
Colon-Targeted eNAMPT-Specific Peptide Systems for Treatment of DSS-Induced Acute and Chronic Colitis in Mouse

 and
and 
Abstract
1. Introduction
2. Materials and Methdos
2.1. Analysis of Public Single-Cell RNA-Seq Data for Ulcerative Colitis
- (1)
- DEG: DEGs were identified separately for the two datasets with a p-value cutoff of 0.05. Rather than using an adjusted p-value, we used a non-adjusted one to obtain as many DEGs as possible.
- (2)
- Selection of common DEGs: We then selected those DEGs that have the same sign of log-fold change in the two datasets and then took the intersection of the two sets of DEGs, which is used for the GSEA.
- (3)
- GSEAs with common DEGs: GSEA analyses were performed with the selected DEGs with the log-fold changes for SCP259, since it might provide a better result when using a greater number of cells.
2.2. Mice, and Tissue-Culture
2.3. Reagents and Antibodies
2.4. Plasmid Construction
2.5. Peptides
2.6. Recombinant Protein
2.7. GST Pulldown, Immunoblot, and Immunoprecipitation Analysis
2.8. Flow Cytometry
2.9. Enzyme-Linked Immunosorbent Assay
2.10. Lentiviral shRNA Production
2.11. In Vivo Lentivirus Transduction
2.12. Mouse Model of Colitis
2.13. Clinical Score and Histology
2.14. Statistical Analysis
3. Result
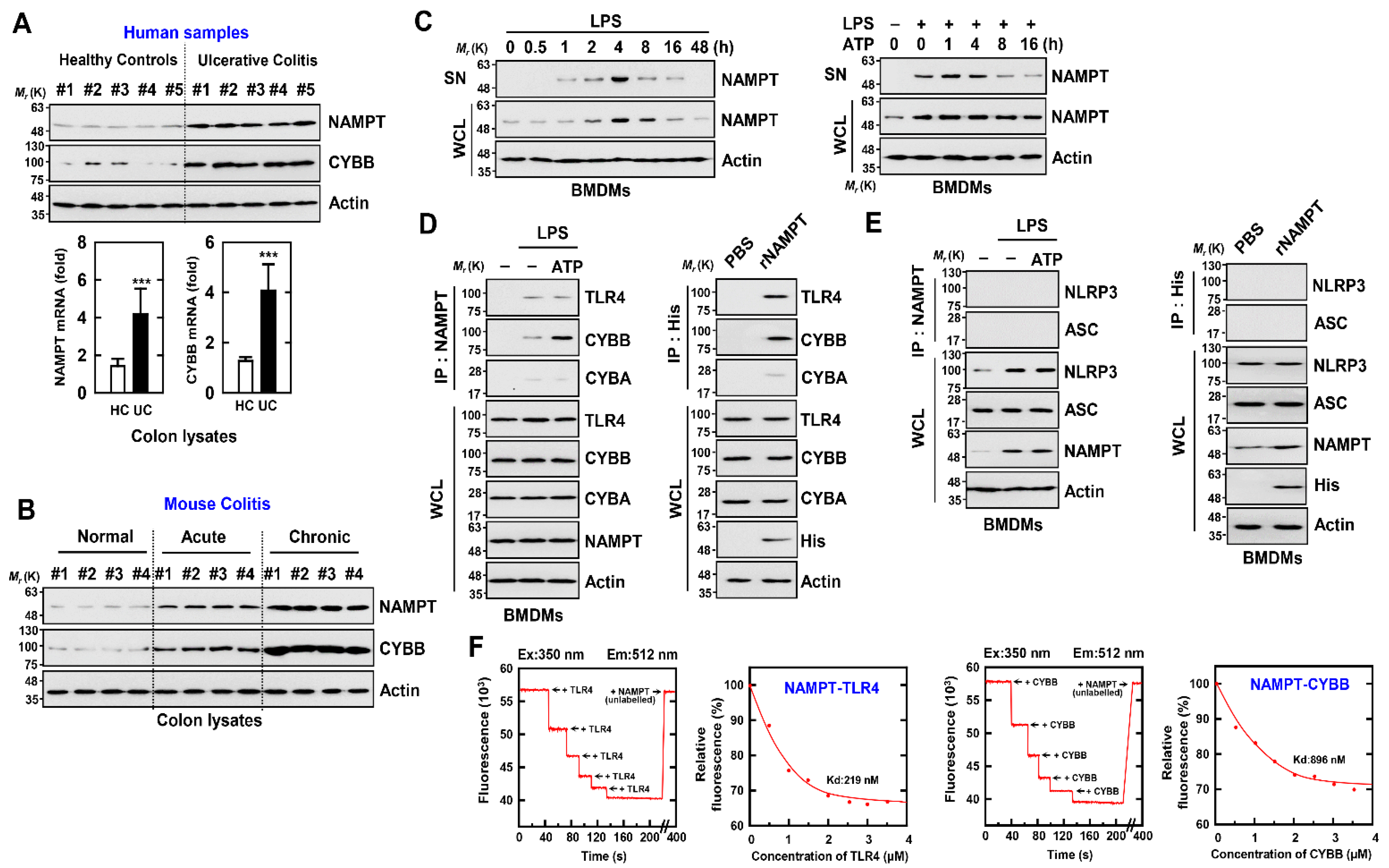
3.1. Overexpressed eNAMPT and CYBB May Cause Chronic Inflammation in IBD
3.2. eNAMPT Interacts with CYBB and TLR4
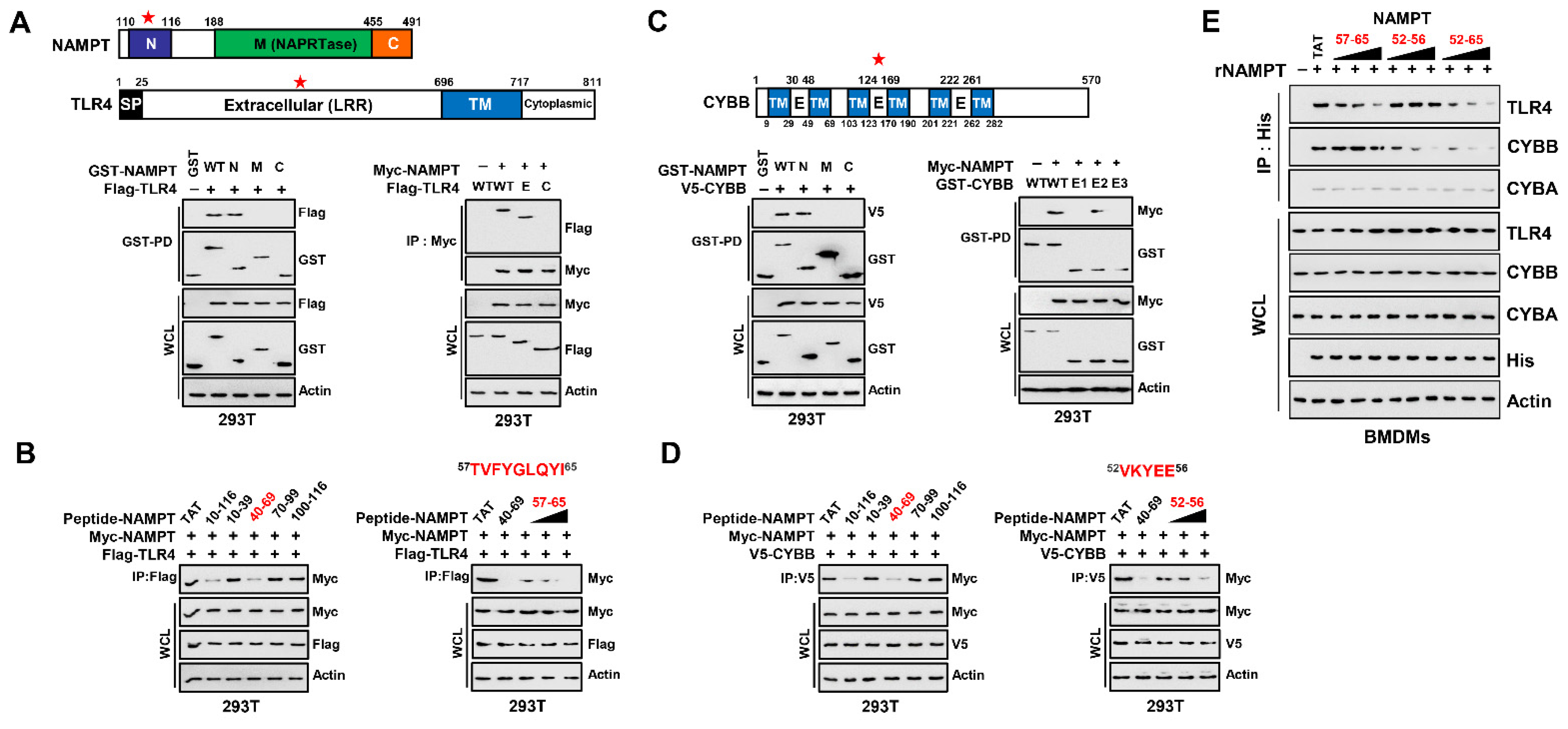
3.3. NAMPT Amino Acid Sequence Essential for Binding TLR4 and CYBB
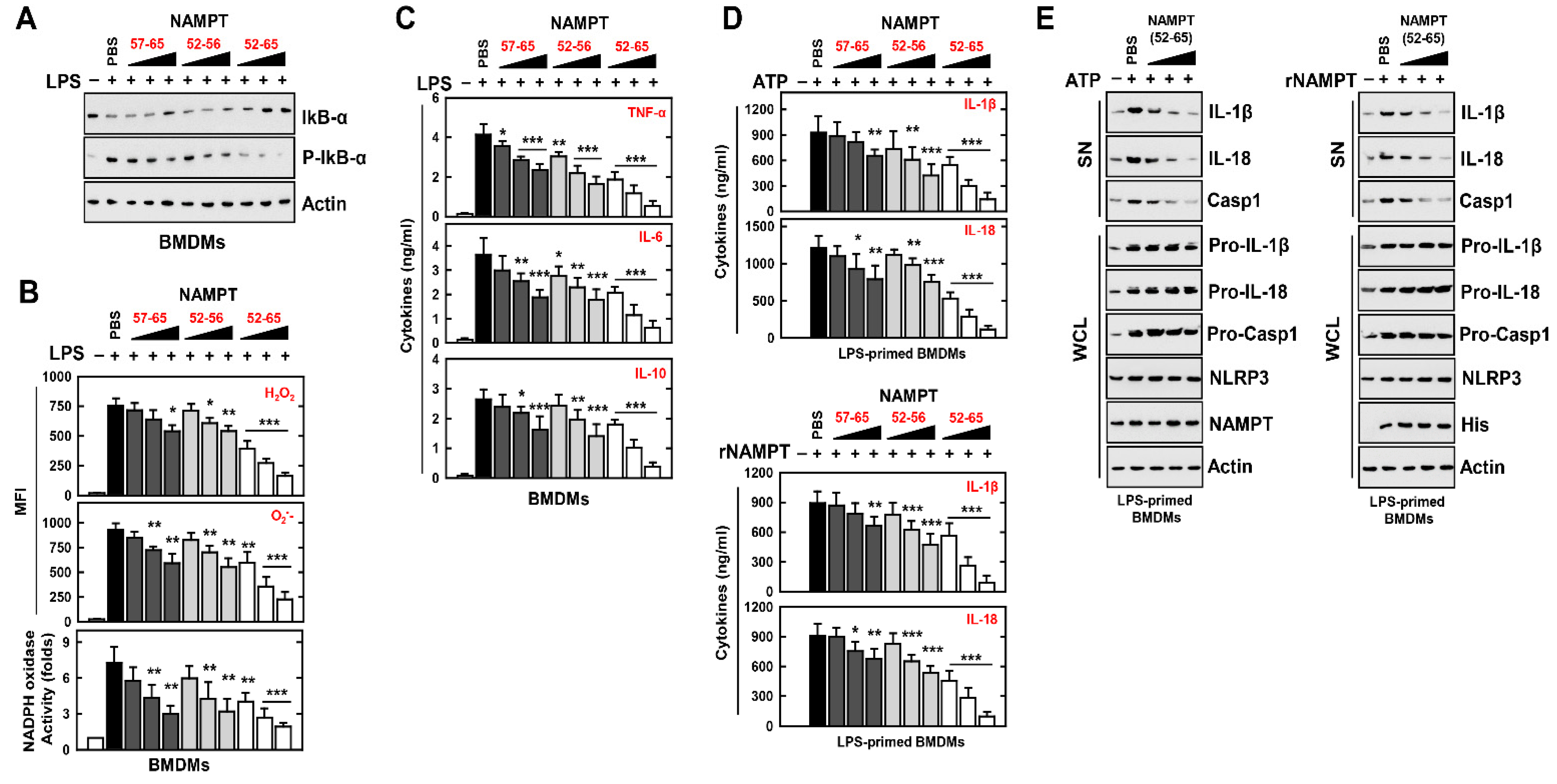
3.4. An NAMPT Peptide Inhibits Activation of the NLRP3 Inflammasome
3.5. Recombinant Multifunctional CT-NAMPT Protein Designed to Target the Pathologically Inflamed Colon
3.6. rCT-NAMPT Relieves Acute and Chronic DSS-Induced Colitis in a Mouse Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nielsen, O.H. New strategies for treatment of inflammatory bowel disease. Front. Med. (Lausanne) 2014, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.G.; Ng, S.C. Understanding and Preventing the Global Increase of Inflammatory Bowel Disease. Gastroenterology 2017, 152, 313–321.e2. