
Leonurine Attenuates Obesity-Related Vascular Dysfunction and Inflammation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
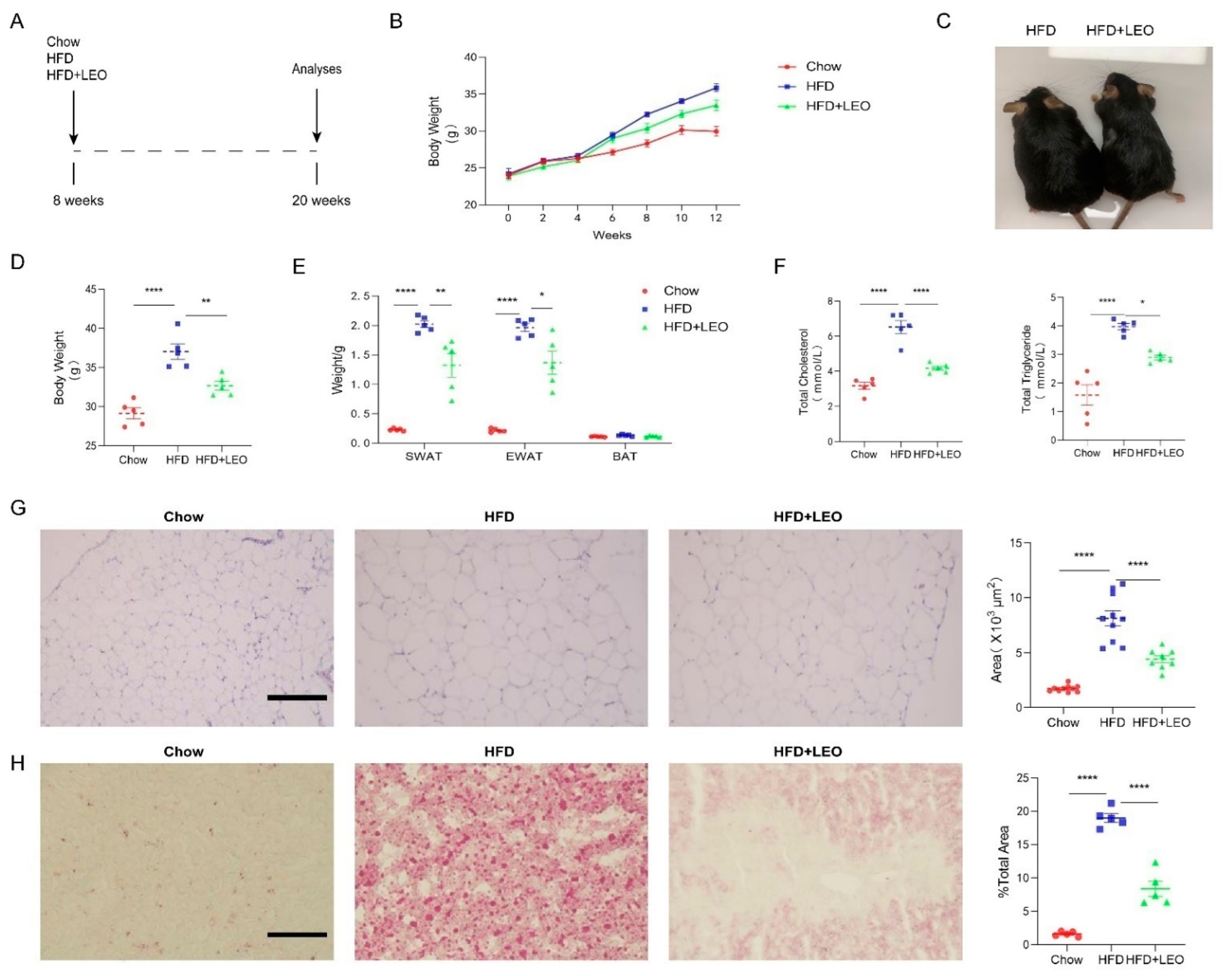
2.2. Animal and Diets
2.3. Anatomic Assessment and Biochemical Analyses
2.4. Quantitative Real-Time PCR
2.5. Haematoxylin and Eosin Staining
2.6. Oil Red O Staining
2.7. Immunofluorescence
2.8. Masson’s Trichrome Staining
2.9. In Vitro Mouse Mesenteric Artery Ring Experiment
2.10. Cell Culture
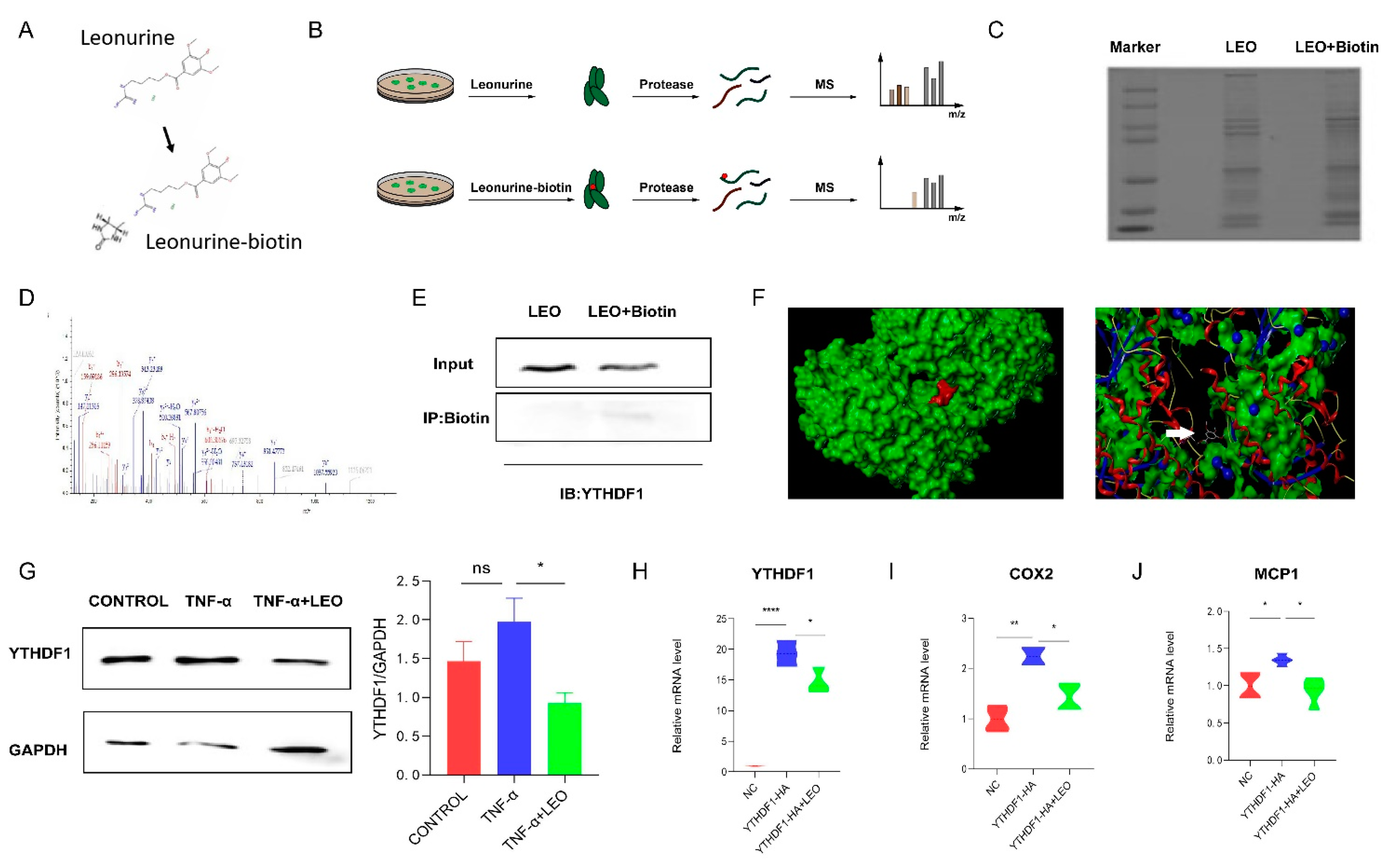
2.11. Pull-Down Assay and Western Blot
2.12. Statistical Analyses
3. Results
3.1. LEO Alleviates Obesity and Lipid Deposition Induced by HFD
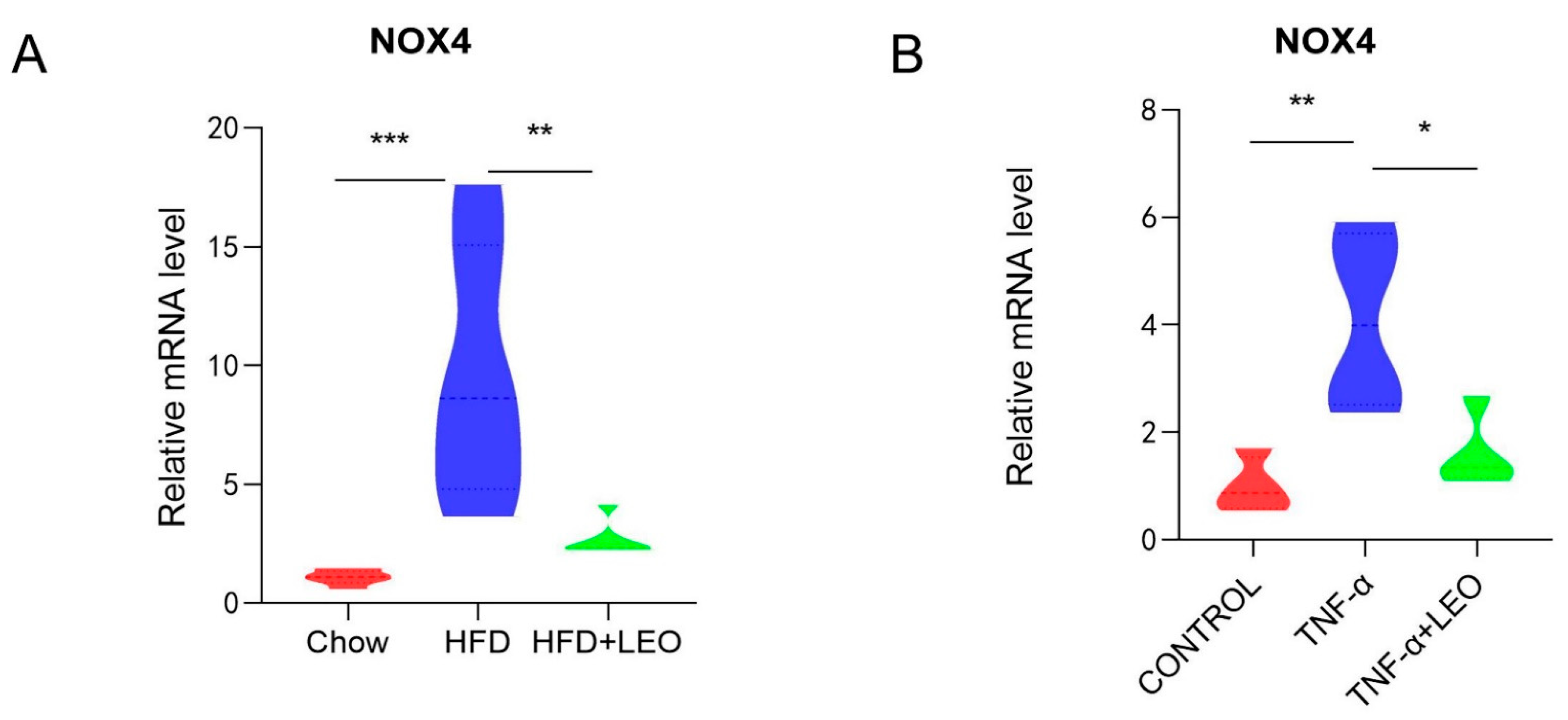
3.2. LEO Improves Inflammation In Vivo and In Vitro
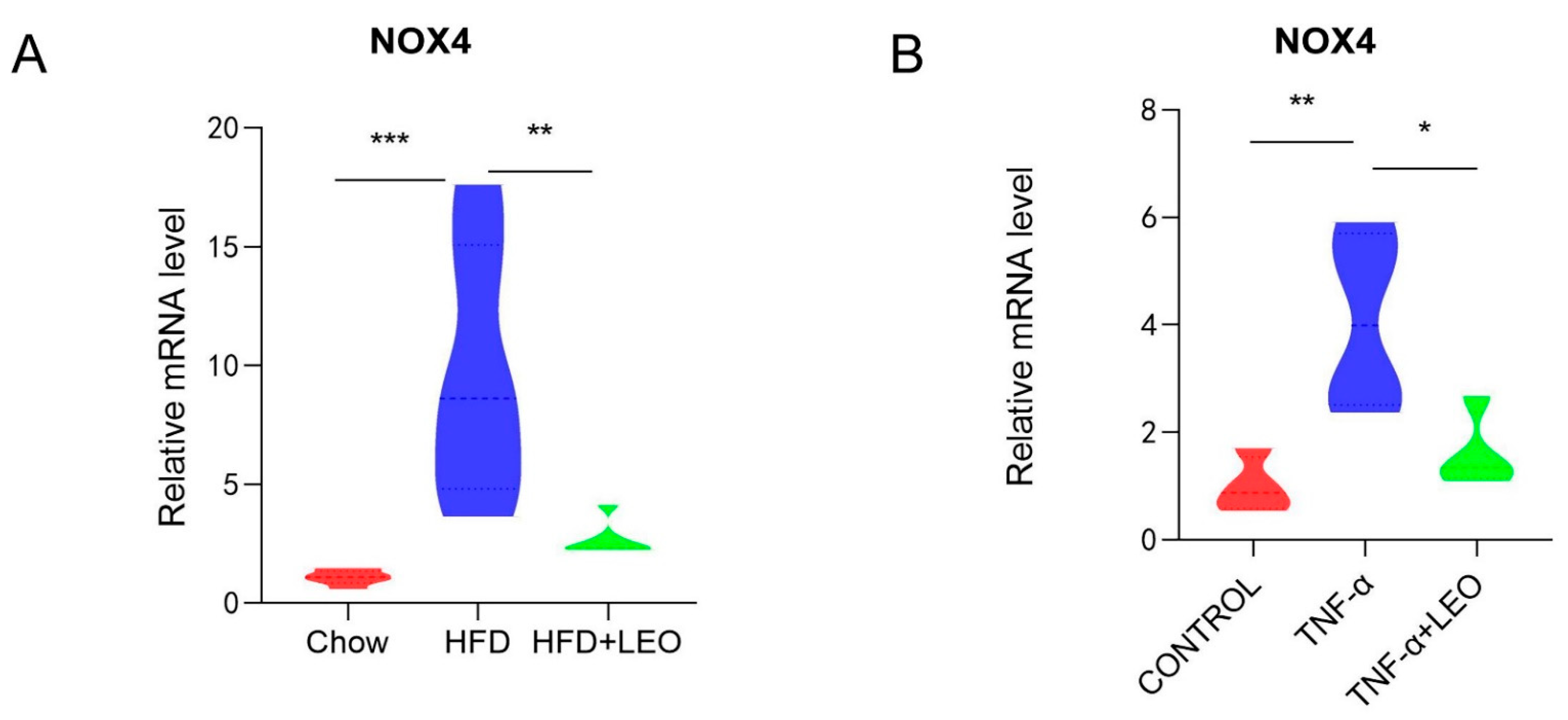
3.3. LEO Alleviates Oxidative Stress In Vivo and In Vitro
3.4. LEO Ameliorates Mesenteric Artery Dysfunction Induced by Obesity
3.5. LEO Binds YTHDF1 Protein
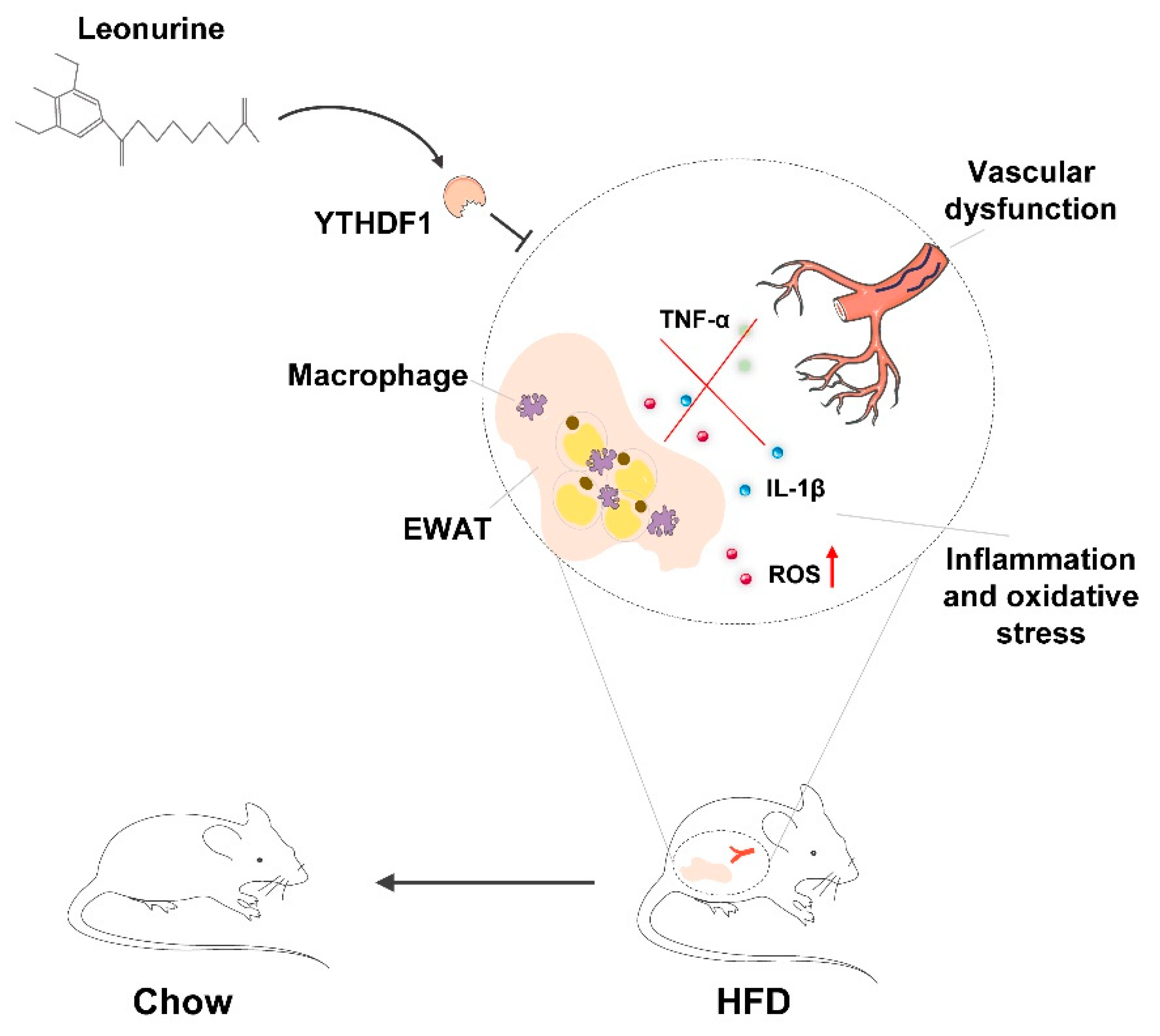
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fitzgerald, F.T. The problem of obesity. Annu. Rev. Med. 1981, 32, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Gadde, K.M.; Martin, C.K.; Berthoud, H.R.; Heymsfield, S.B. Obesity: Pathophysiology and Management. J. Am. Coll. Cardiol. 2018, 71, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Funcke, J.B.; Scherer, P.E. Beyond adiponectin and leptin: Adipose tissue-derived mediators of inter-organ communication. J. Lipid Res. 2019, 60, 1648–1684. [Google Scholar] [CrossRef]
