
The Impact of N-Acetyl Cysteine and Coenzyme Q10 Supplementation on Skeletal Muscle Antioxidants and Proteome in Fit Thoroughbred Horses



Abstract
:1. Introduction
2. Materials and Methods
3. Analyses
3.1. Blood Samples
3.2. Muscle Biochemistry
3.3. Statistical Analysis
3.4. Muscle Proteomics
4. Results
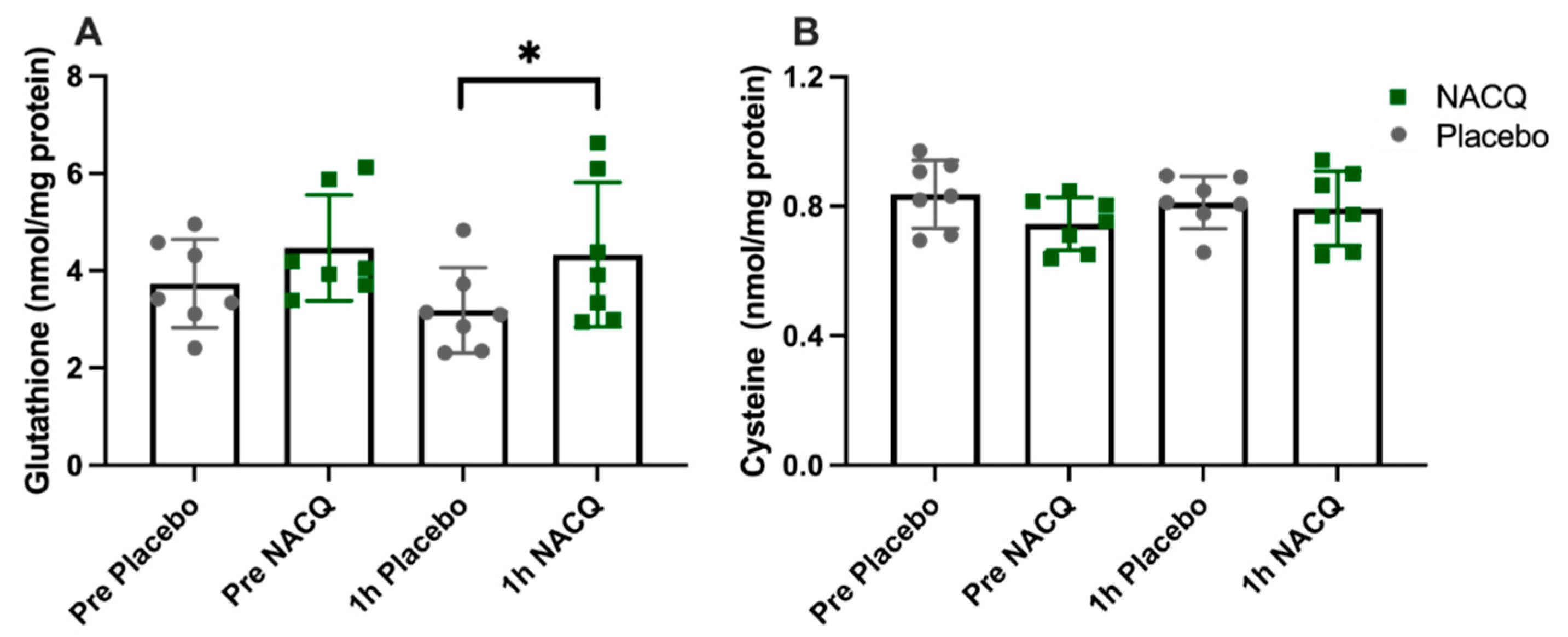
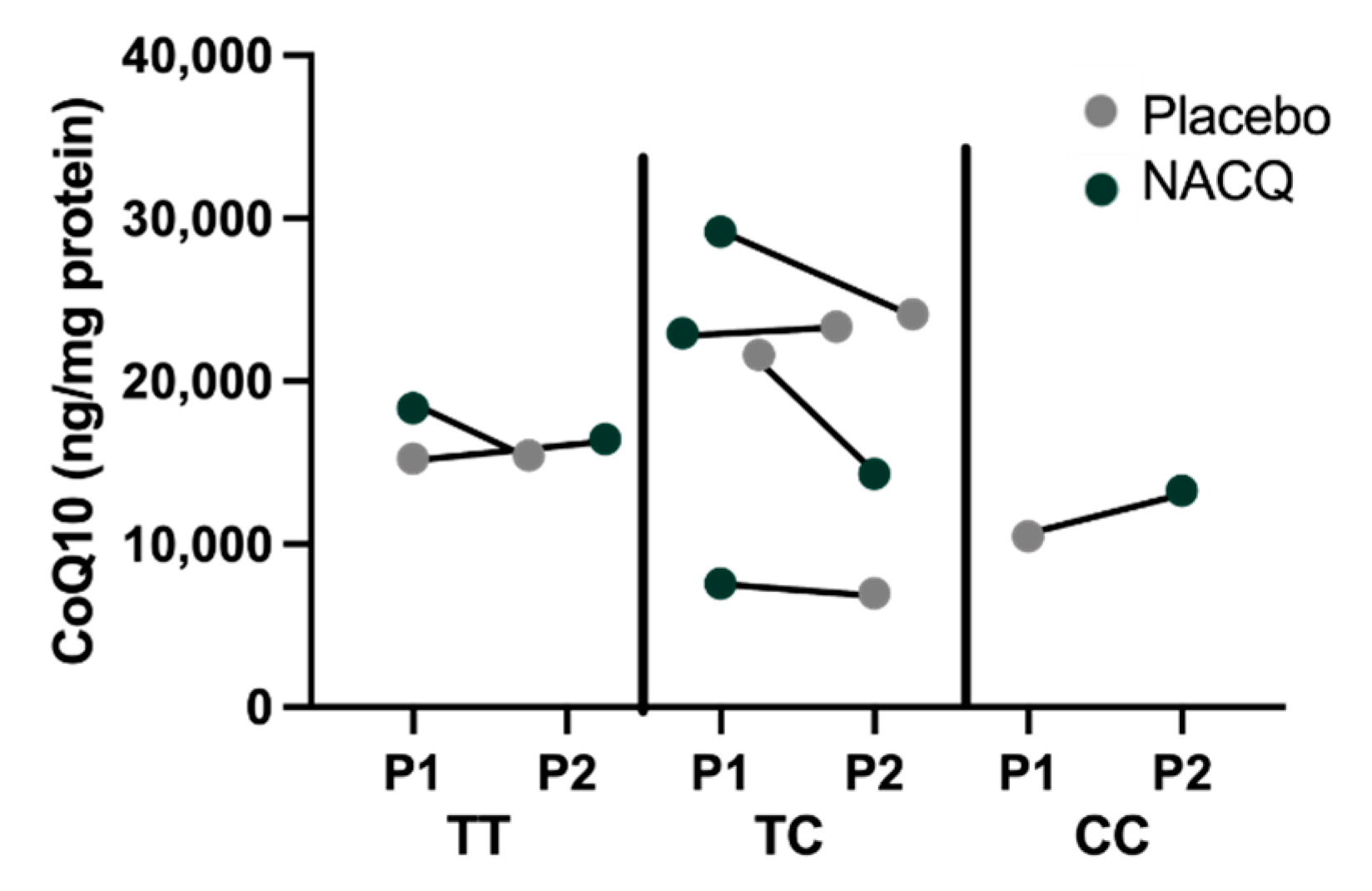
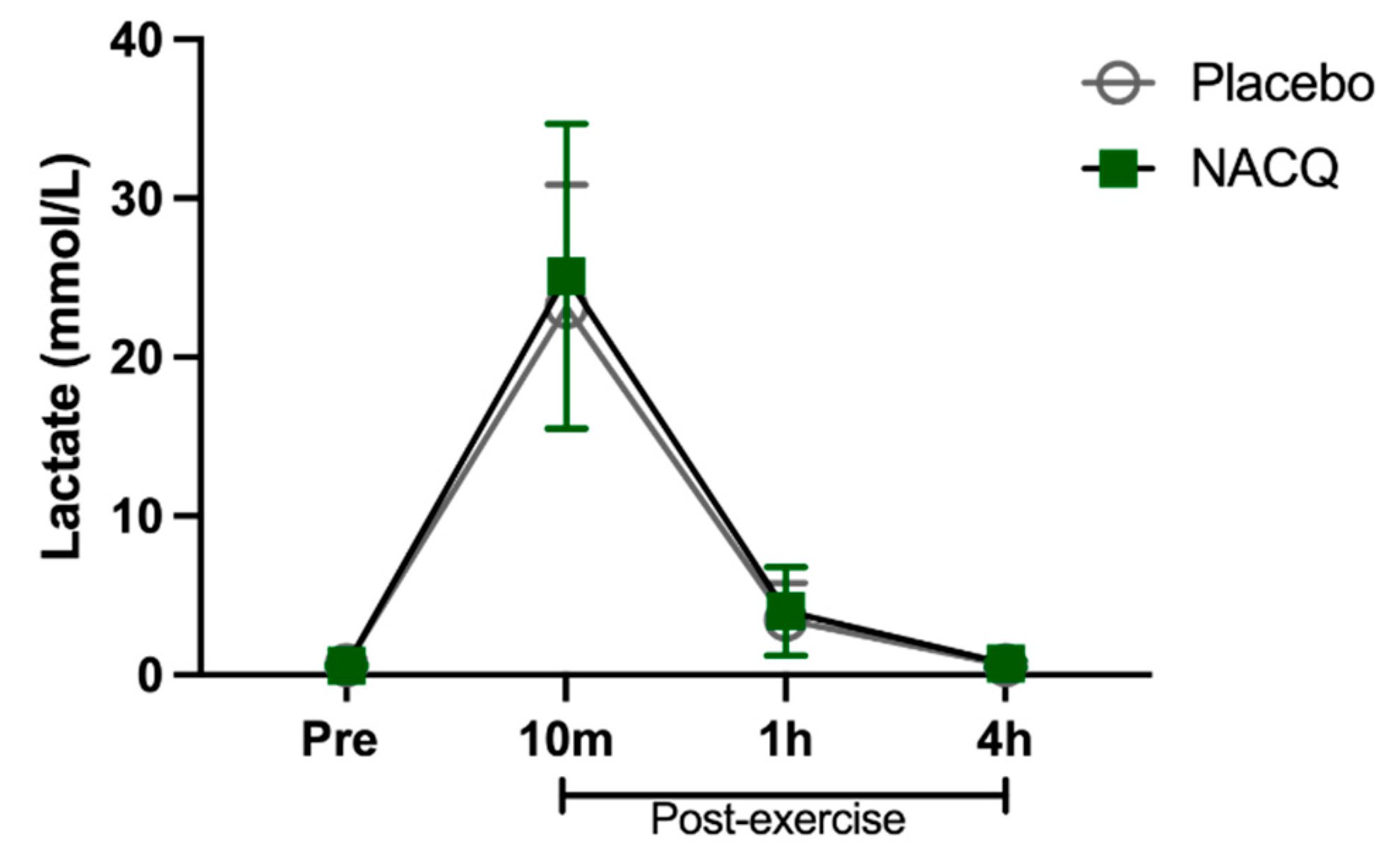
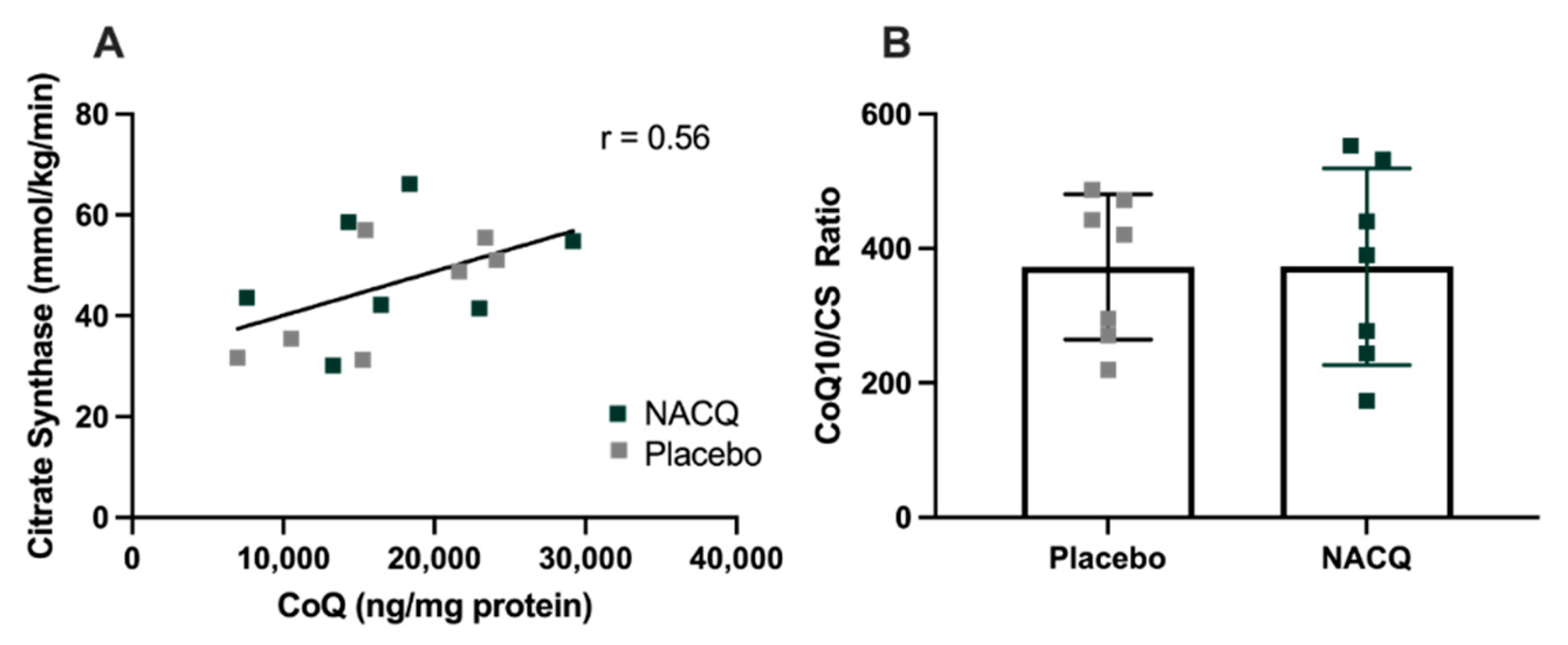
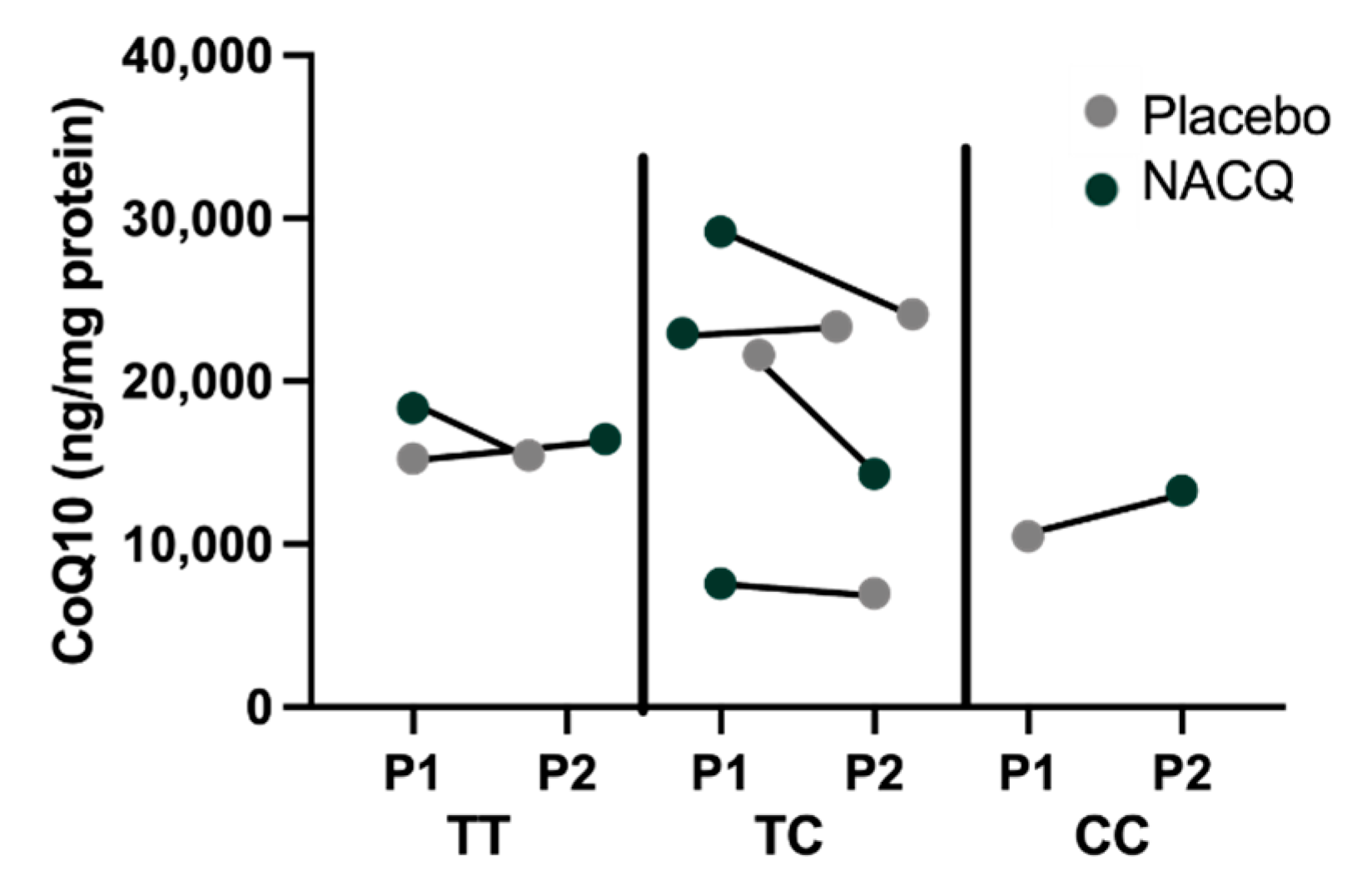
4.1. Muscle Biochemistry
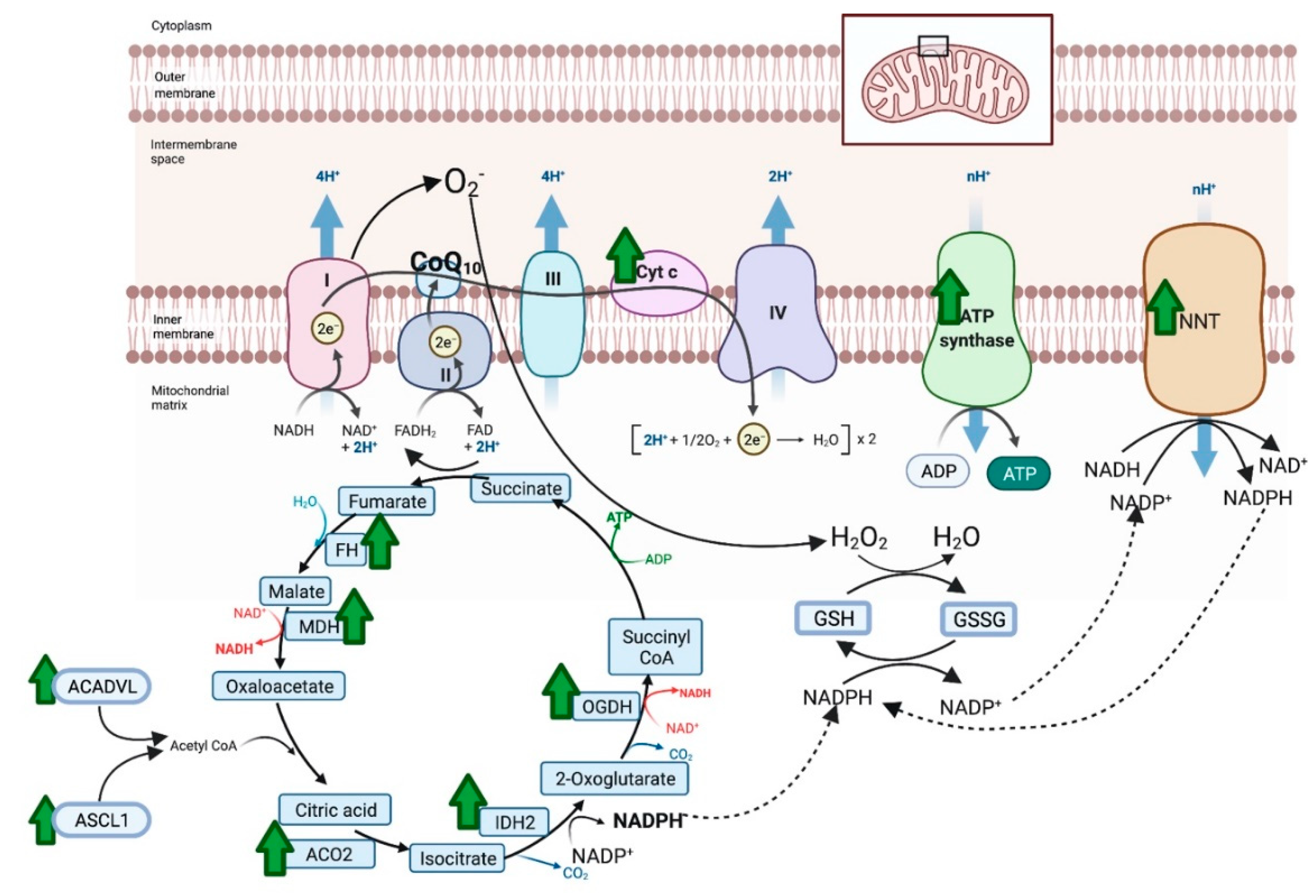
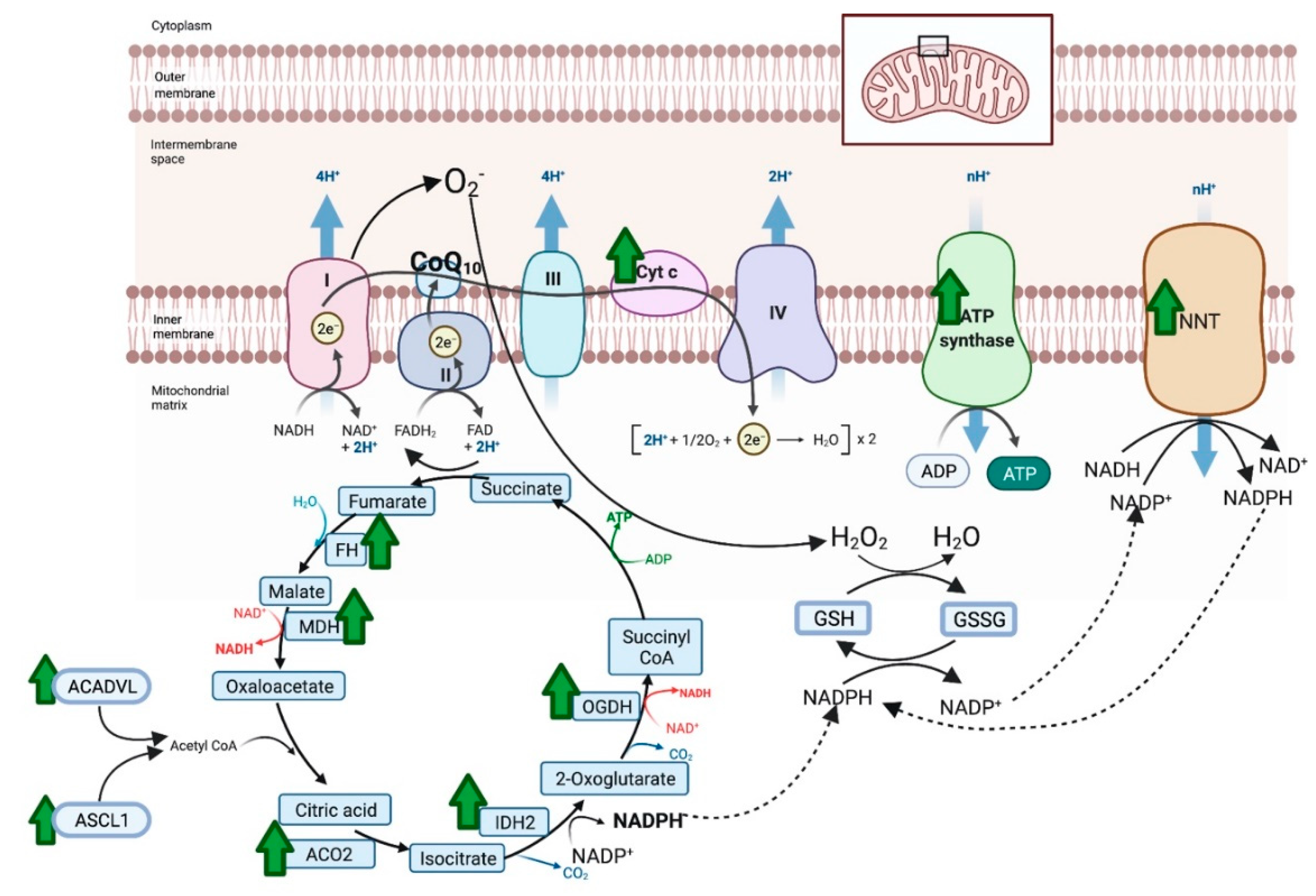
4.2. Proteomics
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Essen-Gustavsson, E. Activity and inactivity related muscle adaptations in the animal kingdom. Biochem. Exerc. 1986, 6, 435–454. [Google Scholar]
- Shadel, G.S.; Horvath, T.L. Mitochondrial ROS signaling in organismal homeostasis. Cell 2015, 163, 560–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribas, V.; Garcia-Ruiz, C.; Fernandez-Checa, J.C. Glutathione and mitochondria. Front. Pharm. 2014, 5, 151. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, L.F.; Reid, M.B. Muscle-derived ROS and thiol regulation in muscle fatigue. J. Appl. Physiol. 2008, 104, 853–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.H.; Stipanuk, M.H. Cysteine regulates expression of cysteine dioxygenase and gamma-glutamylcysteine synthetase in cultured rat hepatocytes. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E804–E815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Long, Y.C. Crosstalk between cystine and glutathione is critical for the regulation of amino acid signaling pathways and ferroptosis. Sci. Rep. 2016, 6, 30033. [Google Scholar] [CrossRef]
- Droge, W. Oxidative stress and ageing: Is ageing a cysteine deficiency syndrome? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 2355–2372. [Google Scholar] [CrossRef] [PubMed]
- Atkuri, K.R.; Mantovani, J.J.; Herzenberg, L.A.; Herzenberg, L.A. N-Acetylcysteine--a safe antidote for cysteine/glutathione deficiency. Curr. Opin. Pharmacol. 2007, 7, 355–359. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Tamba, M.; Ishii, T.; Bannai, S. Cloning and expression of a plasma membrane cystine/glutamate exchange transporter composed of two distinct proteins. J. Biol. Chem. 1999, 274, 11455–11458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witte, T.S.; Melkus, E.; Walter, I.; Senge, B.; Schwab, S.; Aurich, C.; Heuwieser, W. Effects of oral treatment with N-acetylcysteine on the viscosity of intrauterine mucus and endometrial function in estrous mares. Theriogenology 2012, 78, 1199–1208. [Google Scholar] [CrossRef]
