
Paraoxonase-1 and -3 Protein Expression in the Brain of the Tg2576 Mouse Model of Alzheimer’s Disease

 ,
, 
 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Antibodies
2.3. Peroxidase-Based Immunostaining
2.4. Fluorescence-Based Immunostaining
3. Results
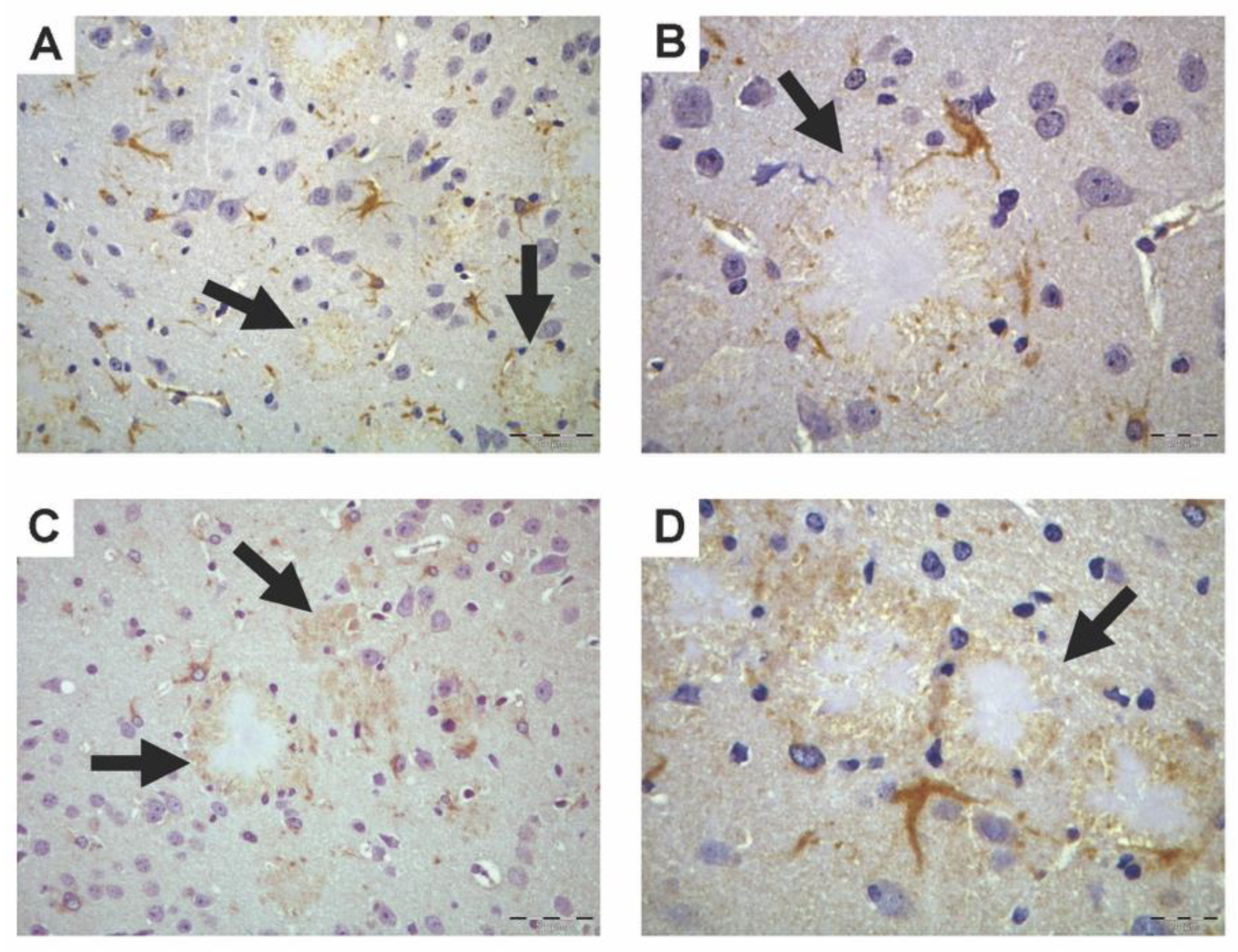
3.1. PON1 and PON3 Peroxidase-Based Immunohistochemistry
3.2. Fluorescence-Based Immunohistochemistry
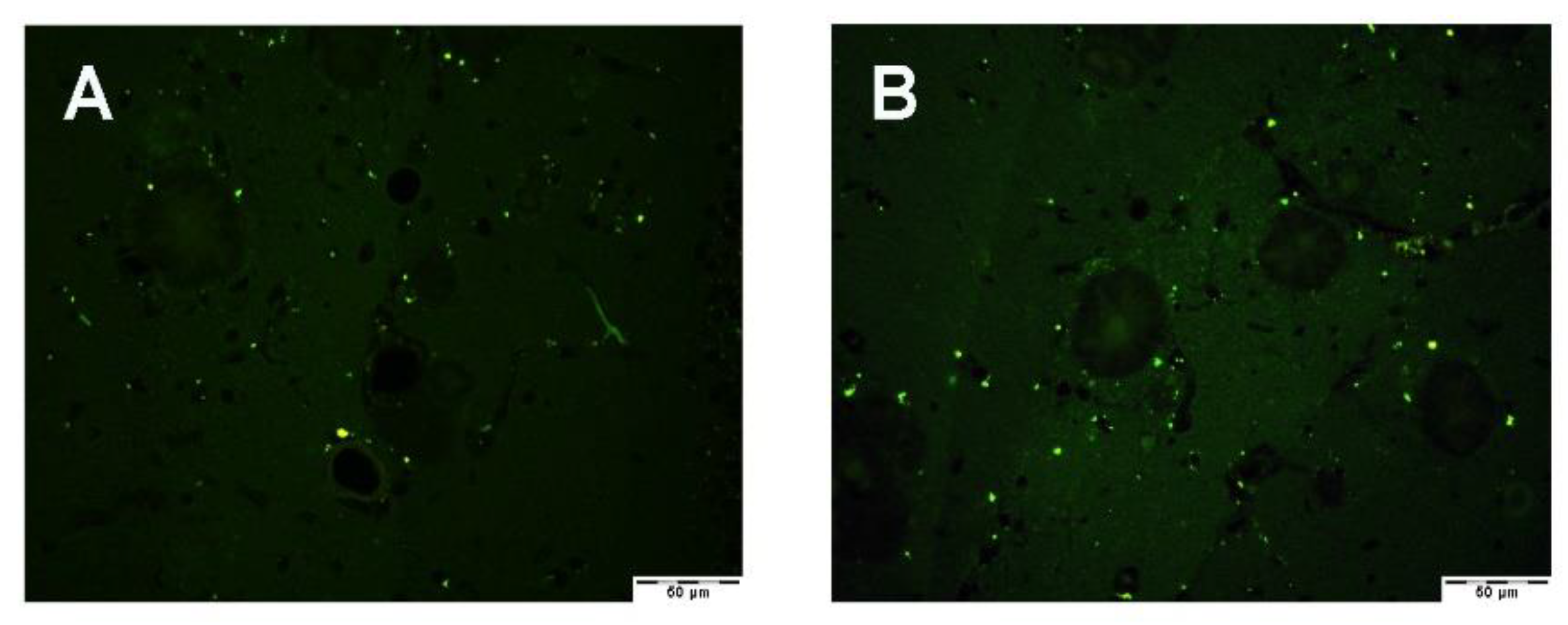
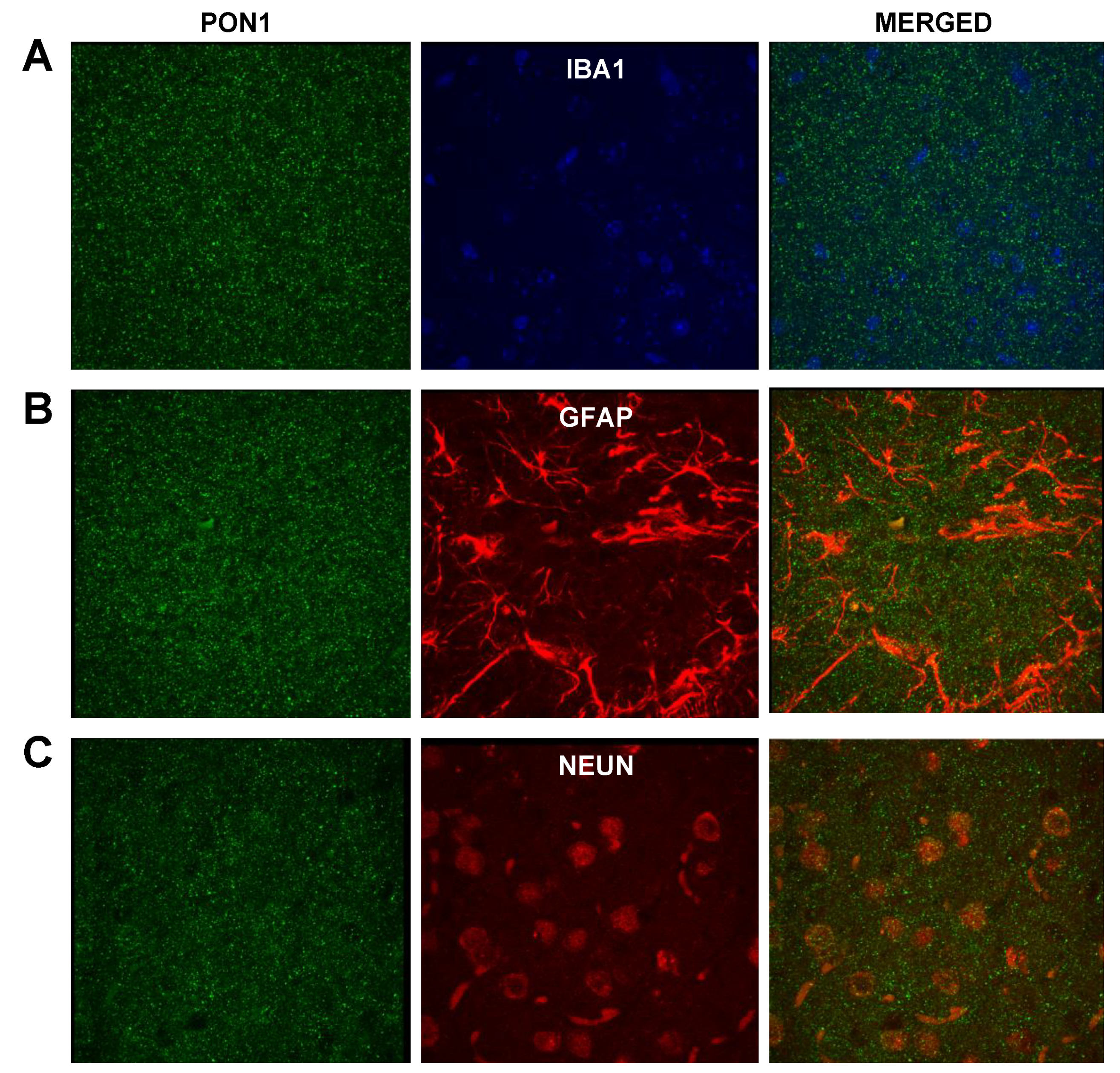
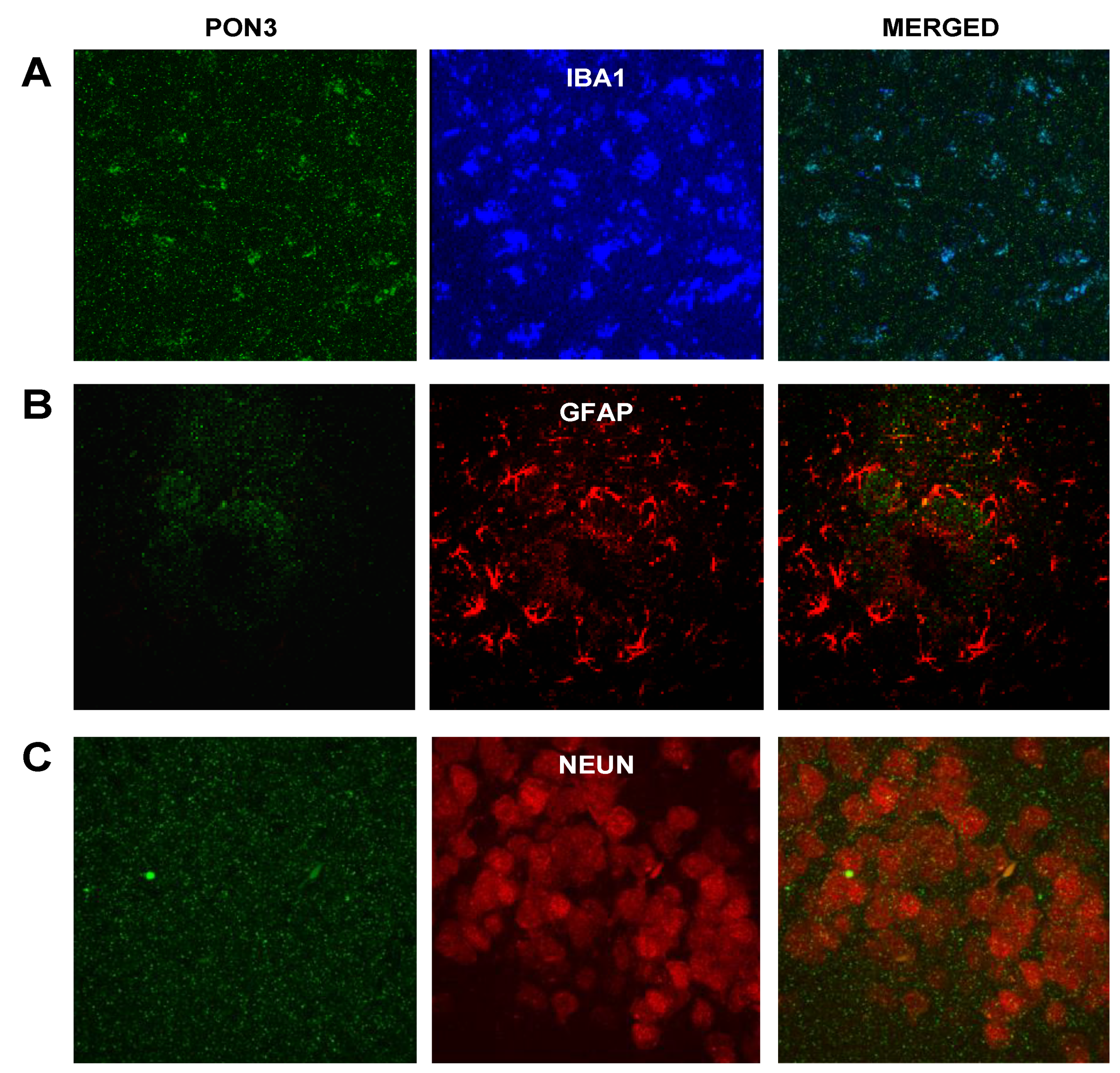
3.2.1. PON1 and PON3 Immunofluorescence
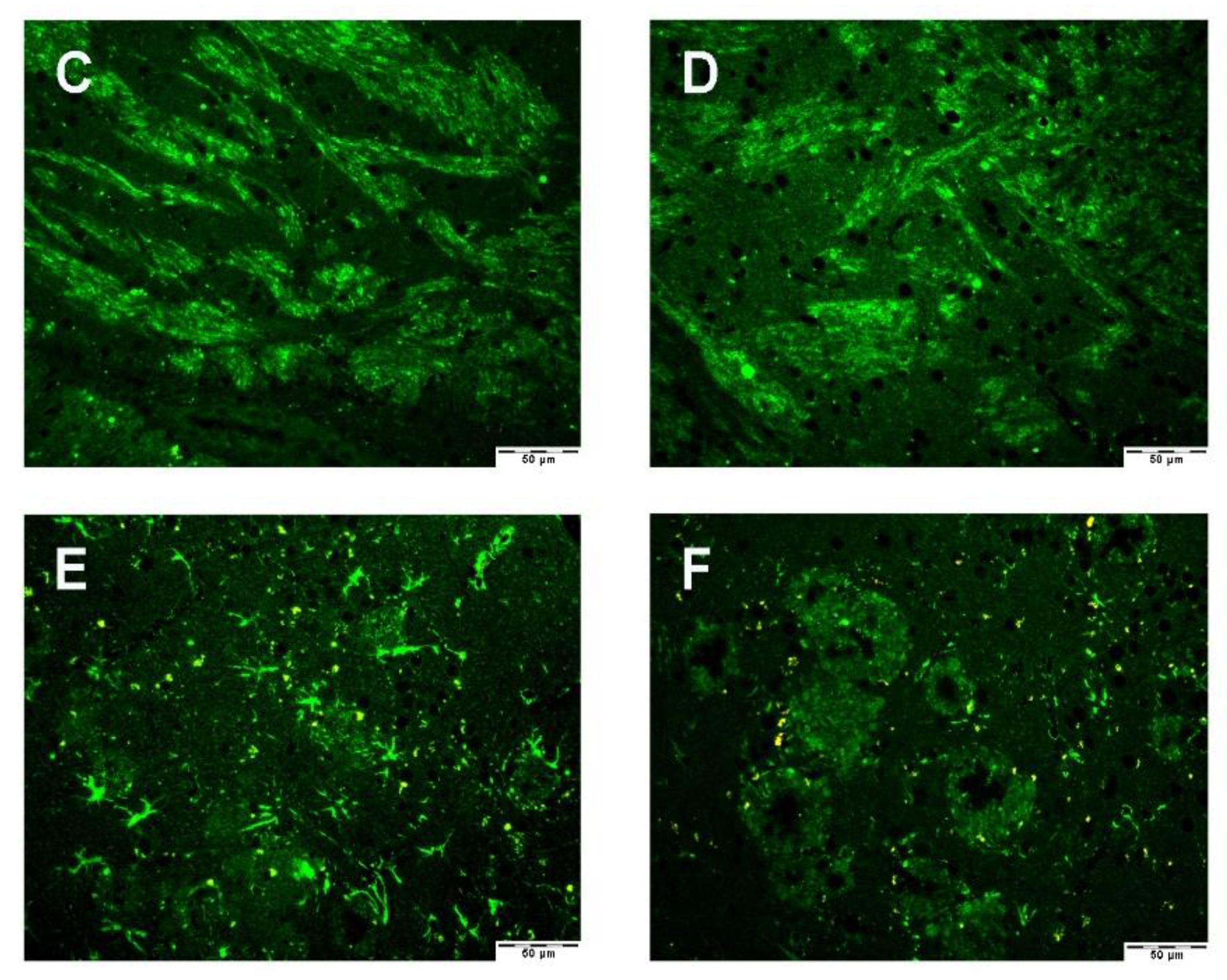
3.2.2. Astrocyte–Microglia–Neuron “Triad” Immunofluorescence
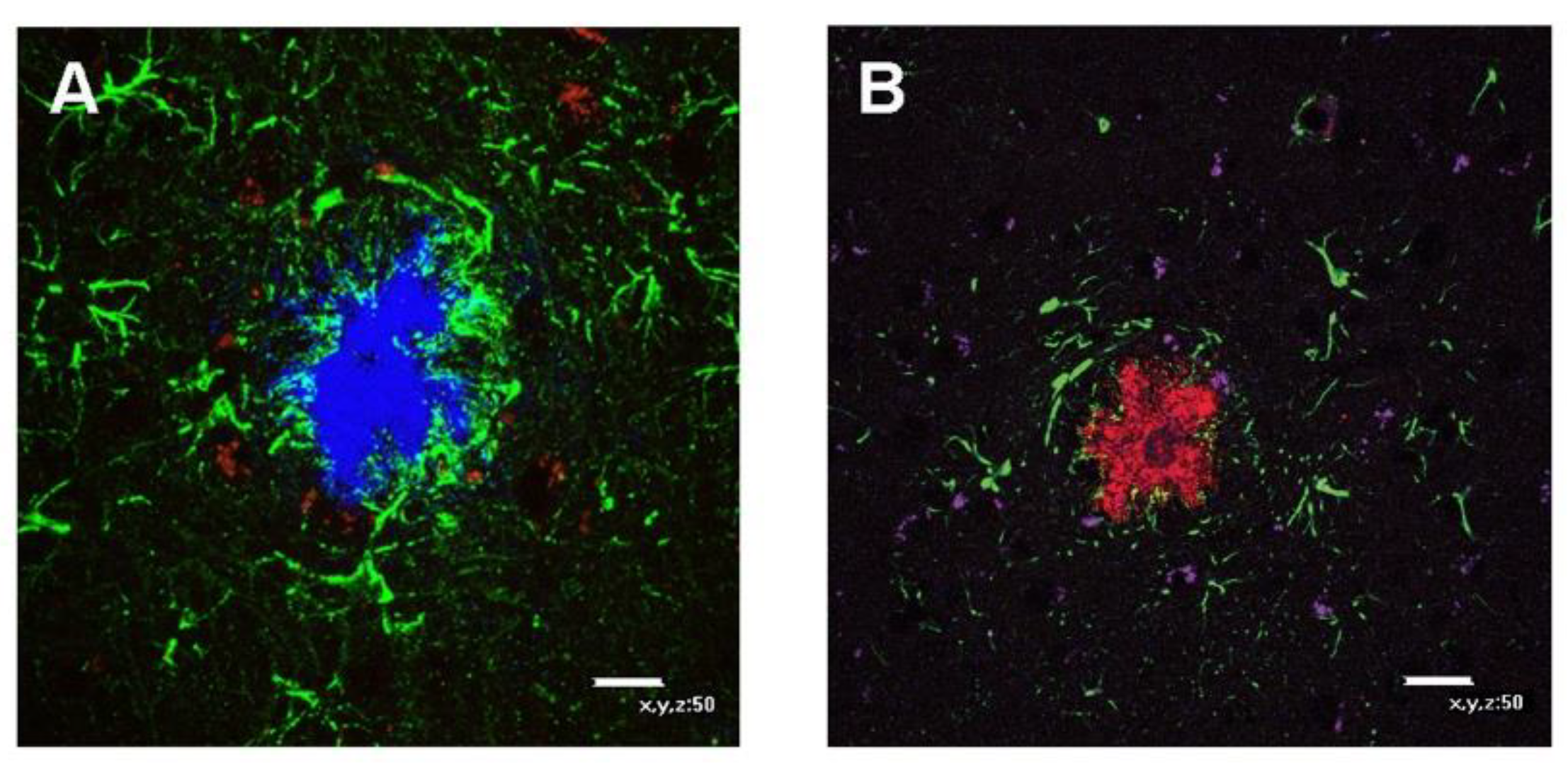
3.2.3. PON1 and Astrocyte–Microglia–Neuron “Triad” Double Immunofluorescence
3.2.4. PON3 and Astrocyte–Microglia–Neuron “Triad” Double Immunofluorescence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Action Plan on the Public Health Response to Dementia 2017–2025; World Health Organization: Geneva, Switzerland, 2017; ISBN 978-92-4-151348-7.
- Metsaars, W.P.; Hauw, J.-J.; Van Welsem, M.E.; Duyckaerts, C. A Grading System of Alzheimer Disease Lesions in Neocortical Areas. Neurobiol. Aging 2003, 24, 563–572. [Google Scholar] [CrossRef]
- Rajasekhar, K.; Chakrabarti, M.; Govindaraju, T. Function and Toxicity of Amyloid Beta and Recent Therapeutic Interventions Targeting Amyloid Beta in Alzheimer’s Disease. Chem. Commun. 2015, 51, 13434–13450. [Google Scholar] [CrossRef]
- Hsiao, K.; Chapman, P.; Nilsen, S.; Eckman, C.; Harigaya, Y.; Younkin, S.; Yang, F.; Cole, G. Correlative Memory Deficits, Abeta Elevation, and Amyloid Plaques in Transgenic Mice. Science 1996, 274, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Pappolla, M.A.; Chyan, Y.J.; Omar, R.A.; Hsiao, K.; Perry, G.; Smith, M.A.; Bozner, P. Evidence of Oxidative Stress and in Vivo Neurotoxicity of Beta-Amyloid in a Transgenic Mouse Model of Alzheimer’s Disease: A Chronic Oxidative Paradigm for Testing Antioxidant Therapies in Vivo. Am. J. Pathol. 1998, 152, 871–877. [Google Scholar]
