
Increase of HO-1 Expression in Critically Ill COVID-19 Patients Is Associated with Poor Prognosis and Outcome

 , , ,
, , ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Histopathology and Immunohistochemistry
2.3. RNA Extraction, Reverse Transcription and Real-Time PCR
2.4. Statistical Analysis
3. Results
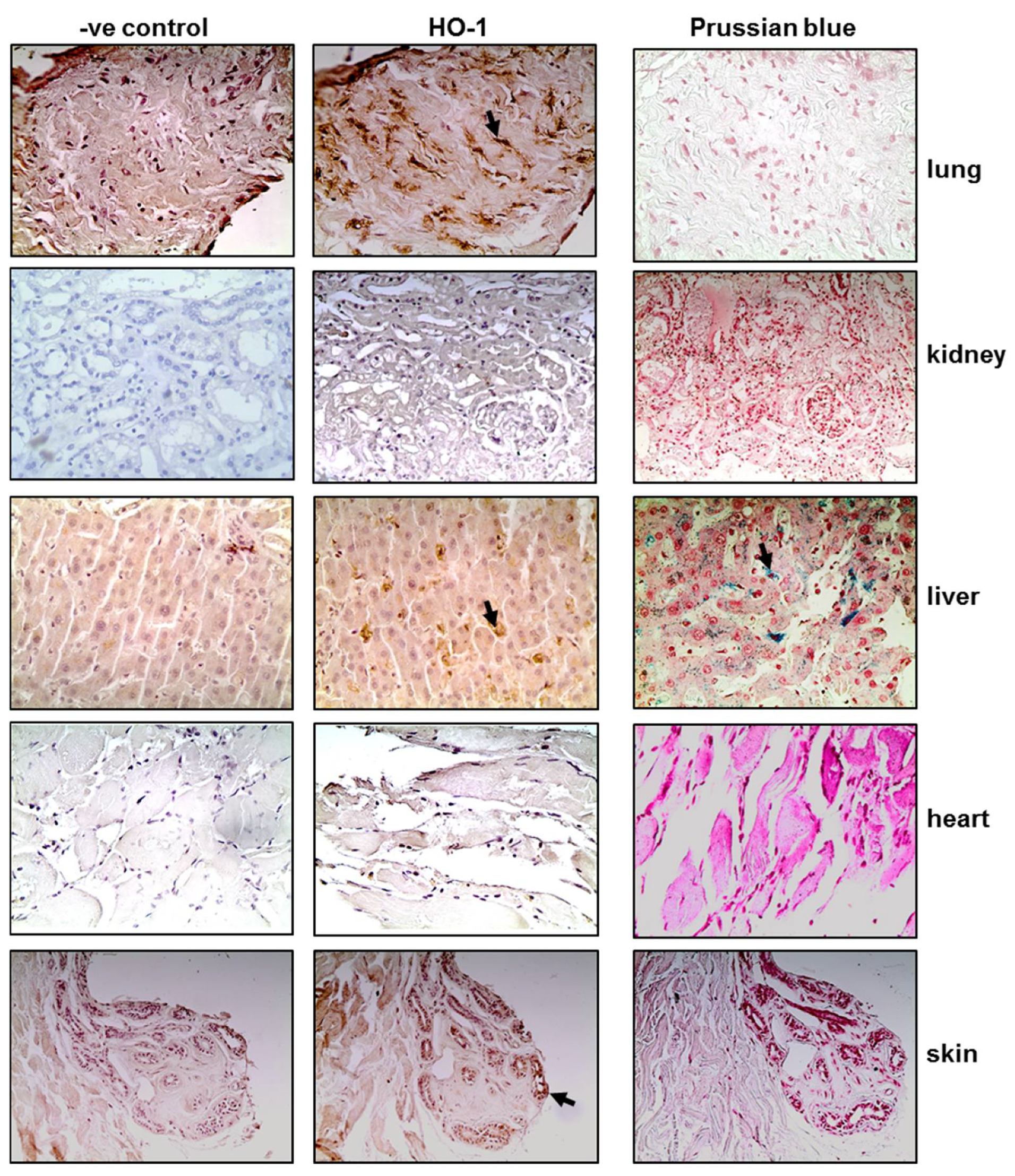
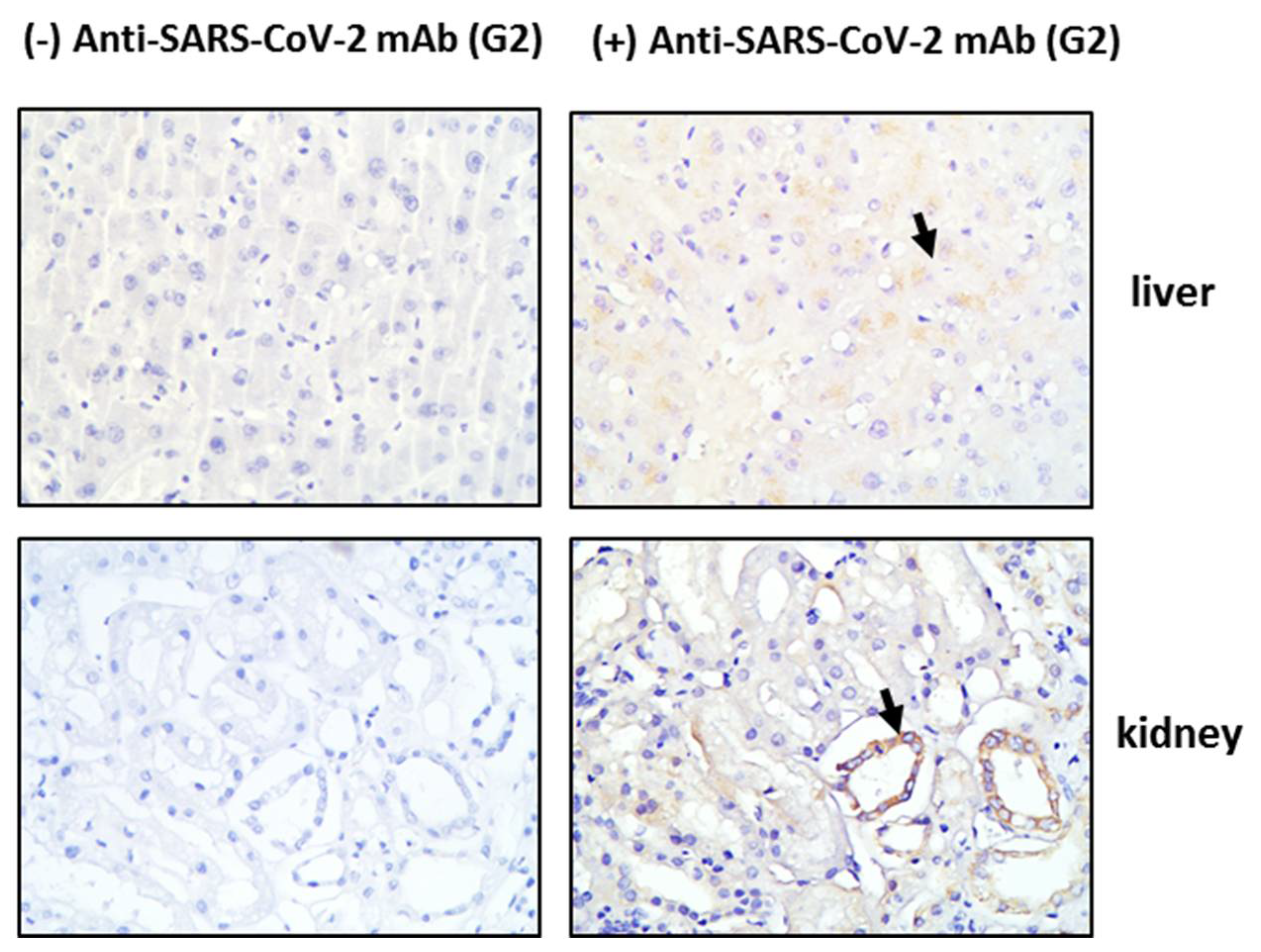
3.1. Tissue HO-1 Induction Is Independent from SARS-CoV-2 Presence
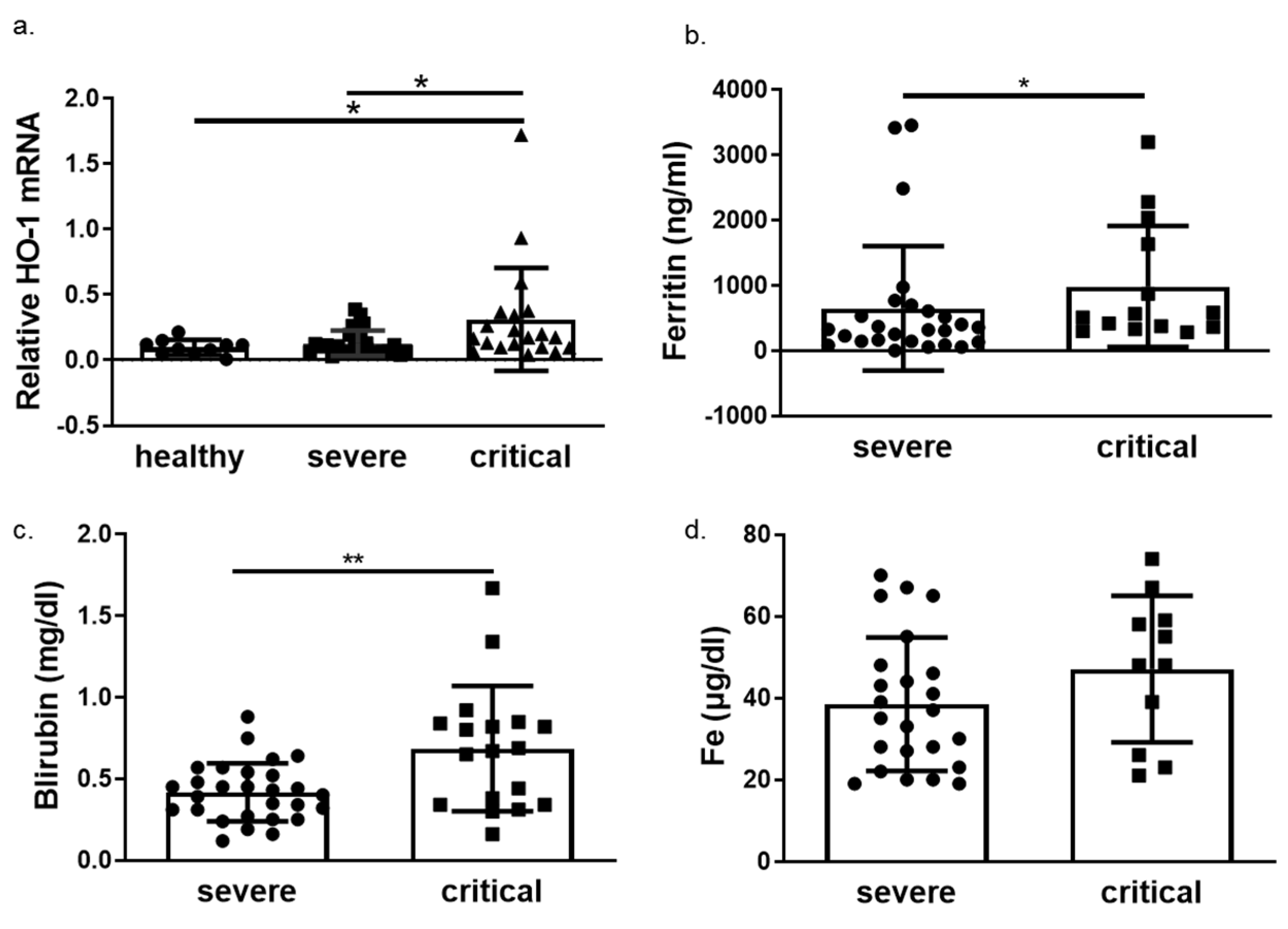
3.2. HO-1 Induction in Blood Samples of COVID-19 Patients
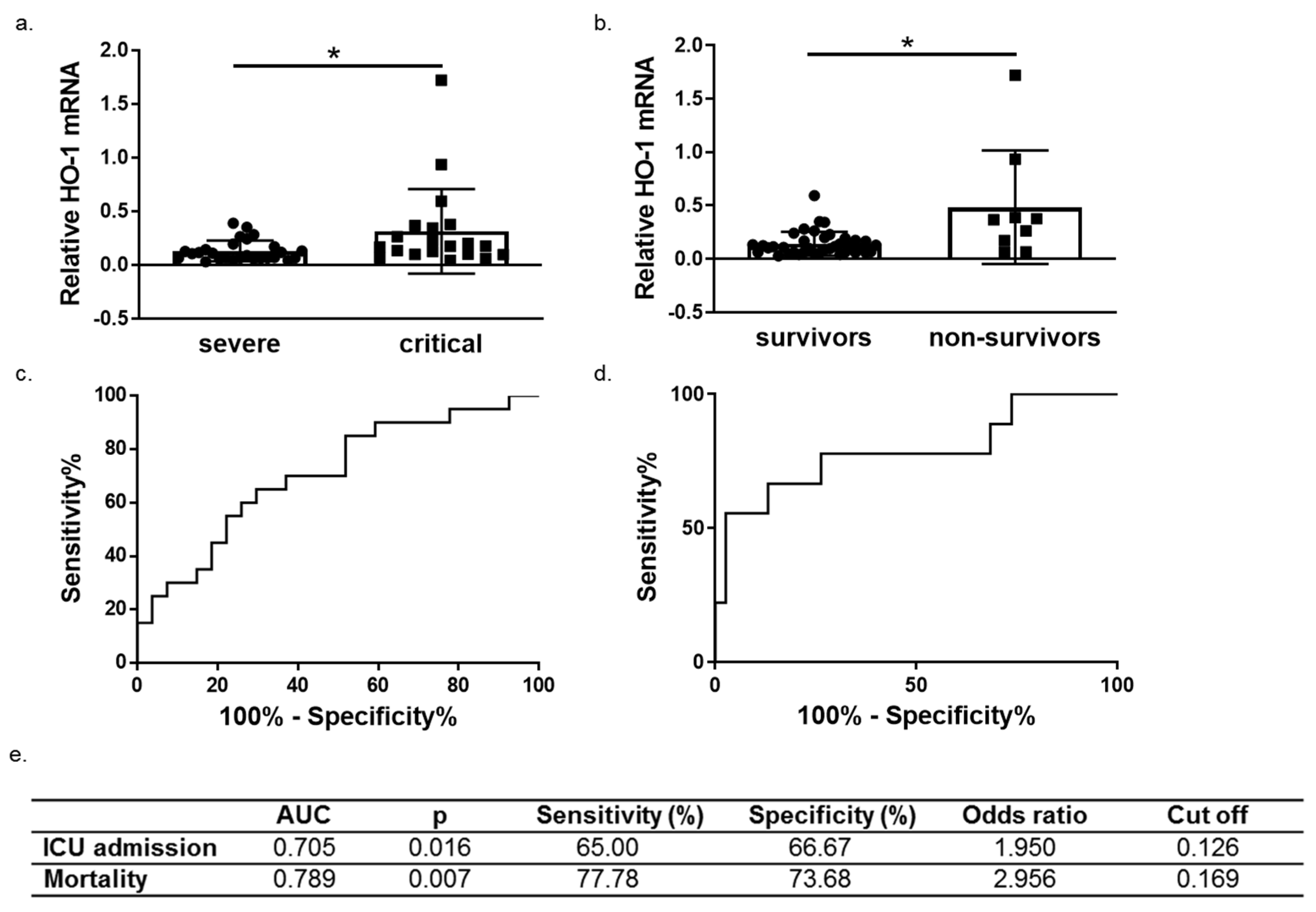
3.3. HO-1 Induction in COVID-19 Patients Is Associated with Disease Progression and Mortality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maines, M.D. The heme oxygenase system: A regulator of second messenger gases. Annu. Rev. Pharmacol. Toxicol. 1997, 37, 517–554. [Google Scholar] [CrossRef] [PubMed]
- Grochot-Przeczek, A.; Dulak, J.; Jozkowicz, A. Haem oxygenase-1: Non-canonical roles in physiology and pathology. Clin. Sci. 2012, 122, 93–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poss, K.D.; Tonegawa, S. Heme oxygenase 1 is required for mammalian iron reutilization. Proc. Natl. Acad. Sci. USA 1997, 94, 10919–10924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinoza, J.A.; Gonzalez, P.A.; Kalergis, A.M. Modulation of Antiviral Immunity by Heme Oxygenase-1. Am. J. Pathol. 