- Roneus, B.; Hakkarainen, J. Vitamin E in serum and skeletal muscle tissue and blood glutathione peroxidase activity from horses with the azoturia-tying-up syndrome. Acta Vet. Scand. 1985, 26, 425–427. [Google Scholar] [CrossRef]
- Roneus, B.O.; Hakkarainen, R.V.; Lindholm, C.A.; Tyopponen, J.T. Vitamin E requirements of adult Standardbred horses evaluated by tissue depletion and repletion. Equine Vet. J. 1986, 18, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.C.; Valberg, S.J.; Hogg, M.; Finno, C.J. Effects of feeding two RRR-alpha-tocopherol formulations on serum, cerebrospinal fluid and muscle alpha-tocopherol concentrations in horses with subclinical vitamin E deficiency. Equine Vet. J. 2017, 49, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Maraver, J.; Cordero, M.D.; Oropesa-Avila, M.; Vega, A.F.; de la Mata, M.; Pavon, A.D.; Alcocer-Gomez, E.; Calero, C.P.; Paz, M.V.; Alanis, M.; et al. Clinical applications of coenzyme Q10. Front. Biosci. 2014, 19, 619–633. [Google Scholar] [CrossRef] [Green Version]
- Sinatra, S.T.; Jankowitz, S.N.; Chopra, R.K.; Bhagavan, H.N. Plasma Coenzyme Q10 and Tocopherols in Thoroughbred Race Horses: Effect of Coenzyme Q10 Supplementation and Exercise. J. Equine Vet. Sci. 2014, 34, 265–269. [Google Scholar] [CrossRef]
- Ruiz, A.J.; Tibary, A.; Heaton, R.A.; Hargreaves, I.P.; Leadon, D.P.; Bayly, W.M. Effects of Feeding Coenzyme Q10-Ubiquinol on Plasma Coenzyme Q10 Concentrations and Semen Quality in Stallions. J. Equine Vet. Sci. 2021, 96, 103303. [Google Scholar] [CrossRef] [PubMed]
- Thueson, E.; Leadon, D.P.; Heaton, R.; Hargreaves, I.P.; Bayly, W.M. Effect of daily supplementation with ubiquinol on muscle coenzyme Q10 concentrations in Thoroughbred racehorses. Comp. Exerc. Physiol. 2019, 15, 219–226. [Google Scholar] [CrossRef]
- Poole, D.C. Current concepts of oxygen transport during exercise. Equine Comp. Exerc. Physiol. 2003, 1, 5–22. [Google Scholar] [CrossRef] [Green Version]
- Hoppeler, H.; Weibel, E.R. Limits for oxygen and substrate transport in mammals. J. Exp. Biol. 1998, 201, 1051–1064. [Google Scholar] [CrossRef] [PubMed]