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, O.H.; Ainsworth, M.A. Tumor necrosis factor inhibitors for inflammatory bowel disease. N. Engl. J. Med. 2013, 369, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Vulliemoz, M.; Brand, S.; Juillerat, P.; Mottet, C.; Ben-Horin, S.; Michetti, P.; on behalf of Swiss IBDnet, an official working group of the Swiss Society of Gastroenterology. TNF-Alpha Blockers in Inflammatory Bowel Diseases: Practical Recommendations and a User’s Guide: An Update. Digestion 2020, 101 (Suppl. S1), 16–26. [Google Scholar] [CrossRef] [PubMed]
- Chiarugi, A.; Dolle, C.; Felici, R.; Ziegler, M. The NAD metabolome—A key determinant of cancer cell biology. Nat. Rev. Cancer 2012, 12, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Carbone, F.; Liberale, L.; Bonaventura, A.; Vecchie, A.; Casula, M.; Cea, M.; Monacelli, F.; Caffa, I.; Bruzzone, S.; Montecucco, F.; et al. Regulation and Function of Extracellular Nicotinamide Phosphoribosyltransferase/Visfatin. Compr. Physiol. 2017, 7, 603–621. [Google Scholar] [PubMed]
- Travelli, C.; Colombo, G.; Mola, S.; Genazzani, A.A.; Porta, C. NAMPT: A pleiotropic modulator of monocytes and macrophages. Pharm. Res. 2018, 135, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.M.; Lee, A.Y.; Hwang, S.M.; Ha, Y.J.; Kim, M.J.; Min, S.; Hwang, W.; Yoon, G.; Kwon, S.M.; Woo, H.G.; et al. NAMPT mitigates colitis severity by supporting redox-sensitive activation of phagocytosis in inflammatory macrophages. Redox. Biol. 2022, 50, 102237. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Nabeshima, Y. Nampt/PBEF/Visfatin: A new player in beta cell physiology and in metabolic diseases? Cell Metab. 2007, 6, 341–343. [Google Scholar] [CrossRef] [PubMed]
- Garten, A.; Schuster, S.; Penke, M.; Gorski, T.; de Giorgis, T.; Kiess, W. Physiological and pathophysiological roles of NAMPT and NAD metabolism. Nat. Rev. Endocrinol. 2015, 11, 535–546. [Google Scholar] [CrossRef]
- Galli, U.; Colombo, G.; Travelli, C.; Tron, G.C.; Genazzani, A.A.; Grolla, A.A. Recent Advances in NAMPT Inhibitors: A Novel Immunotherapic Strategy. Front. Pharm. 2020, 11, 656. [Google Scholar] [CrossRef] [PubMed]