2017, 187, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Wiesel, P.; Patel, A.P.; DiFonzo, N.; Marria, P.B.; Sim, C.U.; Pellacani, A.; Maemura, K.; LeBlanc, B.W.; Marino, K.; Doerschuk, C.M.; et al. Endotoxin-induced mortality is related to increased oxidative stress and end-organ dysfunction, not refractory hypotension, in heme oxygenase-1-deficient mice. Circulation 2000, 102, 3015–3022. [Google Scholar] [CrossRef] [Green Version]
- Saukkonen, K.; Lakkisto, P.; Kaunisto, M.A.; Varpula, M.; Voipio-Pulkki, L.M.; Varpula, T.; Pettila, V.; Pulkki, K. Heme oxygenase 1 polymorphisms and plasma concentrations in critically ill patients. Shock 2010, 34, 558–564. [Google Scholar] [CrossRef]
- Tung, W.H.; Hsieh, H.L.; Lee, I.T.; Yang, C.M. Enterovirus 71 induces integrin beta1/EGFR-Rac1-dependent oxidative stress in SK-N-SH cells: Role of HO-1/CO in viral replication. J. Cell. Physiol. 2011, 226, 3316–3329. [Google Scholar] [CrossRef]
- Ma, L.L.; Zhang, P.; Wang, H.Q.; Li, Y.F.; Hu, J.; Jiang, J.D.; Li, Y.H. Heme oxygenase-1 agonist CoPP suppresses influenza virus replication through IRF3-mediated generation of IFN-alpha/beta. Virology 2019, 528, 80–88. [Google Scholar] [CrossRef]
- Wu, Y.H.; Chen, W.C.; Tseng, C.K.; Chen, Y.H.; Lin, C.K.; Lee, J.C. Heme oxygenase-1 inhibits DENV-induced endothelial hyperpermeability and serves as a potential target against dengue hemorrhagic fever. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2022, 36, e22110. [Google Scholar] [CrossRef]