- Mackness, M.I. ’A’-Esterases. Enzymes Looking for a Role? Biochem. Pharmacol. 1989, 38, 385–390. [Google Scholar] [CrossRef]
- Hassett, C.; Richter, R.J.; Humbert, R.; Chapline, C.; Crabb, J.W.; Omiecinski, C.J.; Furlong, C.E. Characterization of CDNA Clones Encoding Rabbit and Human Serum Paraoxonase: The Mature Protein Retains Its Signal Sequence. Biochemistry 1991, 30, 10141–10149. [Google Scholar] [CrossRef]
- Reddy, S.T.; Wadleigh, D.J.; Grijalva, V.; Ng, C.; Hama, S.; Gangopadhyay, A.; Shih, D.M.; Lusis, A.J.; Navab, M.; Fogelman, A.M. Human Paraoxonase-3 Is an HDL-Associated Enzyme with Biological Activity Similar to Paraoxonase-1 Protein but Is Not Regulated by Oxidized Lipids. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 542–547. [Google Scholar] [CrossRef]
- Rothem, L.; Hartman, C.; Dahan, A.; Lachter, J.; Eliakim, R.; Shamir, R. Paraoxonases Are Associated with Intestinal Inflammatory Diseases and Intracellularly Localized to the Endoplasmic Reticulum. Free Radic. Biol. Med. 2007, 43, 730–739. [Google Scholar] [CrossRef]
- Schweikert, E.-M.; Devarajan, A.; Witte, I.; Wilgenbus, P.; Amort, J.; Förstermann, U.; Shabazian, A.; Grijalva, V.; Shih, D.M.; Farias-Eisner, R.; et al. PON3 Is Upregulated in Cancer Tissues and Protects against Mitochondrial Superoxide-Mediated Cell Death. Cell Death Differ. 2012, 19, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Deakin, S.; Leviev, I.; Gomaraschi, M.; Calabresi, L.; Franceschini, G.; James, R.W. Enzymatically Active Paraoxonase-1 Is Located at the External Membrane of Producing Cells and Released by a High Affinity, Saturable, Desorption Mechanism. J. Biol. Chem. 2002, 277, 4301–4308. [Google Scholar] [CrossRef] [PubMed]
