical variation in total and inorganic arsenic content of polished (white) rice. Environ. Sci. Technol. 2009, 43, 1612–1617. [Google Scholar] [CrossRef]
- Carey, A.-M.; Scheckel, K.G.; Lombi, E.; Newville, M.; Choi, Y.; Norton, G.J.; Charnock, J.M.; Feldmann, J.; Price, A.H.; Meharg, A. Grain unloading of arsenic species in rice. Plant Physiol. 2010, 152, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed priming for abiotic stress tolerance: An overview. Acta Physiol. Plant. 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.; Langenbach, C.J.; Jaskiewicz, M.R. Priming for enhanced defense. Annu. Rev. Phytopathol. 2015, 53, 97–119. [Google Scholar] [CrossRef]
- Johnson, R.; Puthur, J.T. Seed priming as a cost-effective technique for developing plants with cross tolerance to salinity stress. Plant Physiol. Biochem. 2021, 162, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Lafferty, B.J.; Loeppert, R.H. Methyl arsenic adsorption and desorption behavior on iron oxide. Envrion. Sci. Technol. 2005, 39, 2120–2127. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Nakamura, T.; Dong, D.; Takahashi, Y.; Amachi, S.; Makino, T. Arsenic release from flooded paddy soils is influenced by speciation, Eh, pH and iron dissolution. Chemosphere 2011, 83, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Bakhat, H.F.; Zia, Z.; Fahad, S.; Abbas, S.; Hammad, H.M.; Shahzad, A.N.; Abbas, F.; Alharby, H.; Shahid, M. Arsenic uptake, accumulation and toxicity in rice plants: Possible remedies for its detoxification: A review. Environ. Sci. Pollut. Res. Int. 2017, 24, 9142–9158. [Google Scholar] [CrossRef]
- Moulick, D.; Santra, S.C.; Ghosh, D. Rice seed priming with Se: A novel approach to mitigate As induced adverse consequences on growth, yield, and As load in brown rice. J. Hazard. Mater. 2018, 355, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Moulick, D.; Santra, S.C.; Ghosh, D. Seed priming with Se alleviate As induced phytotoxicity during germination and seedling growth by restricting As translocation in rice (Oryza sativa L. cv IET-4094). Ecotoxicol. Environ. Saf. 2017, 145, 449–456. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Yruela, I. Transition Metals in Plant Photosynthesis. Metallomics 2013, 5, 1090–1109. [Google Scholar] [CrossRef] [Green Version]
- Ishimaru, Y.; Bashir, K.; Nishizawa, N.K. Zn uptake and translocation in rice plants. Rice 2011, 4, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Gibson, R.S. Zinc deficiency and human health: Etiology, health consequences, and future solutions. Plant Soil 2012, 361, 291–299. [Google Scholar] [CrossRef]
- Barak, P.; Helmke, P.A. The chemistry of zinc. In Zinc in Soils and Plants; Robson, A.D., Ed.; Springer: Dordrecht, The Netherlands, 1993; pp. 1–13. [Google Scholar]