- Jablonowska, E.; Wojcik, K.; Szymanska, B.; Omulecka, A.; Cwiklinska, H.; Piekarska, A. Hepatic HMOX1 expression positively correlates with Bach-1 and miR-122 in patients with HCV mono and HIV/HCV coinfection. PLoS ONE 2014, 9, e95564. [Google Scholar] [CrossRef]
- Wagener, F.; Pickkers, P.; Peterson, S.J.; Immenschuh, S.; Abraham, N.G. Targeting the Heme-Heme Oxygenase System to Prevent Severe Complications Following COVID-19 Infections. Antioxidants 2020, 9, 540. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Piagnerelli, M.; Van Meerhaeghe, A.; Zouaoui Boudjeltia, K. Heme oxygenase-1 (HO-1) cytoprotective pathway: A potential treatment strategy against coronavirus disease 2019 (COVID-19)-induced cytokine storm syndrome. Med. Hypotheses 2020, 144, 110242. [Google Scholar] [CrossRef] [PubMed]
- Evangelou, K.; Veroutis, D.; Paschalaki, K.; Foukas, P.G.; Lagopati, N.; Dimitriou, M.; Papaspyropoulos, A.; Konda, B.; Hazapis, O.; Polyzou, A.; et al. Pulmonary infection by SARS-CoV-2 induces senescence accompanied by an inflammatory phenotype in severe COVID-19: Possible implications for viral mutagenesis. Eur. Respir. J. 2022. [Google Scholar] [CrossRef] [PubMed]