- Ceusters, J.D.; Mouithys-Mickalad, A.A.; de la Rebiere de Pouyade, G.; Franck, T.J.; Votion, D.M.; Deby-Dupont, G.P.; Serteyn, D.A. Assessment of reactive oxygen species production in cultured equine skeletal myoblasts in response to conditions of anoxia followed by reoxygenation with or without exposure to peroxidases. Am. J. Vet. Res. 2012, 73, 426–434. [Google Scholar] [CrossRef]
- Bookbinder, L.; Finno, C.J.; Firshman, A.M.; Katzman, S.A.; Burns, E.; Peterson, J.; Dahlgren, A.; Ming-Whitfield, B.; Glessner, S.; Borer-Matsui, A.; et al. Impact of alpha-tocopherol deficiency and supplementation on sacrocaudalis and gluteal muscle fiber histopathology and morphology in horses. J. Vet. Intern. Med. 2019, 33, 2770–2779. [Google Scholar] [CrossRef] [PubMed]
- Valberg, S.J.; McKenzie, E.C.; Eyrich, L.V.; Shivers, J.; Barnes, N.E.; Finno, C.J. Suspected myofibrillar myopathy in Arabian horses with a history of exertional rhabdomyolysis. Equine Vet. J. 2016, 48, 548–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindholm, A.; Piehl, K. Fibre composition, enzyme activity and concentrations of metabolites and electrolytes in muscles of standardbred horses. Acta Vet. Scand. 1974, 15, 287–309. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.L.; Valberg, S.J.; Mickelson, J.R.; McCue, M.E. Haplotype diversity in the equine myostatin gene with focus on variants associated with race distance propensity and muscle fiber type proportions. Anim. Genet. 2014, 45, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Hill, E.W.; McGivney, B.A.; Gu, J.; Whiston, R.; Machugh, D.E. A genome-wide SNP-association study confirms a sequence variant (g.66493737C>T) in the equine myostatin (MSTN) gene as the most powerful predictor of optimum racing distance for Thoroughbred racehorses. BMC Genom. 2010, 11, 552. [Google Scholar] [CrossRef] [Green Version]
- Pandey, R.; Riley, C.L.; Mills, E.M.; Tiziani, S. Highly sensitive and selective determination of redox states of coenzymes Q9 and Q10 in mice tissues: Application of orbitrap mass spectrometry. Anal. Chim. Acta 2018, 1011, 68–76. [Google Scholar] [CrossRef]
- Essen-Gustavsson, B.; Henriksson, J. Enzyme levels in pools of microdissected human muscle fibres of identified type. Adaptive response to exercise. Acta Physiol. Scand. 1984, 120, 505–515. [Google Scholar] [CrossRef]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef] [PubMed]