- Majumder, S.; Datta, K.; Datta, S.K. Rice biofortification: High iron, zinc, and vitamin-A to fight against “hidden hunger”. Agronomy 2019, 9, 803. [Google Scholar] [CrossRef] [Green Version]
- Andresen, E.; Peiter, E.; Kupper, H. Trace metal metabolism in plants. J. Exp. Bot. 2018, 69, 909–954. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.; Moulick, D.; Mazumder, M.K. Secondary metabolites protect against metal and metalloid stress in rice: An in-silico investigation using dehydroascorbate reductase. Acta Physiol. Plant. 2021, 43, 3. [Google Scholar] [CrossRef]
- Choudhury, S.; Mazumder, M.K.; Moulick, D.; Sharma, P.; Tata, S.K.; Ghosh, D.; Ali, H.M.; Siddiqui, M.H.; Brestic, M.; Skalicky, M.; et al. A Computational Study of the Role of Secondary Metabolites for Mitigation of Acid Soil Stress in Cereals Using Dehydroascorbate and Mono-Dehydroascorbate Reductases. Antioxidants 2022, 11, 458. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, M.K.; Sharma, P.; Moulick, D.; Tata, S.K.; Choudhury, S. Salicylic acid ameliorates zinc and chromium-induced stress responses in wheat seedlings: A biochemical and computational analysis. Cereal Res. Commun. 2021. [Google Scholar] [CrossRef]
- Mazumder, M.K.; Moulick, D.; Choudhury, S. Iron (Fe3+)-mediated redox responses and amelioration of oxidative stress in cadmium (Cd2+) stressed mung bean seedlings: A biochemical and computational analysis. J. Plant Biochem. Biotechnol. 2022, 31, 49–60. [Google Scholar] [CrossRef]
- Moulick, D.; Chowardhara, B.; Panda, S.K. Agro-ecotoxicological aspect of arsenic (As) and cadmium (Cd) on Field crops and its mitigation: Current status and future prospect. In Plant-Metal Interactions; Springer: Dordrecht, The Netherlands, 2019; pp. 217–246. [Google Scholar]
- Sagisaka, S. The occurrence of peroxide in a perennial plant, Populus gelrica. Plant Physiol. 1976, 57, 308–309. [Google Scholar] [CrossRef] [Green Version]
- Elstner, E.F.; Heupel, A. Inhibition of nitrate formation from hydroxyl ammonium chloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 746–778. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidase. Method Enzymol. 1955, 2, 276–287. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assay and assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Smith, I.K.; Vierhgeller, T.L.; Throne, C.A. Assay of glutathione reductase in crude tissue homogenate using 5,5′-dithiobis-(2-nitrobenzoic acid). Anal. Biochem. 1988, 175, 408–413. [Google Scholar] [CrossRef]