- Imai, S. Nicotinamide phosphoribosyltransferase (Nampt): A link between NAD biology, metabolism, and diseases. Curr. Pharm. Des. 2009, 15, 20–28. [Google Scholar] [CrossRef]
- Audrito, V.; Manago, A.; Gaudino, F.; Deaglio, S. Targeting metabolic reprogramming in metastatic melanoma: The key role of nicotinamide phosphoribosyltransferase (NAMPT). Semin. Cell Dev. Biol. 2020, 98, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Audrito, V.; Messana, V.G.; Deaglio, S. NAMPT and NAPRT: Two Metabolic Enzymes With Key Roles in Inflammation. Front. Oncol. 2020, 10, 358. [Google Scholar] [CrossRef] [PubMed]
- Starr, A.E.; Deeke, S.A.; Ning, Z.; Chiang, C.K.; Zhang, X.; Mottawea, W.; Singleton, R.; Benchimol, E.I.; Wen, M.; Mack, D.R.; et al. Proteomic analysis of ascending colon biopsies from a paediatric inflammatory bowel disease inception cohort identifies protein biomarkers that differentiate Crohn’s disease from UC. Gut 2017, 66, 1573–1583. [Google Scholar] [CrossRef]
- Colombo, G.; Clemente, N.; Zito, A.; Bracci, C.; Colombo, F.S.; Sangaletti, S.; Jachetti, E.; Ribaldone, D.G.; Caviglia, G.P.; Pastorelli, L.; et al. Neutralization of extracellular NAMPT (nicotinamide phosphoribosyltransferase) ameliorates experimental murine colitis. J. Mol. Med. (Berl.) 2020, 98, 595–612. [Google Scholar] [CrossRef]
- Camp, S.M.; Ceco, E.; Evenoski, C.L.; Danilov, S.M.; Zhou, T.; Chiang, E.T.; Moreno-Vinasco, L.; Mapes, B.; Zhao, J.; Gursoy, G.; et al. Unique Toll-Like Receptor 4 Activation by NAMPT/PBEF Induces NFkappaB Signaling and Inflammatory Lung Injury. Sci. Rep. 2015, 5, 13135. [Google Scholar] [CrossRef] [PubMed]
- Manago, A.; Audrito, V.; Mazzola, F.; Sorci, L.; Gaudino, F.; Gizzi, K.; Vitale, N.; Incarnato, D.; Minazzato, G.; Ianniello, A.; et al. Extracellular nicotinate phosphoribosyltransferase binds Toll like receptor 4 and mediates inflammation. Nat. Commun. 2019, 10, 4116. [Google Scholar] [CrossRef] [PubMed]