- Shadforth, I.P.; Dunkley, T.P.; Lilley, K.S.; Bessant, C. i-Tracker: For quantitative proteomics using iTRAQ. BMC Genom. 2005, 6, 145. [Google Scholar] [CrossRef] [Green Version]
- Oberg, A.L.; Mahoney, D.W.; Eckel-Passow, J.E.; Malone, C.J.; Wolfinger, R.D.; Hill, E.G.; Cooper, L.T.; Onuma, O.K.; Spiro, C.; Therneau, T.M.; et al. Statistical analysis of relative labeled mass spectrometry data from complex samples using ANOVA. J. Proteome Res. 2008, 7, 225–233. [Google Scholar] [CrossRef]
- McGivney, B.A.; Browne, J.A.; Fonseca, R.G.; Katz, L.M.; Machugh, D.E.; Whiston, R.; Hill, E.W. MSTN genotypes in Thoroughbred horses influence skeletal muscle gene expression and racetrack performance. Anim. Genet. 2012, 43, 810–812. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.L.; Hammarqvist, F.; Andersson, K.; Wernerman, J. Skeletal muscle glutathione after surgical trauma. Ann. Surg. 1996, 223, 420–427. [Google Scholar] [CrossRef]
- Hammarqvist, F.; Luo, J.L.; Cotgreave, I.A.; Andersson, K.; Wernerman, J. Skeletal muscle glutathione is depleted in critically ill patients. Crit. Care Med. 1997, 25, 78–84. [Google Scholar] [CrossRef]
- Hammarqvist, F.; Andersson, K.; Luo, J.L.; Wernerman, J. Free amino acid and glutathione concentrations in muscle during short-term starvation and refeeding. Clin. Nutr. 2005, 24, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Michailidis, Y.; Karagounis, L.G.; Terzis, G.; Jamurtas, A.Z.; Spengos, K.; Tsoukas, D.; Chatzinikolaou, A.; Mandalidis, D.; Stefanetti, R.J.; Papassotiriou, I.; et al. Thiol-based antioxidant supplementation alters human skeletal muscle signaling and attenuates its inflammatory response and recovery after intense eccentric exercise. Am. J. Clin. Nutr. 2013, 98, 233–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martensson, J.; Meister, A. Mitochondrial damage in muscle occurs after marked depletion of glutathione and is prevented by giving glutathione monoester. Proc. Natl. Acad. Sci. USA 1989, 86, 471–475. [Google Scholar] [CrossRef] [Green Version]
- Ji, L.L.; Fu, R.; Mitchell, E.W. Glutathione and antioxidant enzymes in skeletal muscle: Effects of fiber type and exercise intensity. J. Appl. Physiol. 1992, 73, 1854–1859. [Google Scholar] [CrossRef] [PubMed]
- Dam, A.D.; Mitchell, A.S.; Rush, J.W.; Quadrilatero, J. Elevated skeletal muscle apoptotic signaling following glutathione depletion. Apoptosis 2012, 17, 48–60. [Google Scholar] [CrossRef]