- Oser, B.L. Hawk’s Physiological Chemistry; Dubois, M., Ed.; McGraw Hill: New Delhi, India, 1979. [Google Scholar]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. High accurate protein structure with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Velankar, S.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Moulick, D.; Ghosh, D.; Skalicky, M.; Gharde, Y.; Mazumder, M.K.; Choudhury, S.; Biswas, J.K.; Santra, S.C.; Brestic, M.; Vachova, P.; et al. Interrelationship Among Rice Grain Arsenic, Micronutrients Content and Grain Quality Attributes: An Investigation From Genotype × Environment Perspective. Front. Environ. Sci. 2022, 10, 857629. [Google Scholar] [CrossRef]
- Verma, S.; Mukherjee, A.; Choudhury, R.; Mahanta, C. Brahmaputra river basin groundwater: Solute distribution, chemical evolution and arsenic occurrences in different geomorphic settings. J. Hydrol. Reg. Stud. 2015, 4, 131–153. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Das, A.; Das, N.; Goswami, R.; Singh, U.K. Co-occurrence perspective of arsenic and fluoride in the groundwater of Diphu, Assam, Northeastern India. Chemosphere 2016, 150, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Rahman, M.M.; Ramanathan, A.L.; Naidu, R. Arsenic and other elements in drinking water and dietary components from the middle Gangetic plain of Bihar, India: Health risk index. Sci. Total Environ. 2016, 539, 125–134. [Google Scholar] [CrossRef]
- Das, N.; Das, A.; Sarma, K.P.; Kumar, M. Provenance, prevalence and health perspective of co-occurrences of arsenic, fluoride and uranium in the aquifers of the Brahmaputra River floodplain. Chemosphere 2018, 194, 755–772. [Google Scholar] [CrossRef] [PubMed]
- Das, N.; Sarma, K.P.; Patel, A.K.; Deka, J.P.; Das, A.; Kumar, A.; Shea, P.J.; Kumar, M. Seasonal disparity in the co-occurrence of arsenic and fluoride in the aquifers of the Brahmaputra flood plains, Northeast India. Environ. Earth Sci. 2017, 76, 183. [Google Scholar] [CrossRef]
- Patel, A.K.; Das, N.; Goswami, R.; Kumar, M. Arsenic mobility and potential coleaching of fluoride from the sediments of three tributaries of the Uer Brahmaputra floodplain, Lakhimpur, Assam, India. J. Geochem. Explor. 2019, 203, 45–58. [Google Scholar] [CrossRef]
- World Bank. Towards a More Effective Operational Response: Arsenic Contamination of Groundwater in South and East Asian Countries; Policy Report; World Bank: Washington, DC, USA, 2005; Volume 1. [Google Scholar]
- Mukherjee, A.; Sengupta, M.K.; Hossain, M.A.; Ahamed, S.; Das, B.; Nayak, B.; Lodh, D.; Rahman, M.M.; Chakraborti, D. Arsenic contamination in groundwater: A global perspective with emphasis on the Asian scenario. J. Health Popul. Nutr. 2006, 24, 142–163. [Google Scholar] [PubMed]
- Chakraborti, D.; Singh, S.K.; Rahman, M.M.; Dutta, R.N.; Mukherjee, S.C.; Pati, S.; Kar, P.B. Groundwater arsenic contamination in the Ganga River Basin: A future health danger. Int. J. Environ. Res. Public Health 2018, 15, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alloway, B.J. Zinc in Soils and Crop Nutrition, 2nd ed.; IZA: Brussels, Belgium; IFA: Paris, France, 2008. [Google Scholar]