- Chok, M.K.; Ferlicot, S.; Conti, M.; Almolki, A.; Durrbach, A.; Loric, S.; Benoit, G.; Droupy, S.; Eschwege, P. Renoprotective potency of heme oxygenase-1 induction in rat renal ischemia-reperfusion. Inflamm. Allergy Drug Targets 2009, 8, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Delbauve, S.; Roumeguere, T.; Wespes, E.; Leo, O.; Flamand, V.; Le Moine, A.; Hougardy, J.M. HO-1 mitigates acute kidney injury and subsequent kidney-lung cross-talk. Free. Radic. Res. 2019, 53, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- El Kalamouni, C.; Frumence, E.; Bos, S.; Turpin, J.; Nativel, B.; Harrabi, W.; Wilkinson, D.A.; Meilhac, O.; Gadea, G.; Despres, P.; et al. Subversion of the Heme Oxygenase-1 Antiviral Activity by Zika Virus. Viruses 2018, 11, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill-Batorski, L.; Halfmann, P.; Neumann, G.; Kawaoka, Y. The cytoprotective enzyme heme oxygenase-1 suppresses Ebola virus replication. J. Virol. 2013, 87, 13795–13802. [Google Scholar] [CrossRef] [Green Version]
- Devadas, K.; Dhawan, S. Hemin activation ameliorates HIV-1 infection via heme oxygenase-1 induction. J. Immunol. 2006, 176, 4252–4257. [Google Scholar] [CrossRef]
- Maestro, S.; Cordoba, K.M.; Olague, C.; Argemi, J.; Avila, M.A.; Gonzalez-Aseguinolaza, G.; Smerdou, C.; Fontanellas, A. Heme oxygenase-1 inducer hemin does not inhibit SARS-CoV-2 virus infection. Biomed. Pharmacother. 2021, 137, 111384. [Google Scholar] [CrossRef]