- Efrat, M.; Aviram, M. Macrophage Paraoxonase 1 (PON1) Binding Sites. Biochem. Biophys. Res. Commun. 2008, 376, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Deakin, S.P.; Bioletto, S.; Bochaton-Piallat, M.-L.; James, R.W. HDL-Associated Paraoxonase-1 Can Redistribute to Cell Membranes and Influence Sensitivity to Oxidative Stress. Free Radic. Biol. Med. 2011, 50, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Mackness, B.; Hunt, R.; Durrington, P.N.; Mackness, M.I. Increased Immunolocalization of Paraoxonase, Clusterin, and Apolipoprotein A-I in the Human Artery Wall with the Progression of Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, L.; Hernández, A.F.; López-Caballero, J.J.; Gil, F.; Pla, A. Immunohistochemical Evidence for the Expression and Induction of Paraoxonase in Rat Liver, Kidney, Lung and Brain Tissue. Implications for Its Physiological Role. Chem. Biol. Interact. 2001, 137, 123–137. [Google Scholar] [CrossRef]
- Marsillach, J.; Mackness, B.; Mackness, M.; Riu, F.; Beltrán, R.; Joven, J.; Camps, J. Immunohistochemical Analysis of Paraoxonases-1, 2, and 3 Expression in Normal Mouse Tissues. Free Radic. Biol. Med. 2008, 45, 146–157. [Google Scholar] [CrossRef]
- Marsillach, J.; Camps, J.; Beltran-Debón, R.; Rull, A.; Aragones, G.; Maestre-Martínez, C.; Sabench, F.; Hernández, M.; Castillo, D.D.; Joven, J.; et al. Immunohistochemical Analysis of Paraoxonases-1 and 3 in Human Atheromatous Plaques. Eur. J. Clin. Investig. 2011, 41, 308–314. [Google Scholar] [CrossRef]
- Furlong, C.E.; Richter, R.J.; Li, W.-F.; Brophy, V.H.; Carlson, C.; Rieder, M.; Nickerson, D.; Costa, L.G.; Ranchalis, J.; Lusis, A.J.; et al. The Functional Consequences of Polymorphisms in the Human PON1 Gene. In The Paraoxonases: Their Role in Disease Development and Xenobiotic Metabolism; Proteins And Cell Regulation; Mackness, B., Mackness, M., Aviram, M., Paragh, G., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 267–281. ISBN 978-1-4020-6561-3. [Google Scholar]