- Molugu, T.R.; Oita, R.C.; Chawla, U.; Camp, S.M.; Brown, M.F.; Garcia, J.G.N. Nicotinamide phosphoribosyltransferase purification using SUMO expression system. Anal. Biochem. 2020, 598, 113597. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Lauritzen, K.H.; Olsen, M.B.; Dahl, T.B.; Ranheim, T.; Ahmed, M.S.; Attramadal, H.; Aukrust, P.; Halvorsen, B.; Nyman, T.A.; et al. Low Cellular NAD(+) Compromises Lipopolysaccharide-Induced Inflammatory Responses via Inhibiting TLR4 Signal Transduction in Human Monocytes. J. Immunol. 2019, 203, 1598–1608. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Tardivel, A.; Thorens, B.; Choi, I.; Tschopp, J. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat. Immunol. 2010, 11, 136–140. [Google Scholar] [CrossRef]
- Abais, J.M.; Xia, M.; Zhang, Y.; Boini, K.M.; Li, P.L. Redox regulation of NLRP3 inflammasomes: ROS as trigger or effector? Antioxid. Redox. Signal. 2015, 22, 1111–1129. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Mu, Y.; Song, Y.; Xie, J.; Yu, H.; Gao, S.; Li, S.; Peng, H.; Zhou, Y.; Lu, W. A new peptide ligand for colon cancer targeted delivery of micelles. Drug Deliv. 2016, 23, 1763–1772. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Bajracharya, R.; Min, J.Y.; Han, J.W.; Park, B.J.; Han, H.K. Strategic Approaches for Colon Targeted Drug Delivery: An Overview of Recent Advancements. Pharmaceutics 2020, 12, 68. [Google Scholar] [CrossRef] [PubMed]
- Smillie, C.S.; Biton, M.; Ordovas-Montanes, J.; Sullivan, K.M.; Burgin, G.; Graham, D.B.; Herbst, R.H.; Rogel, N.; Slyper, M.; Waldman, J.; et al. Intra- and Inter-cellular Rewiring of the Human Colon during Ulcerative Colitis. Cell 2019, 178, 714–730.e22. [Google Scholar] [CrossRef] [PubMed]
- Uzzan, M.; Martin, J.C.; Mesin, L.; Livanos, A.E.; Castro-Dopico, T.; Huang, R.; Petralia, F.; Magri, G.; Kumar, S.; Zhao, Q.; et al. Ulcerative colitis is characterized by a plasmablast-skewed humoral response associated with disease activity. Nat. Med. 2022, 28, 766–779. [Google Scholar] [CrossRef] [PubMed]
- Devlin, J.C.; Axelrad, J.; Hine, A.M.; Chang, S.; Sarkar, S.; Lin, J.D.; Ruggles, K.V.; Hudesman, D.; Cadwell, K.; Loke, P. Single-Cell Transcriptional Survey of Ileal-Anal Pouch Immune Cells From Ulcerative Colitis Patients. Gastroenterology 2021, 160, 1679–1693. [Google Scholar] [CrossRef]