- Çakmak, İ.; Kalaycı, M.; Ekiz, H.; Braun, H.J.; Kılınç, Y.; Yılmaz, A. Zinc deficiency as a practical problem in plant and human nutrition in Turkey: A NATO-science for stability project. Field Crops Res. 1999, 60, 75–188. [Google Scholar] [CrossRef] [Green Version]
- Fageria, N.K. Influence of micronutrients on dry matter yield and interaction with other nutrients in annual crops. Pesqui. Agropecuária Bras. 2002, 37, 1765–1772. [Google Scholar] [CrossRef] [Green Version]
- Quijano-Guerta, C.; Kirk, G.J.D.; Portugal, A.M.; Bartolome, V.I.; McLaren, G.C. Tolerance of rice germplasm to zinc deficiency. Field Crops Res. 2002, 76, 123–130. [Google Scholar] [CrossRef]
- Johnson-Beebout, S.E.; Lauren, J.G.; Duxbury, J.M. Immobilization of zinc fertilizer in flooded soils monitored by adapted DTPA soil test. Commun. Soil Sci. Plant Anal. 2009, 40, 1842–1861. [Google Scholar] [CrossRef]
- Beebout, S.J.; Francis, H.C.R.; Dennis, S.J.T.; Ranee, C.M. Reasons for variation in rice (Oryza sativa) grain zinc response to zinc fertilization. In Proceedings of the 3rd International Zinc Symposium, Hyderabad, India, 10–14 October 2011; pp. 10–14. [Google Scholar]
- Zheng, M.; Tao, Y.; Hussain, S.; Jiang, Q.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Seed priming in dry direct-seeded rice: Consequences for emergence, seedling growth and associated metabolic events under drought stress. Plant Growth Regul. 2016, 78, 167–178. [Google Scholar] [CrossRef]
- Mahakham, W.; Sarmah, A.K.; Maensiri, S.; Theerakulpisut, P. Nanopriming technology for enhancing germination and starch metabolism of aged rice seeds using phytosynthesized silver nanoparticles. Sci. Rep. 2017, 7, 8263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zhang, S.; Shan, X.; Zhu, Y.G. Toxicity of arsenate and arsenite on germination, seedling growth and amylolytic activity of wheat. Chemosphere 2005, 61, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wu, F.; Zhou, S.; Yang, J.; Tang, X.; Ye, W. Zinc oxide nanoparticles alleviate the arsenic toxicity and decrease the accumulation of arsenic in rice (Oryza sativa L.). BMC Plant Biol. 2021, 21, 150. [Google Scholar] [CrossRef] [PubMed]
- Stoeva, N.; Berova, M.; Zlatev, Z. Physiological response of maize to arsenic contamination. Biol. Plant. 2003, 47, 449–452. [Google Scholar] [CrossRef]
- Rahman, M.A.; Hasegawa, H.; Rahman, M.M.; Islamm, M.N.; Miah, M.A.M.; Tasmen, A. Effect of arsenic on photosynthesis, growth and yield of five widely cultivated rice (Oryza sativa L.) varieties in Bangladesh. Chemosphere 2007, 67, 1072–1079. [Google Scholar] [CrossRef] [Green Version]