- Morin, G.; Guiraut, C.; Perez Marcogliese, M.; Mohamed, I.; Lavoie, J.C. Glutathione Supplementation of Parenteral Nutrition Prevents Oxidative Stress and Sustains Protein Synthesis in Guinea Pig Model. Nutrients 2019, 11, 2063. [Google Scholar] [CrossRef] [Green Version]
- Marin, E.; Kretzschmar, M.; Arokoski, J.; Hanninen, O.; Klinger, W. Enzymes of glutathione synthesis in dog skeletal muscles and their response to training. Acta Physiol. Scand. 1993, 147, 369–373. [Google Scholar] [CrossRef]
- Staron, R.S.; Hagerman, F.C.; Hikida, R.S.; Murray, T.F.; Hostler, D.P.; Crill, M.T.; Ragg, K.E.; Toma, K. Fiber type composition of the vastus lateralis muscle of young men and women. J. Histochem. Cytochem. 2000, 48, 623–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldrich, K.; Velez-Irizarry, D.; Fenger, C.; Schott, M.; Valberg, S.J. Pathways of calcium regulation, electron transport, and mitochondrial protein translation are molecular signatures of susceptibility to recurrent exertional rhabdomyolysis in Thoroughbred racehorses. PLoS ONE 2021, 16, e0244556. [Google Scholar] [CrossRef]
- Radtke, K.K.; Coles, L.D.; Mishra, U.; Orchard, P.J.; Holmay, M.; Cloyd, J.C. Interaction of N-acetylcysteine and cysteine in human plasma. J. Pharm. Sci. 2012, 101, 4653–4659. [Google Scholar] [CrossRef] [PubMed]
- Atalay, M.; Seene, T.; Hanninen, O.; Sen, C.K. Skeletal muscle and heart antioxidant defences in response to sprint training. Acta Physiol. Scand. 1996, 158, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Medved, I.; Brown, M.J.; Bjorksten, A.R.; Murphy, K.T.; Petersen, A.C.; Sostaric, S.; Gong, X.; McKenna, M.J. N-acetylcysteine enhances muscle cysteine and glutathione availability and attenuates fatigue during prolonged exercise in endurance-trained individuals. J. Appl. Physiol. 2004, 97, 1477–1485. [Google Scholar] [CrossRef]
- Schubert, H.L.; Blumenthal, R.M.; Cheng, X. Many paths to methyltransfer: A chronicle of convergence. Trends Biochem. Sci. 2003, 28, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Schiaffino, S.; Reggiani, C.; Kostrominova, T.Y.; Mann, M.; Murgia, M. Mitochondrial specialization revealed by single muscle fiber proteomics: Focus on the Krebs cycle. Scand. J. Med. Sci. Sports 2015, 25 (Suppl. S4), 41–48. [Google Scholar] [CrossRef] [Green Version]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef]