- Cazzato, G.; Colagrande, A.; Cimmino, A.; Cicco, G.; Scarcella, V.S.; Tarantino, P.; Lospalluti, L.; Romita, P.; Foti, C.; Demarco, A.; et al. HMGB1-TIM3-HO1: A New Pathway of Inflammation in Skin of SARS-CoV-2 Patients? A Retrospective Pilot Study. Biomolecules 2021, 11, 1219. [Google Scholar] [CrossRef]
- Suttner, D.M.; Dennery, P.A. Reversal of HO-1 related cytoprotection with increased expression is due to reactive iron. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1999, 13, 1800–1809. [Google Scholar] [CrossRef] [PubMed]
- Kvam, E.; Hejmadi, V.; Ryter, S.; Pourzand, C.; Tyrrell, R.M. Heme oxygenase activity causes transient hypersensitivity to oxidative ultraviolet a radiation that depends on release of iron from heme. Free. Radic. Biol. Med. 2000, 28, 1191–1196. [Google Scholar] [CrossRef]
- Su, W.L.; Lin, C.P.; Hang, H.C.; Wu, P.S.; Cheng, C.F.; Chao, Y.C. Desaturation and heme elevation during COVID-19 infection: A potential prognostic factor of heme oxygenase-1. J. Microbiol. Immunol. Infect. 2021, 54, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y.; Oshima, Y.; Tagami, Y.; Aoki, A.; Fujii, H.; Izawa, A.; Seki, K.; Kanai, A.; Yabe, A.; Watanabe, K.; et al. Clinical importance of serum heme oxygenase-1 measurement in patients with acute exacerbation of idiopathic pulmonary fibrosis triggered by coronavirus disease 2019. Respir. Med. Case Rep. 2022, 36, 101615. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Severe COVID-19 | Critical COVID-19 | p Value | |
|---|---|---|---|
| Age (years) | 60.26 ± 15.96 | 66.65 ± 11.45 | 0.149 |