- Das, D.K.; Sur, P.; Das, K. Mobilisation of arsenic in soils and in rice (Oryza sativa L.) plants affected by organic matter and zinc application in irrigation water contaminated with arsenic. Plant Soil Environ. J. 2008, 54, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Das, I.; Sanyal, S.K.; Ghosh, K.; Das, D.K. Arsenic mitigation in soil-plant system through zinc application in West Bengal soils. Bioremediat. J. 2016, 20, 24–37. [Google Scholar] [CrossRef]
- Takahashi, M.A.; Asada, K. Superoxide anion permeability of phospholipid membrane and chloroplast thylakoids. Arch. Biochem. Biophys. 1983, 226, 558–566. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kohli, R.K.; Arora, K. Arsenic-induced root growth inhibition in mung bean (Phaseolus aureus Roxb.) is due to oxidative stress resulting from enhanced lipid peroxidation. Plant Growth Regul. 2007, 53, 65–73. [Google Scholar] [CrossRef]
- Ahmad, P.; Abd_Allah, E.F.; Alyemeni, M.N.; Wijaya, L.; Alam, P.; Bhardwaj, R.; Siddique, K.H. Exogenous application of calcium to 24-epibrassinosteroid pre-treated tomato seedlings mitigates NaCl toxicity by modifying ascorbate–glutathione cycle and secondary metabolites. Sci. Rep. 2018, 8, 13515. [Google Scholar] [CrossRef]
- Ahmad, P.; Abdel Latef, A.A.; Abd_Allah, E.F.; Hashem, A.; Sarwat, M.; Anjum, N.A.; Gucel, S. Calcium and potassium supplementation enhanced growth, osmolyte secondary metabolite production, and enzymatic antioxidant machinery in cadmium-exposed chickpea (Cicer arietinum L.). Front. Plant Sci. 2016, 7, 513. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, E.; Arshad, M.; Khan, M.Z.; Amjad, M.S.; Sadaf, H.M.; Riaz, I.; Sabir, S.; Ahmad, N. Secondary metabolites and their multidimensional prospective in plant life. J. Pharmacog. Phytochem. 2017, 6, 205–214. [Google Scholar]
- Bali, S.; Jamwal, V.L.; Kohli, S.K.; Kaur, P.; Tejpal, R.; Bhalla, V.; Ohri, P.; Gandhi, S.G.; Bhardwaj, R.; Al-Huqail, A.A.; et al. Jasmonic acid application triggers detoxification of lead (Pb) toxicity in tomato through the modifications of secondary metabolites and gene expression. Chemosphere 2019, 235, 734–748. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, R.; Christensen, M.H. MolDock: A new technique for high-accuracy molecular docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
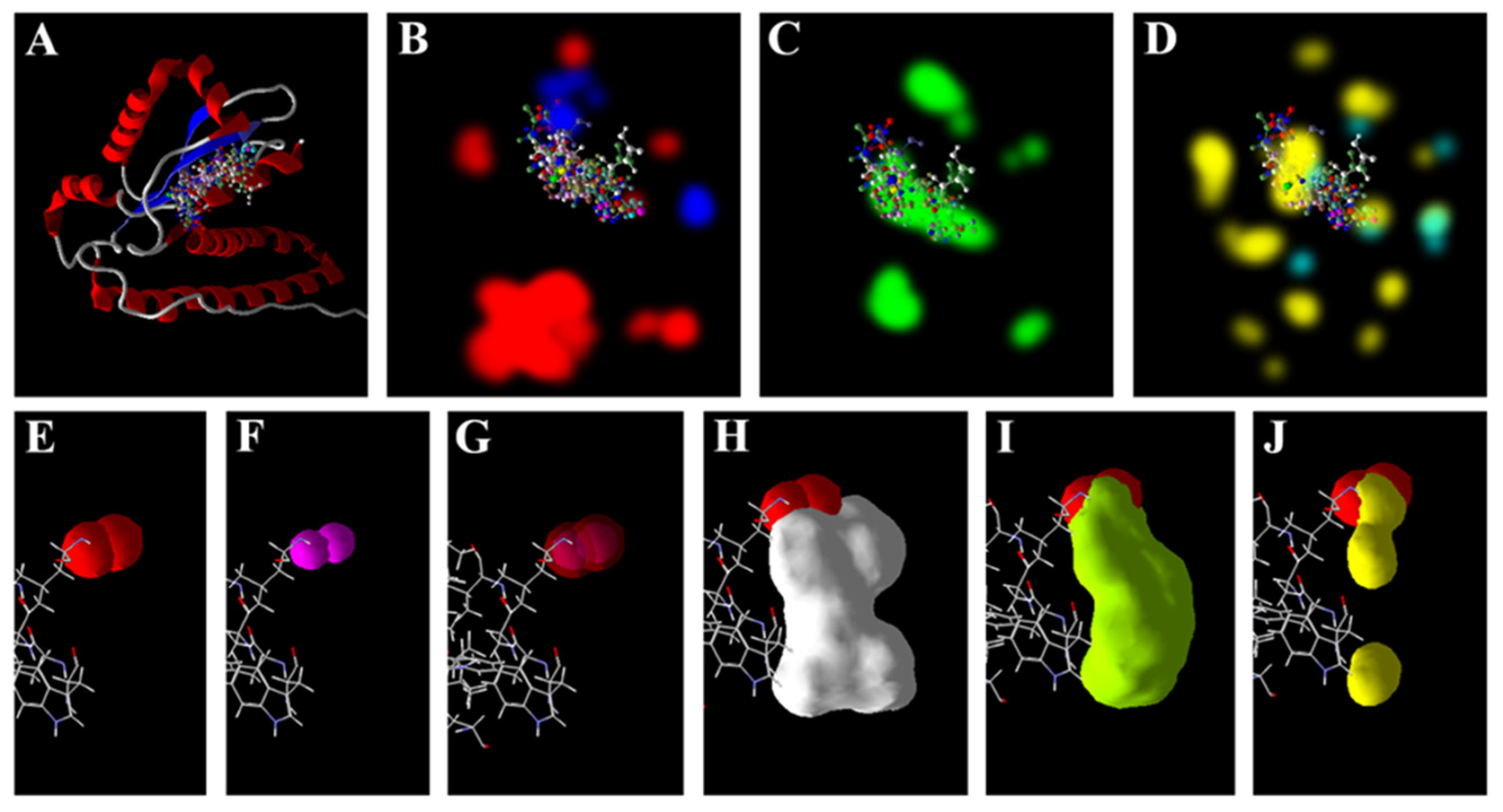

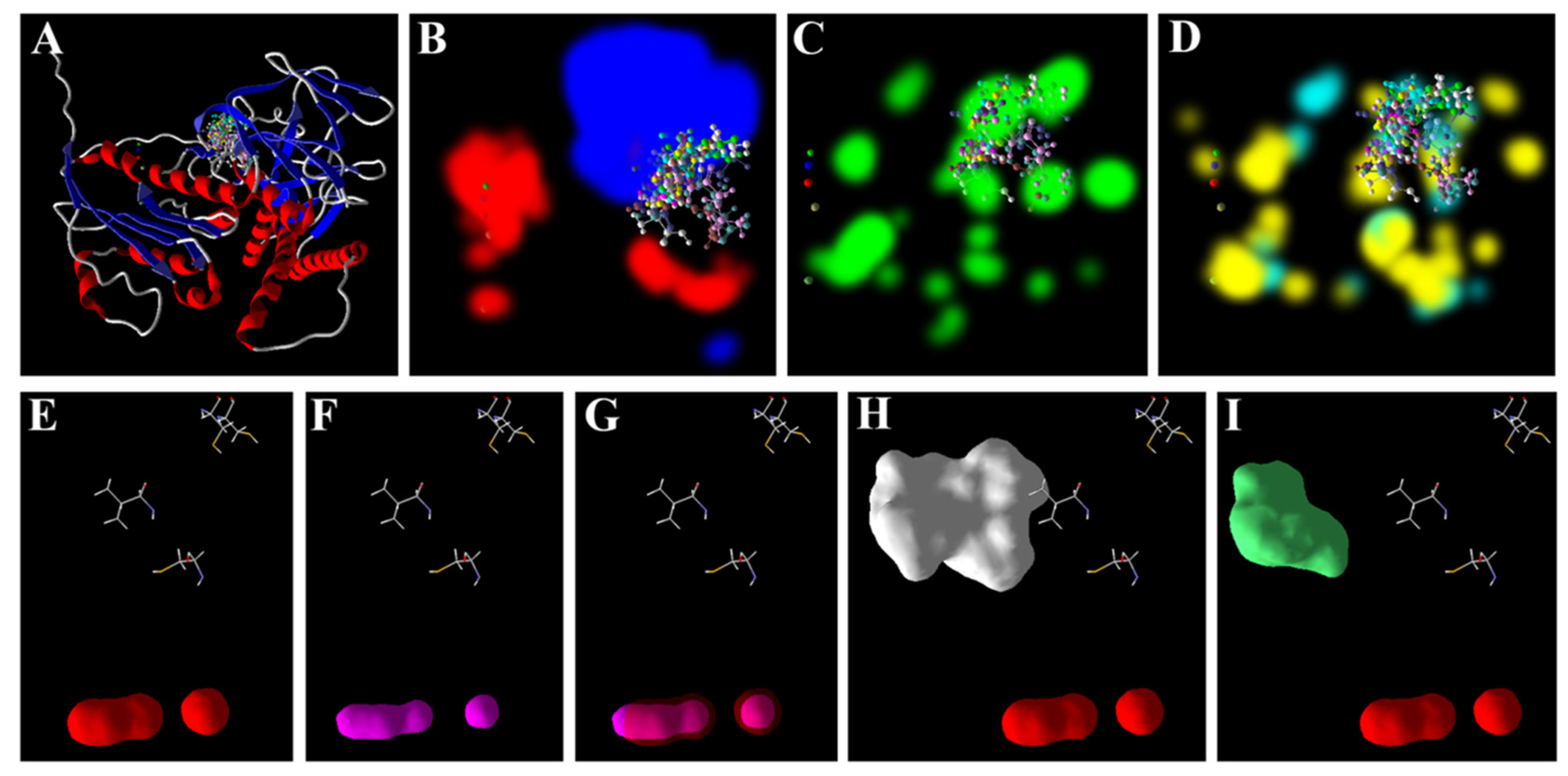
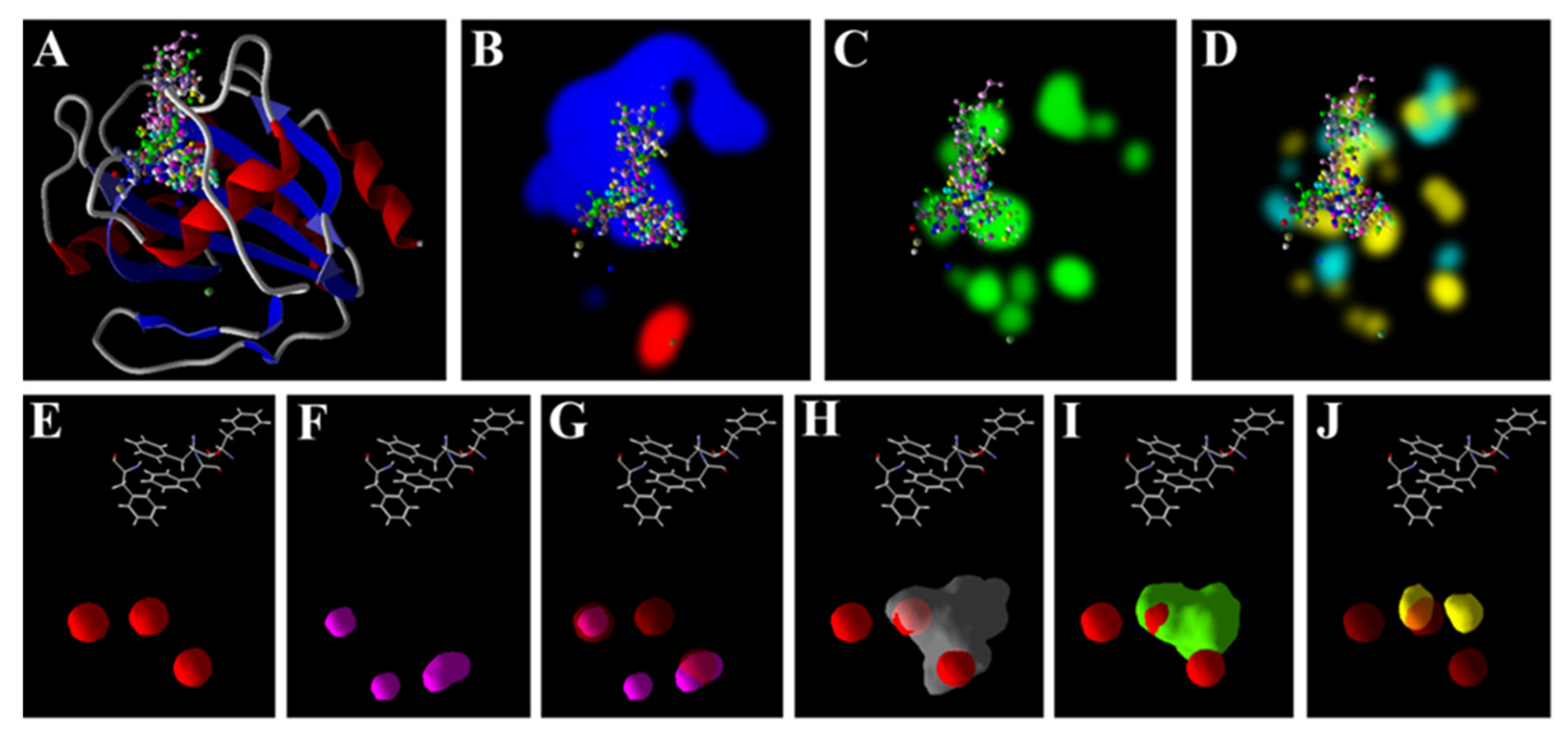

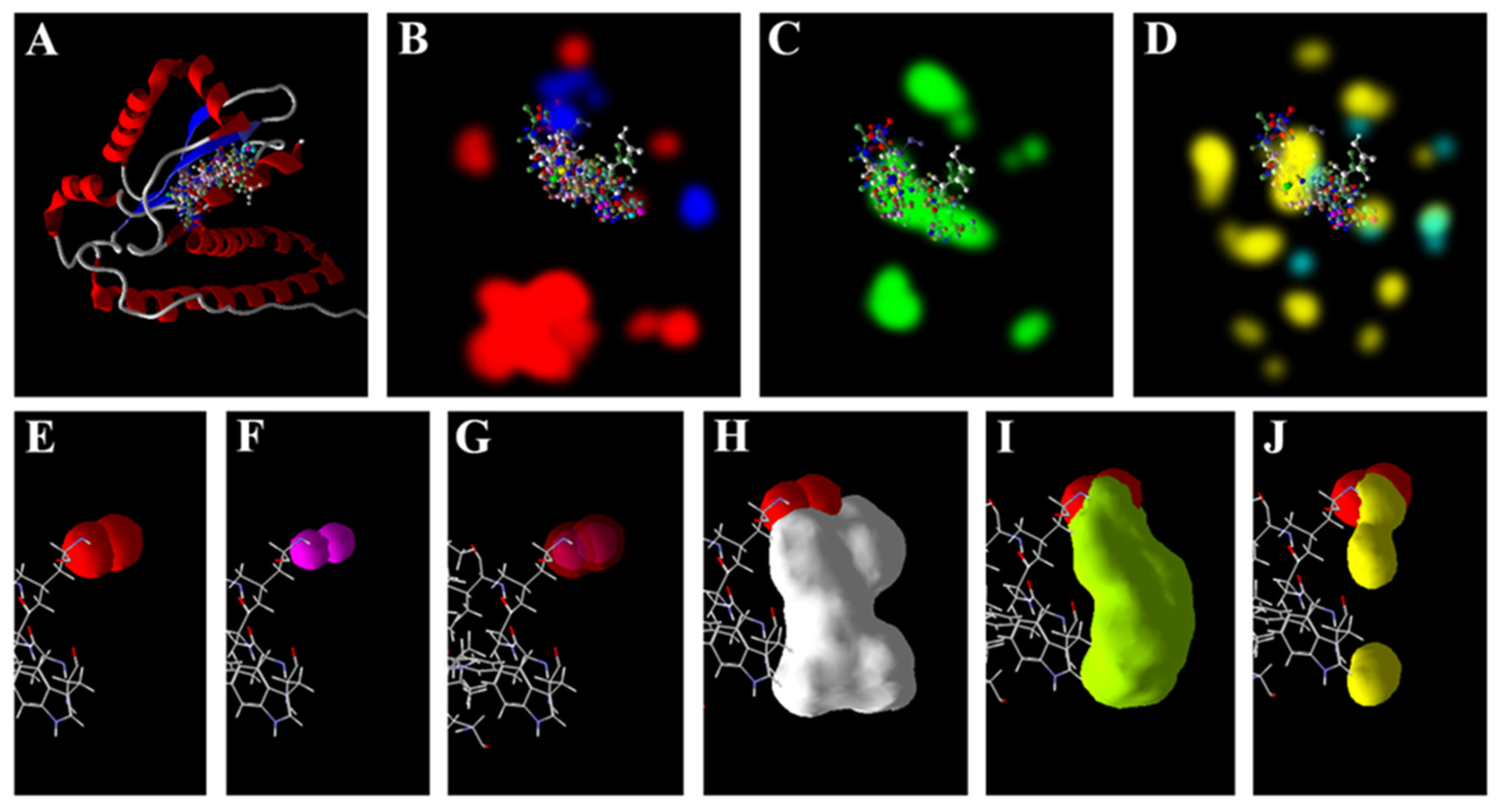{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Elongation (Root) | Elongation (Shoot) | Fresh Weight (Root) | Fresh Weight (Shoot) | Dry Weight (Root) | Dry Weight (Shoot) | Total Chlorophyll | As Content (Root) | As Content (Shoot) |
|---|---|---|---|---|---|---|---|---|---|
| Main Effects | |||||||||
| Variety | |||||||||