- Zhao, S.; Kusminski, C.M.; Scherer, P.E. Adiponectin, Leptin and Cardiovascular Disorders. Circ. Res. 2021, 128, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Van Gaal, L.F.; Mertens, I.L.; De Block, C.E. Mechanisms linking obesity with cardiovascular disease. Nature 2006, 444, 875–880. [Google Scholar] [CrossRef]
- Karam, B.S.; Chavez-Moreno, A.; Koh, W.; Akar, J.G.; Akar, F.G. Oxidative stress and inflammation as central mediators of atrial fibrillation in obesity and diabetes. Cardiovasc. Diabetol. 2017, 16, 120. [Google Scholar] [CrossRef]
- Man, A.W.C.; Xia, N.; Li, H. Circadian Rhythm in Adipose Tissue: Novel Antioxidant Target for Metabolic and Cardiovascular Diseases. Antioxidants 2020, 9, 968. [Google Scholar] [CrossRef]
- Muñoz, M.; López-Oliva, M.E.; Rodríguez, C.; Martínez, M.P.; Sáenz-Medina, J.; Sánchez, A.; Climent, B.; Benedito, S.; García-Sacristán, A.; Rivera, L.; et al. Differential contribution of Nox1, Nox2 and Nox4 to kidney vascular oxidative stress and endothelial dysfunction in obesity. Redox Biol. 2020, 28, 101330. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Blank, A.; Cui, C.; Schoenfelt, K.Q.; Zhou, G.; Xu, Y.; Khramtsova, G.; Olopade, F.; Shah, A.M.; Khan, S.A.; et al. Metabolically activated adipose tissue macrophages link obesity to triple-negative breast cancer. J. Exp. Med. 2019, 216, 1345–1358. [Google Scholar] [CrossRef]
- Engin, A. The Pathogenesis of Obesity-Associated Adipose Tissue Inflammation. Adv. Exp. Med. Biol. 2017, 960, 221–245. [Google Scholar] [PubMed]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am. J. Physiol. Cell Physiol. 2021, 320, C375–C391. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Pan, H.; Wang, X.; He, H.; Li, M. Leonurus japonicus Houtt.: Ethnopharmacology, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 2014, 152, 14–32. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Lin, Y.K.; Liu, X.H.; Wang, L.; Yu, M.; Li, D.J.; Zhu, Y.Z.; Du, M.R. Leonurine: From Gynecologic Medicine to Pleiotropic Agent. Chin. J. Integr. Med. 2020, 26, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Xu, D.Q.; Chen, Y.Y.; Yue, S.J.; Tang, Y.P. Leonurine, a potential drug for the treatment of cardiovascular system and central nervous system diseases. Brain Behav. 2021, 11, e01995. [Google Scholar] [CrossRef]
- Song, X.; Wang, T.; Zhang, Z.; Jiang, H.; Wang, W.; Cao, Y.; Zhang, N. Leonurine exerts anti-inflammatory effect by regulating inflammatory signaling pathways and cytokines in LPS-induced mouse mastitis. Inflammation 2015, 38, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Gong, L.; Zhou, M.; Xue, X.; Li, Y.; Peng, C. Leonurine Ameliorates Oxidative Stress and Insufficient Angiogenesis by Regulating the PI3K/Akt-eNOS Signaling Pathway in H2O2-Induced HUVECs. Oxidative Med. Cell. Longev. 2021, 2021, 9919466. [Google Scholar] [CrossRef]
- Xu, D.; Chen, M.; Ren, X.; Ren, X.; Wu, Y. Leonurine ameliorates LPS-induced acute kidney injury via suppressing ROS-mediated NF-κB signaling pathway. Fitoterapia 2014, 97, 148–155. [Google Scholar] [CrossRef]
- Piera-Velazquez, S.; Jimenez, S.A. Oxidative Stress Induced by Reactive Oxygen Species (ROS) and NADPH Oxidase 4 (NOX4) in the Pathogenesis of the Fibrotic Process in Systemic Sclerosis: A Promising Therapeutic Target. J. Clin. Med. 2021, 10, 4791. [Google Scholar] [CrossRef]