- Furlong, C.E.; Marsillach, J.; Jarvik, G.P.; Costa, L.G. Paraoxonases-1, -2 and -3: What Are Their Functions? Chem. Biol. Interact 2016, 259, 51–62. [Google Scholar] [CrossRef]
- Khersonsky, O.; Tawfik, D.S. Structure-Reactivity Studies of Serum Paraoxonase PON1 Suggest That Its Native Activity Is Lactonase. Biochemistry 2005, 44, 6371–6382. [Google Scholar] [CrossRef]
- Draganov, D.I.; Teiber, J.F.; Speelman, A.; Osawa, Y.; Sunahara, R.; La Du, B.N. Human Paraoxonases (PON1, PON2, and PON3) Are Lactonases with Overlapping and Distinct Substrate Specificities. J. Lipid Res. 2005, 46, 1239–1247. [Google Scholar] [CrossRef]
- Aviram, M.; Rosenblat, M. Paraoxonases 1, 2, and 3, Oxidative Stress, and Macrophage Foam Cell Formation during Atherosclerosis Development. Free Radic. Biol. Med. 2004, 37, 1304–1316. [Google Scholar] [CrossRef]
- Ng Carey, J.; Bourquard, N.; Hama, S.Y.; Shih, D.; Grijalva Victor, R.; Navab, M.; Fogelman Alan, M.; Reddy Srinivasa, T. Adenovirus-Mediated Expression of Human Paraoxonase 3 Protects Against the Progression of Atherosclerosis in Apolipoprotein E–Deficient Mice. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1368–1374. [Google Scholar] [CrossRef]
- Liu, Y.; Mackness, B.; Mackness, M. Comparison of the Ability of Paraoxonases 1 and 3 to Attenuate the in Vitro Oxidation of Low-Density Lipoprotein and Reduce Macrophage Oxidative Stress. Free Radic. Biol. Med. 2008, 45, 743–748. [Google Scholar] [CrossRef]
- Zhang, C.; Peng, W.; Wang, M.; Zhu, J.; Zang, Y.; Shi, W.; Zhang, J.; Qin, J. Studies on Protective Effects of Human Paraoxonases 1 and 3 on Atherosclerosis in Apolipoprotein E Knockout Mice. Gene Ther. 2010, 17, 626–633. [Google Scholar] [CrossRef]