- Boland, B.S.; He, Z.; Tsai, M.S.; Olvera, J.G.; Omilusik, K.D.; Duong, H.G.; Kim, E.S.; Limary, A.E.; Jin, W.; Milner, J.J.; et al. Heterogeneity and clonal relationships of adaptive immune cells in ulcerative colitis revealed by single-cell analyses. Sci. Immunol. 2020, 5, eabb4432. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Liao, J.; Li, C.; Lu, X.; Cheng, J.; Fan, X. CellTalkDB: A manually curated database of ligand-receptor interactions in humans and mice. Brief. Bioinform. 2021, 22, bbaa269. [Google Scholar] [CrossRef] [PubMed]
- Koh, H.J.; Kim, Y.R.; Kim, J.S.; Yun, J.S.; Jang, K.; Yang, C.S. Toxoplasma gondii GRA7-Targeted ASC and PLD1 Promote Antibacterial Host Defense via PKCalpha. PLoS Pathog. 2017, 13, e1006126. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, D.; Kim, S.; Lee, D.; Mun, S.J.; Cho, E.; Son, W.; Jang, K.; Yang, C.S. Mycobacterium tuberculosis Rv2626c-derived peptide as a therapeutic agent for sepsis. EMBO Mol. Med. 2020, 12, e12497. [Google Scholar] [CrossRef]
- Yang, C.S.; Lee, J.S.; Rodgers, M.; Min, C.K.; Lee, J.Y.; Kim, H.J.; Lee, K.H.; Kim, C.J.; Oh, B.; Zandi, E.; et al. Autophagy protein Rubicon mediates phagocytic NADPH oxidase activation in response to microbial infection or TLR stimulation. Cell Host Microbe 2012, 11, 264–276. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, D.; Kim, D.; Mun, S.J.; Cho, E.; Son, W.; Yang, C.S. Toxoplasma gondii GRA8-derived peptide immunotherapy improves tumor targeting of colorectal cancer. Oncotarget 2020, 11, 62–73. [Google Scholar] [CrossRef]
- Kim, J.S.; Mun, S.J.; Cho, E.; Kim, D.; Son, W.; Jeon, H.I.; Kim, H.K.; Jang, K.; Yang, C.S. Toxoplasma gondii GRA9 Regulates the Activation of NLRP3 Inflammasome to Exert Anti-Septic Effects in Mice. Int. J. Mol. Sci. 2020, 21, 8437. [Google Scholar] [CrossRef]
- Kim, J.S.; Cho, E.; Mun, S.J.; Kim, S.; Kim, S.Y.; Kim, D.G.; Son, W.; Jeon, H.I.; Kim, H.K.; Jeong, Y.J.; et al. Multi-Functional MPT Protein as a Therapeutic Agent against Mycobacterium tuberculosis. Biomedicines 2021, 9, 545. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, Y.R.; Jang, S.; Wang, S.G.; Cho, E.; Mun, S.J.; Jeon, H.I.; Kim, H.K.; Min, S.J.; Yang, C.S. Mito-TIPTP Increases Mitochondrial Function by Repressing the Rubicon-p22phox Interaction in Colitis-Induced Mice. Antioxidants 2021, 10, 1954. [Google Scholar] [CrossRef]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef]