- Olesen, N.D.; Fischer, M.; Secher, N.H. Sodium nitroprusside dilates cerebral vessels and enhances internal carotid artery flow in young men. J. Physiol. 2018, 596, 3967–3976. [Google Scholar] [CrossRef]
- Buzinari, T.C.; Oishi, J.C.; De Moraes, T.F.; Vatanabe, I.P.; Selistre-de-Araújo, H.S.; Pestana, C.R.; Rodrigues, G.J. Treatment with sodium nitroprusside improves the endothelial function in aortic rings with endothelial dysfunction. Eur. J. Pharm. Sci. 2017, 105, 144–149. [Google Scholar] [CrossRef]
- Liu, X.; Wang, X.; Pan, Y.; Zhao, L.; Sun, S.; Luo, A.; Bao, C.; Tang, H.; Han, Y. Artemisinin Improves Acetylcholine-Induced Vasodilatation in Rats with Primary Hypertension. Drug Des. Dev. Ther. 2021, 15, 4489–4502. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Xiao, X.; Jie, F.; Cheng, Y.; Jin, M.; Yin, Y.; Wang, Y. YTHDF1 promotes NLRP3 translation to induce intestinal epithelial cell inflammatory injury during endotoxic shock. Sci. China Life Sci. 2021, 64, 1988–1991. [Google Scholar] [CrossRef]
- Li, J.; Wu, H.; Liu, Y.; Yang, L. High fat diet induced obesity model using four strainsof mice: Kunming, C57BL/6, BALB/c and ICR. Exp. Anim. 2020, 69, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Guo, C.C.; Yu, Y.X.; Xie, L.; Chang, C.Q. Establishment of high-fat diet-induced obesity and insulin resistance model in rats. Beijing Da Xue Xue Bao Yi Xue Ban 2020, 52, 557–563. [Google Scholar]
- Goossens, G.H. The Metabolic Phenotype in Obesity: Fat Mass, Body Fat Distribution, and Adipose Tissue Function. Obes. Facts 2017, 10, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef]