- Cooke, M.; Iosia, M.; Buford, T.; Shelmadine, B.; Hudson, G.; Kerksick, C.; Rasmussen, C.; Greenwood, M.; Leutholtz, B.; Willoughby, D.; et al. Effects of acute and 14-day coenzyme Q10 supplementation on exercise performance in both trained and untrained individuals. J. Int. Soc. Sports Nutr. 2008, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Zhang, Y.; Davie, A.; Marshall-Gradisnik, S.; Hu, H.; Wang, J.; Brushett, D. Muscle and plasma coenzyme Q10 concentration, aerobic power and exercise economy of healthy men in response to four weeks of supplementation. J. Sports Med. Phys. Fit. 2005, 45, 337–346. [Google Scholar]
- Montero, R.; Sanchez-Alcazar, J.A.; Briones, P.; Hernandez, A.R.; Cordero, M.D.; Trevisson, E.; Salviati, L.; Pineda, M.; Garcia-Cazorla, A.; Navas, P.; et al. Analysis of coenzyme Q10 in muscle and fibroblasts for the diagnosis of CoQ10 deficiency syndromes. Clin. Biochem. 2008, 41, 697–700. [Google Scholar] [CrossRef] [PubMed]
- Sinatra, S.T.; Chopra, R.K.; Jankowitz, S.; Horohov, D.W.; Bhagavan, H.N. Coenzyme Q10 in Equine Serum: Response to Supplementation. J. Equine Vet. Sci. 2013, 33, 71–73. [Google Scholar] [CrossRef]
- Svete, A.N.; Vovk, T.; Topolovec, M.B.; Kruljc, P. Effects of Vitamin E and Coqnzyme Q(10) Supplementation on Oxidative Stress Parameters in Untrained Leisure Horses Subjected to Acute Moderate Exercise. Antioxidants 2021, 10, 908. [Google Scholar] [CrossRef]
- Rooney, M.F.; Porter, R.K.; Katz, L.M.; Hill, E.W. Skeletal muscle mitochondrial bioenergetics and associations with myostatin genotypes in the Thoroughbred horse. PLoS ONE 2017, 12, e0186247. [Google Scholar] [CrossRef] [PubMed]
- Hoopeler, H.; Jones, J.H.; Lindstedt, S.L.; Claasen, H.; Longworth, K.E.; Taylor, C.R.; Straub, R.; Lindholm, A. Relating maximal oxygen consumption to skeletal muscle mitochodria in horses. Equine Exerc. Physiol. 1987, 2, 278–289. [Google Scholar]
- Fittipaldi, S.; Mercatelli, N.; Dimauro, I.; Jackson, M.J.; Paronetto, M.P.; Caporossi, D. Alpha B-crystallin induction in skeletal muscle cells under redox imbalance is mediated by a JNK-dependent regulatory mechanism. Free Radic. Biol. Med. 2015, 86, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Forrest, K.M.; Al-Sarraj, S.; Sewry, C.; Buk, S.; Tan, S.V.; Pitt, M.; Durward, A.; McDougall, M.; Irving, M.; Hanna, M.G.; et al. Infantile onset myofibrillar myopathy due to recessive CRYAB mutations. Neuromuscul. Disord. 2011, 21, 37–40. [Google Scholar] [CrossRef]