| Gomti | 7.58a | 11.40a | 3.21a | 6.96a | 0.016a | 0.307a | 19.5a | 32.3a | 25.6a |
| Kalijeera | 7.15a | 9.95b | 1.81b | 3.76b | 0.018a | 0.138b | 23.4a | 24.1b | 11.4b |
| Seed priming with As | |||||||||
| Control | 9.01a | 14.09a | 3.72a | 7.01a | 0.029a | 0.390a | 29.3a | - | - |
| As | 5.77d | 8.53c | 1.69c | 4.07c | 0.009c | 0.120c | 15.9b | 30.8a | 18.2a |
| As + Zn (0.5 mg L−1) | 6.93c | 9.42c | 2.15b | 5.02b | 0.015b | 0.170bc | 19.2b | 28.8ab | 20.4a |
| As + An (1.0 mg L−1) | 7.73b | 10.65b | 2.49b | 5.33b | 0.015b | 0.209b | 21.4b | 250b | 170a |
| Interaction Effects | |||||||||
| Gomti × Control | 10.12a | 15.58a | 4.22a | 8.21a | 0.034a | 0.596a | 24.2ab | - | - |
| Gomti ×As | 6.08cd | 8.30d | 2.58bc | 5.98c | 0.006d | 0.151bc | 12.5b | 38.3a | 23.9a |
| Gomti × As × Zn (0.5 mg L−1) | 6.72bcd | 9.80cd | 2.90b | 6.51bc | 0.011cd | 0.200bc | 19.7b | 32.8ab | 29.8a |
| Gomti × As × Zn (1.0 mg L−1) | 7.38bc | 11.92bc | 3.16b | 7.12ab | 0.013cd | 0.281b | 21.5b | 26.0bc | 23.2a |
| Kalijeera × Control | 7.90b | 12.60b | 3.22ab | 5.80c | 0.025ab | 0.184bc | 34.3a | - | - |
| Kalijeera × As | 5.46d | 8.76d | 0.80e | 2.17e | 0.013cd | 0.089c | 19.3b | 23.3c | 12.6b |
| Kalijeera × As × Zn (0.5 mg L−1) | 7.14bc | 9.04d | 1.41de | 3.54d | 0.018bc | 0.141c | 18.6b | 24.9bc | 10.9b |
| Kalijeera × As × Zn (1.0 mg L−1) | 8.08b | 9.38d | 1.82cd | 3.54d | 0.016bcd | 0.136c | 21.2b | 24.0bc | 10.8b |
| ANOVA | |||||||||
| Source of variation | |||||||||
| ANOVA p Values | |||||||||
| Variety | 0.1010 | 0.0007 | <0.0001 | <0.0001 | 0.2881 | <0.0001 | 0.0524 | <0.0001 | <0.0001 |
| Seed priming | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0002 | 0.0323 | 0.1754 |
| Variety × seed priming | 0.0012 | 0.0130 | 0.3651 | 0.0299 | 0.0019 | <0.0001 | 0.1312 | 0.0161 | 0.0837 |
| Treatment | H2O2 (Root) | H2O2 (Shoot) | O2●− (Root) | O2●− (Shoot) | MDA (Root) | MDA (Shoot) | CAT (Root) | CAT (Shoot) | GPX (Root) | GPX (Shoot) | SOD (Root) | SOD (Shoot) | GR (Root) | GR (Shoot) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Main Effects | ||||||||||||||
| Variety | ||||||||||||||
| Gomti | 48.9b | 44.2b | 49.5a | 39.8a | 65.3b | 40.4a | 42.3a | 51.2a | 10.73a | 8.73a | 108.7a | 102.9a | 7.36a | 7.41b |
| Kalijeera | 80.1a | 64.4a | 45.5a | 42.6a | 127.7a | 42.1a | 45.6a | 45.8a | 4.43bb | 4.39b | 71.7b | 77.7b | 6.41b | 12.48a |
| Seed priming | ||||||||||||||
| Control | 16.9d | 11.5c | 11.2c | 9.3c | 27.5d | 13.2d | 47.8b | 50.2b | 7.13b | 8.17ab | 99.2b | 116.4a | 9.73a | 14.76a |
| As | 107.1a | 82.5a | 71.7a | 64.9a | 175.0a | 75.1a | 19.8c | 21.4c | 2.34c | 2.13c | 42.4c | 44.0c | 2.73c | 4.81c |
| As + Zn (0.5 mg L−1) | 72.4b | 63.1b | 55.7b | 47.5b | 103.5b | 41.5b | 50.9ab | 58.4a | 9.72a | 7.11b | 102.6b | 100.3b | 7.00b | 9.90b |