- Camps, J.; Marsillach, J.; Joven, J. The Paraoxonases: Role in Human Diseases and Methodological Difficulties in Measurement. Crit. Rev. Clin. Lab. Sci. 2009, 46, 83–106. [Google Scholar] [CrossRef]
- Cervellati, C.; Valacchi, G.; Tisato, V.; Zuliani, G.; Marsillach, J. Evaluating the Link between Paraoxonase-1 Levels and Alzheimer’s Disease Development. Minerva Med. 2019, 110, 238–250. [Google Scholar] [CrossRef]
- Bennett, S.; Grant, M.M.; Aldred, S. Oxidative Stress in Vascular Dementia and Alzheimer’s Disease: A Common Pathology. J. Alzheimers Dis. 2009, 17, 245–257. [Google Scholar] [CrossRef]
- Cheignon, C.; Tomas, M.; Bonnefont-Rousselot, D.; Faller, P.; Hureau, C.; Collin, F. Oxidative Stress and the Amyloid Beta Peptide in Alzheimer’s Disease. Redox Biol. 2018, 14, 450–464. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Boyd-Kimball, D. Oxidative Stress, Amyloid-β Peptide, and Altered Key Molecular Pathways in the Pathogenesis and Progression of Alzheimer’s Disease. J. Alzheimers Dis. 2018, 62, 1345–1367. [Google Scholar] [CrossRef] [PubMed]
- Kontush, A.; Atwood, C.S. Amyloid-Beta: Phylogenesis of a Chameleon. Brain Res. Brain Res. Rev. 2004, 46, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Bassett, C.N.; Montine, T.J. Lipoproteins and Lipid Peroxidation in Alzheimer’s Disease. J. Nutr. Health Aging 2003, 7, 24–29. [Google Scholar] [PubMed]
- Markesbery, W.R.; Carney, J.M. Oxidative Alterations in Alzheimer’s Disease. Brain Pathol. 1999, 9, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Lovell, M.A.; Ehmann, W.D.; Butler, S.M.; Markesbery, W.R. Elevated Thiobarbituric Acid-Reactive Substances and Antioxidant Enzyme Activity in the Brain in Alzheimer’s Disease. Neurology 1995, 45, 1594–1601. [Google Scholar] [CrossRef] [PubMed]
- Primo-Parmo, S.L.; Sorenson, R.C.; Teiber, J.; Du, B.N.L. The Human Serum Paraoxonase/Arylesterase Gene (PON1) Is One Member of a Multigene Family. Genomics 1996, 33, 498–507. [Google Scholar] [CrossRef]