- Williams, Z.J.; Velez-Irizarry, D.; Gardner, K.; Valberg, S.J. Integrated proteomic and transcriptomic profiling identifies aberrant gene and protein expression in the sarcomere, mitochondrial complex I, and the extracellular matrix in Warmblood horses with myofibrillar myopathy. BMC Genom. 2021, 22, 438. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Amount |
|---|---|
| Crude Protein (Min) | 12.0% |
| Lysine (Min) | 0.7% |
| Crude Fat (Min) | 8.0% |
| Crude Fiber (Max) | 9.0% |
| Calcium (Min) | 0.6% |
| Calcium (Max) | 0.7% |
| Phosphorus (Min) | 0.5% |
| Selenium (Min) | 0.5 ppm |
| Zinc (Min) | 145 ppm |
| Vitamin A (Min) | 11,000 IU/kg |
| Vitamin D (Min) | 1100 IU/kg |
| Vitamin E (Min) | 211 IU/kg |
| GGT (U/L) | CK (U/L) | |
|---|---|---|
| Mean ± Sd | Mean ± Sd | |
| Placebo Pre | 18.7 ± 6.0 a | 190.0 ± 44.3 a |
| Placebo 4 h Post | 25.0 ± 8.0 b | 446.1 ± 171.2 b |
| NACQ Pre | 20.0 ± 7.2 a | 278.4 ± 151.4 a |
| NACQ 4 h Post | 22.6 ± 5.7 b | 387.4 ± 146.2 b |
| Supplement | Rest | 1 h Post-Exercise | p Value | |
|---|---|---|---|---|
| ROS | NACQ | 16.51 ± 6.14 a | 14.68 ± 5.60 a | |
| (μM H2O2 equivalence) | Placebo | 13.81 ± 4.83 a | 17.55 ± 5.00 a | 0.4779 |
| CoQ10 | NACQ | 7090 ± 2898 a | 7653 ± 3632 a | |
| (ng/mg protein) | Placebo | 7679 ± 2442 a | 8560 ± 416 a | 0.8352 |
| GPx | NACQ | 13.81 ± 4.83 a | 17.55 ± 5.00 a | |
| (mmol/min/mg protein) | Placebo | 25.53 ± 5.08 a | 21.52 ± 5.53 a | 0.2065 |
| SOD | NACQ | 7.178 ± 1.238 a | 6.604 ± 0.792 a | |
| (U/mg protein) | Placebo | 7.377 ± 1.551 a | 7.390 ± 1.187 a | 0.6191 |
| Gene ID | Protein Name | Adjusted p Value | Log2 Fold Change |
|---|---|---|---|
| Increased expression | |||
| MB | Myoglobin | 1.90 × 10−3 | 0.22 |
| Mitochondria | |||
| TCA cycle | |||
| IDH2 | Isocitrate Dehydrogenase [NADP] Mitochondrial | 2.10 × 10−4 | 0.12 |
| ACO2 | Aconitate Hydratase Mitochondrial | 5.00 × 10−4 | 0.11 |
| MDH2 | Malate Dehydrogenase Mitochondrial | 4.00 × 10−3 | 0.11 |
| FH | Fumarate Hydratase Mitochondrial | 2.00 × 10−3 | 0.11 |
| OGDH | 2-Oxogluterate Dehydrogenase Mitochondrial Isoform X3 | 8.70 × 10−4 | 0.09 |
| Electron Transfer System | |||
| COX5A | Cytochrome C Oxidase Subunit 5A Mitochondrial | 8.90 × 10−4 | 0.18 |