- Pandey, A.; Shen, C.; Feng, S.; Man, S.M. Cell biology of inflammasome activation. Trends Cell Biol. 2021, 31, 924–939. [Google Scholar] [CrossRef]
- Bolhassani, A.; Jafarzade, B.S.; Mardani, G. In vitro and in vivo delivery of therapeutic proteins using cell penetrating peptides. Peptides 2017, 87, 50–63. [Google Scholar] [CrossRef]
- Bernhofer, M.; Dallago, C.; Karl, T.; Satagopam, V.; Heinzinger, M.; Littmann, M.; Olenyi, T.; Qiu, J.; Schutze, K.; Yachdav, G.; et al. PredictProtein-Predicting Protein Structure and Function for 29 Years. Nucleic Acids Res. 2021, 49, W535–W540. [Google Scholar] [CrossRef]
- Nakahara, H.; Nomizu, M.; Akiyama, S.K.; Yamada, Y.; Yeh, Y.; Chen, W.T. A mechanism for regulation of melanoma invasion. Ligation of alpha6beta1 integrin by laminin G peptides. J. Biol. Chem. 1996, 271, 27221–27224. [Google Scholar] [CrossRef]
- Amit, I.; Winter, D.R.; Jung, S. The role of the local environment and epigenetics in shaping macrophage identity and their effect on tissue homeostasis. Nat. Immunol. 2016, 17, 18–25. [Google Scholar] [CrossRef]
- Colombo, G.; Travelli, C.; Porta, C.; Genazzani, A.A. Extracellular nicotinamide phosphoribosyltransferase boosts IFNgamma-induced macrophage polarization independently of TLR4. iScience 2022, 25, 104147. [Google Scholar] [CrossRef]
- Ratnayake, D.; Nguyen, P.D.; Rossello, F.J.; Wimmer, V.C.; Tan, J.L.; Galvis, L.A.; Julier, Z.; Wood, A.J.; Boudier, T.; Isiaku, A.I.; et al. Macrophages provide a transient muscle stem cell niche via NAMPT secretion. Nature 2021, 591, 281–287. [Google Scholar] [CrossRef]
- Wells, C.A.; Chalk, A.M.; Forrest, A.; Taylor, D.; Waddell, N.; Schroder, K.; Himes, S.R.; Faulkner, G.; Lo, S.; Kasukawa, T.; et al. Alternate transcription of the Toll-like receptor signaling cascade. Genome Biol. 2006, 7, R10. [Google Scholar] [CrossRef]