- Mackness, B.; Beltran-Debon, R.; Aragones, G.; Joven, J.; Camps, J.; Mackness, M. Human Tissue Distribution of Paraoxonases 1 and 2 MRNA. IUBMB Life 2010, 62, 480–482. [Google Scholar] [CrossRef] [PubMed]
- Shih, D.M.; Xia, Y.-R.; Yu, J.M.; Lusis, A.J. Temporal and Tissue-Specific Patterns of Pon3 Expression in Mouse: In Situ Hybridization Analysis. Adv. Exp. Med. Biol. 2010, 660, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.J.; Wadleigh, D.J.; Gangopadhyay, A.; Hama, S.; Grijalva, V.R.; Navab, M.; Fogelman, A.M.; Reddy, S.T. Paraoxonase-2 Is a Ubiquitously Expressed Protein with Antioxidant Properties and Is Capable of Preventing Cell-Mediated Oxidative Modification of Low Density Lipoprotein. J. Biol. Chem. 2001, 276, 44444–44449. [Google Scholar] [CrossRef]
- Wills, A.-M.; Landers, J.E.; Zhang, H.; Richter, R.J.; Caraganis, A.J.; Cudkowicz, M.E.; Furlong, C.E.; Brown, R.H. Paraoxonase 1 (PON1) Organophosphate Hydrolysis Is Not Reduced in ALS. Neurology 2008, 70, 929–934. [Google Scholar] [CrossRef]
- Castellazzi, M.; Trentini, A.; Romani, A.; Valacchi, G.; Bellini, T.; Bonaccorsi, G.; Fainardi, E.; Cavicchio, C.; Passaro, A.; Zuliani, G.; et al. Decreased Arylesterase Activity of Paraoxonase-1 (PON-1) Might Be a Common Denominator of Neuroinflammatory and Neurodegenerative Diseases. Int. J. Biochem. Cell Biol. 2016, 81, 356–363. [Google Scholar] [CrossRef]
- Romani, A.; Trentini, A.; Van der Flier, W.M.; Bellini, T.; Zuliani, G.; Cervellati, C.; Teunissen, C.E. Arylesterase Activity of Paraoxonase-1 in Serum and Cerebrospinal Fluid of Patients with Alzheimer’s Disease and Vascular Dementia. Antioxidants 2020, 9, 456. [Google Scholar] [CrossRef]
- Balazs, Z.; Panzenboeck, U.; Hammer, A.; Sovic, A.; Quehenberger, O.; Malle, E.; Sattler, W. Uptake and Transport of High-Density Lipoprotein (HDL) and HDL-Associated Alpha-Tocopherol by an in Vitro Blood-Brain Barrier Model. J. Neurochem. 2004, 89, 939–950. [Google Scholar] [CrossRef]