| As + An (1.0 mg L−1) | 61.6c | 60.1b | 51.4b | 43.2b | 80.0c | 35.1c | 57.2a | 63.8a | 11.13a | 8.85a | 116.7a | 100.5b | 8.08b | 10.32b |
| Interaction effects | ||||||||||||||
| Gomti × Control | 17.5de | 12.4c | 9.1d | 7.9d | 24.1c | 14.8c | 28.5cd | 37.6de | 7.13c | 9.69b | 100.2b | 112.7b | 9.69a | 14.29a |
| Gomti × As | 95.5b | 77.1a | 86.8a | 70.9a | 134.1b | 70.6a | 16.1d | 20.5e | 2.68d | 2.16d | 43.7cd | 37.2e | 1.89d | 2.22b |
| Gomti × As x Zn (0.5 mg L−1) | 43.7c | 48.6b | 59.3bc | 45.1bc | 58.7c | 42.4b | 53.6ab | 64.6ab | 14.84b | 10.17ab | 131.7a | 128.1ab | 8.12ab | 6.14b |
| Gomti × As × Zn (1.0 mg L−1) | 38.9cd | 38.5b | 42.8c | 35.5c | 44.4c | 33.6b | 70.8a | 81.9a | 18.27a | 12.91a | 159.2a | 133.5a | 9.74a | 6.99b |
| Kalijeera × Control | 16.4e | 10.5c | 13.3d | 10.7d | 31.0c | 11.6c | 67.1a | 62.9bc | 7.12c | 6.64c | 98.2b | 120.0ab | 9.78a | 15.23a |
| Kalijeera × As | 118.6a | 88.0a | 56.7bc | 59.0ab | 216.0a | 79.6a | 23.4d | 22.4e | 2.01d | 2.10d | 41.1d | 50.9de | 3.57cd | 7.39b |
| Kalijeera × As × Zn (0.5 mg L−1) | 101.0ab | 77.5a | 52.1bc | 49.8b | 148.3b | 40.6b | 48.2b | 52.2bcd | 4.60cd | 4.05cd | 73.4bc | 72.6c | 5.88bc | 13.67a |
| Kalijeera × As ×Zn (1.0 mg L−1) | 84.2b | 81.6a | 59.9b | 51.0b | 115.6b | 36.6b | 43.5bc | 45.6cd | 3.99cd | 4.78cd | 74.1b | 67.4cd | 6.42bc | 13.65a |
| ANOVA | ||||||||||||||
| Source of variation | ||||||||||||||
| ANOVA P values | ||||||||||||||
| Variety | <0.0001 | <0.0001 | 0.1254 | 0.2156 | <0.0001 | 0.3201 | 0.2499 | 0.0626 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0498 | <0.0001 |
| Seed priming | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Variety × seed priming | <0.0001 | <0.0001 | <0.0001 | 0.0015 | 0.0002 | 0.0719 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0025 | 0.0335 |
| Treatment | AsA (Root) | AsA (Shoot) | GR/GSH (Root) | GR/GSH (Shoot) |
|---|---|---|---|---|
| Main Effects | ||||
| Variety | ||||
| Gomti | 23.5b | 33.1a | 82.6a | 46.1b |
| Kalijeera | 28.3a | 33.4a | 78.0a | 52.7a |
| Seed priming | ||||
| Control | 36.1a | 48.8a | 110.0a | 90.6a |
| As | 14.4d | 18.7d | 43.9d | 19.5d |
| As + Zn (0.5 mg L−1) | 23.2c | 29.3c | 73.5c | 37.0c |
| As + An (1.0 mg L−1) | 29.9b | 36.2b | 93.7b | 50.5b |
| Interaction Effects | ||||
| Gomti × Control | 31.8b | 51.2a | 111.0a | 82.0a |
| Gomti × As | 12.3e | 18.3d | 32.3c | 19.9d |
| Gomti × As × Zn (0.5 mg L−1) | 21.2cd | 28.4cd | 80.7ab | 28.4cd |
| Gomti × As × Zn (1.0 mg L−1) | 28.9bc | 34.4c | 106.1a | 54.3b |
| Kalijeera × Control | 40.5a | 46.4ab | 109.0a | 99.3a |
| Kalijeera × As | 16.5de | 19.1d | 55.5bc | 19.1d |
| Kalijeera × As × Zn (0.5 mg L−1) | 25.3bc | 30.2c | 66.2b | 45.6bc |
| Kalijeera × As × Zn (1.0 mg L−1) | 30.9b | 37.9bc | 81.3ab | 46.8bc |