- Ning, K.; Wang, M.J.; Lin, G.; Zhang, Y.L.; Li, M.Y.; Yang, B.F.; Chen, Y.; Huang, Y.; Li, Z.M.; Huang, Y.J.; et al. eNOS-Nitric Oxide System Contributes to a Novel Antiatherogenic Effect of Leonurine via Inflammation Inhibition and Plaque Stabilization. J. Pharmacol. Exp. Ther. 2020, 373, 463–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Heijden, R.A.; Sheedfar, F.; Morrison, M.C.; Hommelberg, P.P.; Kor, D.; Kloosterhuis, N.J.; Gruben, N.; Youssef, S.A.; de Bruin, A.; Hofker, M.H.; et al. High-fat diet induced obesity primes inflammation in adipose tissue prior to liver in C57BL/6j mice. Aging 2015, 7, 256–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piché, M.E.; Tchernof, A.; Després, J.P. Obesity Phenotypes, Diabetes, and Cardiovascular Diseases. Circ. Res. 2020, 126, 1477–1500. [Google Scholar] [CrossRef]
- Vecchié, A.; Dallegri, F.; Carbone, F.; Bonaventura, A.; Liberale, L.; Portincasa, P.; Frühbeck, G.; Montecucco, F. Obesity phenotypes and their paradoxical association with cardiovascular diseases. Eur. J. Intern. Med. 2018, 48, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Danielak, A.; Wojcik, D.; Mazur-Bialy, A.; Surmiak, M.; Bilski, J.; Targosz, A.; Magierowski, M.; Chmura, A.; Strzalka, M.; Krzysiek-Maczka, G.; et al. Intestinal Alkaline Phosphatase Combined with Voluntary Physical Activity Alleviates Experimental Colitis in Obese Mice. Involvement of Oxidative Stress, Myokines, Adipokines and Proinflammatory Biomarkers. Antioxidants 2021, 10, 240. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.Z.; Wu, W.; Zhu, Q.; Liu, X. Discovery of Leonuri and therapeutical applications: From bench to bedside. Pharmacol. Ther. 2018, 188, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.Y.; Shi, X.R.; Zhu, K.; Wu, T.T.; Zhu, Y.Z. SCM-198 inhibits microglial overactivation and attenuates Aβ(1–40)-induced cognitive impairments in rats via JNK and NF-κB pathways. J. Neuroinflamm. 2014, 11, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Li, Y.; Li, X.; Liu, X.; Wang, X.; Yu, M.; Zhu, Y.; Du, M. SCM-198 ameliorates endometrial inflammation via suppressing the LPS-JNK-cJUN/cFOS-TLR4-NF-κB pathway. Acta Biochim. Biophys. Sin. 2021, 53, 1207–1215. [Google Scholar] [CrossRef]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [Green Version]
- Čolak, E.; Pap, D. The role of oxidative stress in the development of obesity and obesity-related metabolic disorders. J. Med. Biochem. 2021, 40, 1–9. [Google Scholar] [CrossRef]
- García, J.G.; de Miguel, C.; Milagro, F.I.; Zalba, G.; Ansorena, E. Endothelial NOX5 Expression Modulates Thermogenesis and Lipolysis in Mice Fed with a High-Fat Diet and 3T3-L1 Adipocytes through an Interleukin-6 Dependent Mechanism. Antioxidants 2021, 11, 30. [Google Scholar] [CrossRef]
- Pérez-Torres, I.; Castrejón-Téllez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative Stress, Plant Natural Antioxidants, and Obesity. Int. J. Mol. Sci. 2021, 22, 1786. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Lei, X.; Chai, N.; Linghu, E. NOX4: A potential therapeutic target for pancreatic cancer and its mechanism. J. Transl. Med. 2021, 19, 515. [Google Scholar] [CrossRef]