- Guo, T.; Hou, D.; Yu, D. Bioinformatics analysis of gene expression profile data to screen key genes involved in intracranial aneurysms. Mol. Med. Rep. 2019, 20, 4415–4424. [Google Scholar] [CrossRef]
- Bakshi, H.A.; Quinn, G.A.; Aljabali, A.A.A.; Hakkim, F.L.; Farzand, R.; Nasef, M.M.; Abuglela, N.; Ansari, P.; Mishra, V.; Serrano-Aroca, A.; et al. Exploiting the Metabolism of the Gut Microbiome as a Vehicle for Targeted Drug Delivery to the Colon. Pharmaceuticals 2021, 14, 1211. [Google Scholar] [CrossRef]
- Arevalo-Perez, R.; Maderuelo, C.; Lanao, J.M. Recent advances in colon drug delivery systems. J. Control. Release 2020, 327, 703–724. [Google Scholar]
- Garcia, M.A.; Varum, F.; Al-Gousous, J.; Hofmann, M.; Page, S.; Langguth, P. In Vitro Methodologies for Evaluating Colon-Targeted Pharmaceutical Products and Industry Perspectives for Their Applications. Pharmaceutics 2022, 14, 291. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Tissue | Condition and Number of Samples | Total Number of Cells | Cell Type Annotation | ||
|---|---|---|---|---|---|---|
| Healthy | Inflamed | Non-Inflamed | ||||
| SCP259 | Colon (LP and Epi) | 12 | 18 | 18 | 365 K | Available |
| GSE182270 | Colon (LP) | 4 | 5 | - | 32 K | Not available |
| Genes | SCP259 (# Macrophages: 13888) | GSE182270 (# Macrophages: 555) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| p.val | p.val.adj | logFC | pct.test | pct.ctrl | p.val | p.val.adj | logFC | pct.test | pct.ctrl | |
| NAMPT | 2.64 × 10−13 | 4.80 × 10−09 | 0.72 | 0.36 | 0.34 | 0.0011 | 1 | 0.32 | 0.88 | 0.82 |
| CYBB | 7.57 × 10−21 | 1.37 × 10−16 | 0.71 | 0.39 | 0.34 | 4.19 × 10−06 | 0.141 | 0.62 | 0.66 | 0.48 |
| NCF1 | 3.79 × 10−65 | 6.87 × 10−61 | −0.43 * | 0.29 | 0.53 | 0.049602 | 1 | −0.27 * | 0.39 | 0.51 |
| NCF2 | 7.77 × 10−11 | 1.41 × 10−06 | 0.53 | 0.24 | 0.20 | 0.001618 | 1 | 0.54 | 0.51 | 0.37 |
| NCF4 | 1.7 × 10−132 | 3.1 × 10−128 | −0.86 * | 0.21 | 0.49 | 0.001578 | 1 | −0.53 * | 0.33 | 0.47 |
| P2RY6 | 6.00 × 10−87 | 1.1 × 10−82 | −0.67 | 0.09 | 0.26 | 2.23 × 10−05 | 0.752 | −0.59 | 0.21 | 0.41 |
| SOD2 | 6.58 × 10−12 | 1.2 × 10−07 | 0.489 | 0.51 | 0.522 | 5.89 × 10−19 | 1.9 × 10−14 | 1.074 | 0.901 | 0.653 |