| Male | 12 (44.44%) | 15 (75.00%) | 0.111 |
| Comorbidities | 19 (70.37%) | 15 (75.00%) | 0.940 |
| Symptoms | |||
| paO2/FIO2 | 318.80 ± 63.62 | 137.90 ± 79.23 | <0.0001 |
| Days of illness before admission | 5.65± 1.62 | 5.77 ± 2.98 | 0.852 |
| Laboratory baseline | |||
| White blood cells (cells/µL) | 6664.00 ± 3841 | 13897 ± 1231 | <0.0001 |
| Neutrophils (cells/µL) | 68.34 ± 16.62 | 79.80 ± 13.01 | 0.012 |
| Lymphocytes (cells/µL) | 27.58 ± 19.17 | 12.02 ± 13.30 | 0.001 |
| Platelets (cells/µL) | 207333 ± 88056 | 271737 ± 139869 | 0.100 |
| C-reactive protein (mg/L) | 7.05 ± 7.57 | 13.67 ± 11.52 | 0.025 |
| Troponin (ng/mL) | 37.58 ± 109.70 | 311.80 ± 845.90 | 0.114 |
| Urea (mg/dL) | 34.89 ± 24.78 | 84.16 ± 94.24 | <0.001 |
| Creatinine (mg/dL) | 0.82 ± 0.19 | 1.10 ± 0.88 | 0.368 |
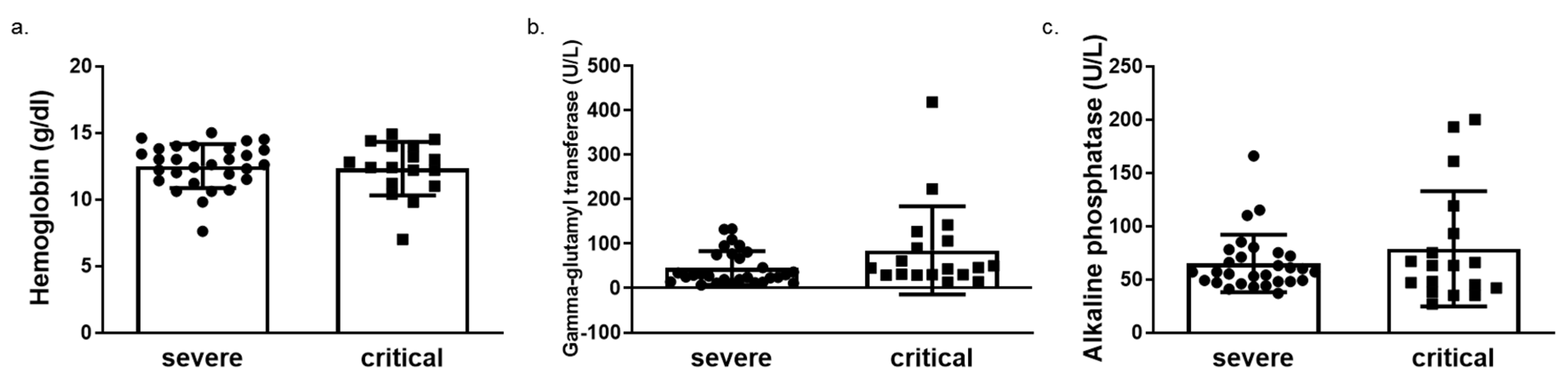
| Aspartate aminotransferase (U/L) | 43.33 ± 37.50 | 150.70 ± 419.20 | 0.237 |
| Alanine transaminase (U/L) | 34.76 ± 22.94 | 62.17 ± 74.60 | 0.002 |
| Gamma-Glutamyltransferase (U/L) | 46.37 ± 39.22 | 83.26 ± 96.73 | 0.062 |
| Lactate Dehydrogenase (U/L) | 306.8 ± 128.9 | 535.6 ± 552.1 | 0.021 |
| Albumin (g/dL) | 3.846 ± 0.409 | 3.210 ± 0.506 | <0.0001 |
| Days of hospital stay | 8.00 ± 4.019 | 18.80 ± 14.10 | <0.0001 |