- Sharma, A.; Singh, S.; Ahmad, S.; Gulzar, F.; Schertzer, J.D.; Tamrakar, A.K. NOD1 activation induces oxidative stress via NOX1/4 in adipocytes. Free Radic. Biol. Med. 2021, 162, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Guzik, T.J.; Touyz, R.M. Oxidative Stress, Inflammation, and Vascular Aging in Hypertension. Hypertension 2017, 70, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Byzova, T.V. Oxidative stress in angiogenesis and vascular disease. Blood 2014, 123, 625–631. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.W.; Shi, H.; Winkler, M.A.; Lee, R.; Weintraub, N.L. Perivascular Adipose Tissue and Vascular Perturbation/Atherosclerosis. Arter. Thromb. Vasc. Biol. 2020, 40, 2569–2576. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Reifenberg, G.; Schirra, C.; Li, H. The Involvement of Sirtuin 1 Dysfunction in High-Fat Diet-Induced Vascular Dysfunction in Mice. Antioxidants 2022, 11, 541. [Google Scholar] [CrossRef]
- Qi, J.; Wang, J.J.; Duan, J.L.; Lu, Z.Y.; Yuan, Y.G. Leonurine Improves Age-Dependent Impaired Angiogenesis: Possible Involvement of Mitochondrial Function and HIF-1α Dependent VEGF Activation. Front. Pharmacol. 2017, 8, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, C.; Lee, M.D.; McCarron, J.G. Acetylcholine released by endothelial cells facilitates flow-mediated dilatation. J. Physiol. 2016, 594, 7267–7307. [Google Scholar] [CrossRef] [PubMed]
- Zoupa, E.; Pitsikas, N. The Nitric Oxide (NO) Donor Sodium Nitroprusside (SNP) and Its Potential for the Schizophrenia Therapy: Lights and Shadows. Molecules 2021, 26, 3196. [Google Scholar] [CrossRef]