- Fung, K.Y.; Wang, C.; Nyegaard, S.; Heit, B.; Fairn, G.D.; Lee, W.L. SR-BI Mediated Transcytosis of HDL in Brain Microvascular Endothelial Cells Is Independent of Caveolin, Clathrin, and PDZK1. Front Physiol. 2017, 8, 841. [Google Scholar] [CrossRef]
- Blain, J.-F.; Poirier, J. Cholesterol Homeostasis and the Pathophysiology of Alzheimer’s Disease. Expert Rev. Neurother. 2004, 4, 823–829. [Google Scholar] [CrossRef]
- Lassmann, H.; Van Horssen, J. Oxidative Stress and Its Impact on Neurons and Glia in Multiple Sclerosis Lesions. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2016, 1862, 506–510. [Google Scholar] [CrossRef]
- Yamada, T.; Tsuboi, Y.; Takahashi, M. Interrelationship between Beta-Amyloid Deposition and Complement-Activated Oligodendroglia. Dement. Geriatr. Cogn. Disord. 1997, 8, 267–272. [Google Scholar] [CrossRef]
- Luca, A.; Calandra, C.; Luca, M. Molecular Bases of Alzheimer’s Disease and Neurodegeneration: The Role of Neuroglia. Aging Dis. 2018, 9, 1134–1152. [Google Scholar] [CrossRef] [PubMed]
- Juurlink, B.H.J.; Thorburne, S.K.; Hertz, L. Peroxide-Scavenging Deficit Underlies Oligodendrocyte Susceptibility to Oxidative Stress. Glia 1998, 22, 371–378. [Google Scholar] [CrossRef]
- Li, Q.; Barres, B.A. Microglia and Macrophages in Brain Homeostasis and Disease. Nat. Rev. Immunol. 2018, 18, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Mathys, H.; Davila-Velderrain, J.; Peng, Z.; Gao, F.; Mohammadi, S.; Young, J.Z.; Menon, M.; He, L.; Abdurrob, F.; Jiang, X.; et al. Single-Cell Transcriptomic Analysis of Alzheimer’s Disease. Nature 2019, 570, 332–337. [Google Scholar] [CrossRef]
- Loving, B.A.; Bruce, K.D. Lipid and Lipoprotein Metabolism in Microglia. Front Physiol. 2020, 11. [Google Scholar] [CrossRef]