| Survival | 25 (92.59%) | 12 (60.00%) | 0.026 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Detsika, M.G.; Nikitopoulou, I.; Veroutis, D.; Vassiliou, A.G.; Jahaj, E.; Tsipilis, S.; Athanassiou, N.; Gakiopoulou, H.; Gorgoulis, V.G.; Dimopoulou, I.; et al. Increase of HO-1 Expression in Critically Ill COVID-19 Patients Is Associated with Poor Prognosis and Outcome. Antioxidants 2022, 11, 1300. https://doi.org/10.3390/antiox11071300
Detsika MG, Nikitopoulou I, Veroutis D, Vassiliou AG, Jahaj E, Tsipilis S, Athanassiou N, Gakiopoulou H, Gorgoulis VG, Dimopoulou I, et al. Increase of HO-1 Expression in Critically Ill COVID-19 Patients Is Associated with Poor Prognosis and Outcome. Antioxidants. 2022; 11(7):1300. https://doi.org/10.3390/antiox11071300
Chicago/Turabian StyleDetsika, Maria G., Ioanna Nikitopoulou, Dimitris Veroutis, Alice G. Vassiliou, Edison Jahaj, Stamatis Tsipilis, Nikolaos Athanassiou, Hariklia Gakiopoulou, Vassilis G. Gorgoulis, Ioanna Dimopoulou, and et al. 2022. "Increase of HO-1 Expression in Critically Ill COVID-19 Patients Is Associated with Poor Prognosis and Outcome" Antioxidants 11, no. 7: 1300. https://doi.org/10.3390/antiox11071300
APA StyleDetsika, M. G., Nikitopoulou, I., Veroutis, D., Vassiliou, A. G., Jahaj, E., Tsipilis, S., Athanassiou, N., Gakiopoulou, H., Gorgoulis, V. G., Dimopoulou, I., Orfanos, S. E., & Kotanidou, A. (2022). Increase of HO-1 Expression in Critically Ill COVID-19 Patients Is Associated with Poor Prognosis and Outcome. Antioxidants, 11(7), 1300. https://doi.org/10.3390/antiox11071300