- Touyz, R.M. Reactive oxygen species and angiotensin II signaling in vascular cells: Implications in cardiovascular disease. Braz. J. Med. Biol. Res. 2004, 37, 1263–1273. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence (5′–3′) |
|---|---|
| GAPDH-F | CTAAAGGGCATCCTGGGC |
| GAPDH-R | TTACTCCTTGGAGGCCAT |
| MCP-1-F | CAGGTCCCTGTCATGCTTCT |
| MCP-1-R | GTGGGGCGTTAACTGCATCT |
| IL-1β-F | GCAACTGTTCCTGAACTCAACT |
| IL-1β-R | ATCTTTTGGGGTCCGTCAACT |
| TNF-α-F | CCCTCACACTCAGATCATCTTCT |
| TNF-α-R | GCTACGACGTGGGCTACAG |
| NOX4-F | CCAAATGTTGGGCGATTGTGT |
| NOX4-R | ATCCATACTCTGCTGTGCCA |
| YTHDF1-F | CTGCAGTTAAGACGGTGGGT |
| YTHDF1-R | TAGCAATGGCTGCCCATGAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, X.-D.; Zhang, J.-X.; Hu, X.-D.; Zhuang, T.; Lu, N.; Ruan, C.-C. Leonurine Attenuates Obesity-Related Vascular Dysfunction and Inflammation. Antioxidants 2022, 11, 1338. https://doi.org/10.3390/antiox11071338
Shi X-D, Zhang J-X, Hu X-D, Zhuang T, Lu N, Ruan C-C. Leonurine Attenuates Obesity-Related Vascular Dysfunction and Inflammation. Antioxidants. 2022; 11(7):1338. https://doi.org/10.3390/antiox11071338
Chicago/Turabian StyleShi, Xiao-Dong, Jia-Xin Zhang, Xi-De Hu, Tao Zhuang, Ning Lu, and Cheng-Chao Ruan. 2022. "Leonurine Attenuates Obesity-Related Vascular Dysfunction and Inflammation" Antioxidants 11, no. 7: 1338. https://doi.org/10.3390/antiox11071338
APA StyleShi, X.-D., Zhang, J.-X., Hu, X.-D., Zhuang, T., Lu, N., & Ruan, C.-C. (2022). Leonurine Attenuates Obesity-Related Vascular Dysfunction and Inflammation. Antioxidants, 11(7), 1338. https://doi.org/10.3390/antiox11071338