- Yuan, P.; Condello, C.; Keene, C.D.; Wang, Y.; Bird, T.D.; Paul, S.M.; Luo, W.; Colonna, M.; Baddeley, D.; Grutzendler, J. TREM2 Haplodeficiency in Mice and Humans Impairs the Microglia Barrier Function Leading to Decreased Amyloid Compaction and Severe Axonal Dystrophy. Neuron 2016, 90, 724–739. [Google Scholar] [CrossRef]
- Su, B.; Wang, X.; Nunomura, A.; Moreira, P.I.; Lee, H.; Perry, G.; Smith, M.A.; Zhu, X. Oxidative Stress Signaling in Alzheimer’s Disease. Curr. Alzheimer Res. 2008, 5, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Simpson, D.S.A.; Oliver, P.L. ROS Generation in Microglia: Understanding Oxidative Stress and Inflammation in Neurodegenerative Disease. Antioxidants 2020, 9, 743. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar, J.G.; Marsillach, J.; Reverte, I.; Mackness, B.; Mackness, M.; Joven, J.; Camps, J.; Colomina, M.T. Paraoxonase-1 and -3 Protein Expression in the Brain of the Tg2576 Mouse Model of Alzheimer’s Disease. Antioxidants 2021, 10, 339. https://doi.org/10.3390/antiox10030339
Salazar JG, Marsillach J, Reverte I, Mackness B, Mackness M, Joven J, Camps J, Colomina MT. Paraoxonase-1 and -3 Protein Expression in the Brain of the Tg2576 Mouse Model of Alzheimer’s Disease. Antioxidants. 2021; 10(3):339. https://doi.org/10.3390/antiox10030339
Chicago/Turabian StyleSalazar, Jose Gregorio, Judit Marsillach, Ingrid Reverte, Bharti Mackness, Michael Mackness, Jorge Joven, Jordi Camps, and Maria Teresa Colomina. 2021. "Paraoxonase-1 and -3 Protein Expression in the Brain of the Tg2576 Mouse Model of Alzheimer’s Disease" Antioxidants 10, no. 3: 339. https://doi.org/10.3390/antiox10030339
APA StyleSalazar, J. G., Marsillach, J., Reverte, I., Mackness, B., Mackness, M., Joven, J., Camps, J., & Colomina, M. T. (2021). Paraoxonase-1 and -3 Protein Expression in the Brain of the Tg2576 Mouse Model of Alzheimer’s Disease. Antioxidants, 10(3), 339. https://doi.org/10.3390/antiox10030339