| ANOVA | ||||
| Sources of Variations | ||||
| ANOVA p-value | ||||
| Variety | 0.0007 | 0.8447 | 0.3557 | 0.0397 |
| Seed priming | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Variety × seed priming | 0.3103 | 0.3570 | 0.0098 | 0.0118 |
| Receptor | Ligands | MolDock Score | Rerank Score | HBond |
|---|---|---|---|---|
| Superoxide dismutase (Mn-SOD) | Superoxide radical | −18.6347 | −16.1787 | −3.10127 |
| As3+ | −19.8086 | −20.6329 | 0 | |
| Zn2+ | −19.8081 | −20.6325 | 0 | |
| AsA | −80.6404 | −71.5325 | −9.56283 | |
| GSH | −114.497 | −98.2422 | −10.4638 | |
| Catalase (CAT) | H2O2 | −18.8881 | −16.2571 | −5.68477 |
| As3+ | −21.7768 | −21.3154 | 0 | |
| Zn2+ | −20.6794 | −20.1665 | 0 | |
| AsA | −65.3445 | −61.1279 | −10.3087 | |
| GSH | −85.3666 | −73.7135 | −7.49339 | |
| Glutathione reductase (GR) | As | −27.7162 | −19.5761 | 0 |
| Zn | −28.2799 | −24.9889 | 0 | |
| AsA | −79.9338 | −71.3906 | −9.22549 | |
| GSH | −116.038 | −100.133 | −11.5623 | |
| Glutathione peroxidase (GPx) | H2O2 | −20.0855 | −17.0414 | −5.25808 |
| As3+ | −22.9651 | −24.3977 | 0 | |
| Zn2+ | −22.9649 | −24.3974 | 0 | |
| AsA | −74.8541 | −68.4545 | −9.85995 | |
| GSH | −106.223 | −92.9154 | −14.2971 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choudhury, S.; Moulick, D.; Mazumder, M.K.; Pattnaik, B.K.; Ghosh, D.; Vemireddy, L.R.; Aldhahrani, A.; Soliman, M.M.; Gaber, A.; Hossain, A. An In Vitro and In Silico Perspective Study of Seed Priming with Zinc on the Phytotoxicity and Accumulation Pattern of Arsenic in Rice Seedlings. Antioxidants 2022, 11, 1500. https://doi.org/10.3390/antiox11081500
Choudhury S, Moulick D, Mazumder MK, Pattnaik BK, Ghosh D, Vemireddy LR, Aldhahrani A, Soliman MM, Gaber A, Hossain A. An In Vitro and In Silico Perspective Study of Seed Priming with Zinc on the Phytotoxicity and Accumulation Pattern of Arsenic in Rice Seedlings. Antioxidants. 2022; 11(8):1500. https://doi.org/10.3390/antiox11081500
Chicago/Turabian StyleChoudhury, Shuvasish, Debojyoti Moulick, Muhammed Khairujjaman Mazumder, Binaya Kumar Pattnaik, Dibakar Ghosh, Lakshminarayana R. Vemireddy, Adil Aldhahrani, Mohamed Mohamed Soliman, Ahmed Gaber, and Akbar Hossain. 2022. "An In Vitro and In Silico Perspective Study of Seed Priming with Zinc on the Phytotoxicity and Accumulation Pattern of Arsenic in Rice Seedlings" Antioxidants 11, no. 8: 1500. https://doi.org/10.3390/antiox11081500
APA StyleChoudhury, S., Moulick, D., Mazumder, M. K., Pattnaik, B. K., Ghosh, D., Vemireddy, L. R., Aldhahrani, A., Soliman, M. M., Gaber, A., & Hossain, A. (2022). An In Vitro and In Silico Perspective Study of Seed Priming with Zinc on the Phytotoxicity and Accumulation Pattern of Arsenic in Rice Seedlings. Antioxidants, 11(8), 1500. https://doi.org/10.3390/antiox11081500