| N × AT1 | 0 | 0 | 1.171 | 0.798 | 0.585 | 3.21 × 10−12 | 1.1 × 10−07 | 0.467 | 0.971 | 0.874 |
| HIF1A | 1.63 × 10−16 | 2.9 × 10−12 | 0.790 | 0.196 | 0.135 | 0.002 | 1 | 0.624 | 0.48 | 0.358 |
| TXNIP | 7.71 × 10−70 | 1.4 × 10−65 | 0.985 | 0.455 | 0.312 | 0.025 | 1 | −0.323 * | 0.417 | 0.505 |
| TXN | Not available | 3.74 × 10−05 | 1 | 0.569 | 0.764 | 0.695 | ||||
| NLRP3 | 1.54 × 10−05 | 0.279 | 0.420 | 0.067 | 0.044 | Not available | ||||
| CASP1 | 0.026 | 1 | 0.298 | 0.329 | 0.346 | Not available | ||||
| IL1B | Not available | 0.008 | 1 | 0.402 | 0.728 | 0.705 | ||||
| NFKB1 | 0.038 | 1 | 0.504 | 0.159 | 0.153 | 0.013 | 1 | 0.543 | 0.422 | 0.316 |
| TNFAIP8 | 0.024 | 1 | 0.377 | 0.156 | 0.146 | 0.024 | 1 | 0.473 | 0.453 | 0.368 |
| TNFAIP8L2 | 6.22 × 10−18 | 1.13 × 10−13 | −0.306 | 0.131 | 0.212 | 0.001 | 1 | −0.599 | 0.253 | 0.421 |
| TNFRSF1B | 3.36 × 10−17 | 6.11 × 10−13 | 0.714 | 0.277 | 0.218 | 0.011 | 1 | 0.611 | 0.436 | 0.368 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-S.; Kim, H.K.; Kim, M.; Jang, S.; Cho, E.; Mun, S.-J.; Lee, J.; Hong, D.; Yoon, S.; Yang, C.-S. Colon-Targeted eNAMPT-Specific Peptide Systems for Treatment of DSS-Induced Acute and Chronic Colitis in Mouse. Antioxidants 2022, 11, 2376. https://doi.org/10.3390/antiox11122376
Kim J-S, Kim HK, Kim M, Jang S, Cho E, Mun S-J, Lee J, Hong D, Yoon S, Yang C-S. Colon-Targeted eNAMPT-Specific Peptide Systems for Treatment of DSS-Induced Acute and Chronic Colitis in Mouse. Antioxidants. 2022; 11(12):2376. https://doi.org/10.3390/antiox11122376
Chicago/Turabian StyleKim, Jae-Sung, Hyo Keun Kim, Minsoo Kim, Sein Jang, Euni Cho, Seok-Jun Mun, Joongho Lee, Dawon Hong, Seokhyun Yoon, and Chul-Su Yang. 2022. "Colon-Targeted eNAMPT-Specific Peptide Systems for Treatment of DSS-Induced Acute and Chronic Colitis in Mouse" Antioxidants 11, no. 12: 2376. https://doi.org/10.3390/antiox11122376
APA StyleKim, J.-S., Kim, H. K., Kim, M., Jang, S., Cho, E., Mun, S.-J., Lee, J., Hong, D., Yoon, S., & Yang, C.-S. (2022). Colon-Targeted eNAMPT-Specific Peptide Systems for Treatment of DSS-Induced Acute and Chronic Colitis in Mouse. Antioxidants, 11(12), 2376. https://doi.org/10.3390/antiox11122376