| COX4I1 | Cytochrome C Oxidase Subunit 4 Isoform 1 Mitochondrial | 5.00 × 10−4 | 0.13 |
| ATP5F1B | ATP Synthase Subunit beta Mitochondrial | 1.00 × 10−4 | 0.10 |
| Fat metabolism | |||
| ACADVL | Very Long-Chain Specific Acyl-CoA Dehydrogenase Mitochondrial | 1.00 × 10−3 | 0.11 |
| ASCL1 | Long-Chain-Fatty-Acid–CoA Ligase 1 Isoform X2 | 2.70 × 10−3 | 0.10 |
| FABP3 | Fatty Acid-Binding protein Heart | 1.00 × 10−3 | 0.27 |
| Other | |||
| NNT | NAD(P) Transhydrogenase Mitochondrial Isoform X1 | 3.00 × 10−3 | 0.07 |
| VDAC2 | Voltage-Dependent Anion-Selective Channel Protein 2 | 2.00 × 10−3 | 0.14 |
| Sarcomere Proteins | |||
| Z disc | |||
| CRYAB | Alpha-Crystallin B Chain | 1.00 × 10−4 | 0.28 |
| ACTN2 | Alpha-Actinin-2 Isoform X1 | 1.00 × 10−4 | 0.16 |
| FLNC | Filamin-C Isoform X1 | 1.30 × 10−4 | 0.12 |
| PDLIM5 | PDZ and LIM Domain Protein 5 Isoform X9 | 7.20 × 10−4 | 0.12 |
| Myosin | |||
| MYBPC1 | Myosin-Binding Protein C Slow-Type Isoform X2 | 1.00 × 10−4 | 0.11 |
| Blood Proteins | |||
| ALB | Serum Albumin Precursor | 1.00 × 10−4 | 0.38 |
| APOA1 | Apolipoprotein A-I | 1.00 × 10−3 | 0.34 |
| LOC100061692 | Alpha-2 Macroglobulin | 3.50 × 10−3 | 0.22 |
| Decreased expression | |||
| Cysteine Synthesis | |||
| ARMT1 | Protein-Glutamate O-Methyltransferase | 7.30 × 10−4 | −0.40 |
| Glycolysis/Gluconeogenesis | |||
| PFKM | ATP-Dependent 6-Phosphofructokinase Muscle Type Isoform X3 | 2.00 × 10−3 | −0.07 |
| PKM | Pyruvate Kinase PKM Isoform M1 | 1.00 × 10−4 | −0.10 |
| LDHA | L-lactate Dehydrogenase A Chain | 1.80 × 10−4 | −0.10 |
| FBP2 | Fructose -1,6-bisphosphatase isozyme 2 | 4.00 × 10−3 | −0.11 |
| GPI | Glucose-6-Phosphate Isomerase | 1.00 × 10−4 | −0.14 |
| PHKA1 | Phosphorylase B Kinase Regulatory Subunit alpha Skeletal Muscle Isoform X1 | 1.00 × 10−3 | −0.14 |
| PHKB | Phosphorylase B Kinase Regulatory Subunit beta | 2.00 × 10−3 | −0.14 |
| ENO3 | Beta-Enolase | 1.00 × 10−4 | −0.15 |
| PGM1 | Phosphoglucomutase-1 Isoform X2 | 1.00 × 10−4 | −0.15 |
| Sarcomere | |||
| MYOM1 | Myomesin-1 Isoform X4 | 1.00 × 10−3 | −0.08 |
| MYOM2 | Myomesin-2 Isoform X3 | 1.00 × 10−4 | −0.12 |
| MYH1 | Myosin-1 Isoform X1 | 2.70 × 10−4 | −0.11 |
| MYBPC2 | Myosin-Binding Protein C Fast-Type | 1.00 × 10−4 | −0.17 |
| MYLK2 | Myosin Light Chain Kinase 2 Skeletal/Cardiac Muscle | 2.50 × 10−3 | −0.20 |
| Calcium Regulation | |||
| SRL | Sarcalumenin Isoform X1 | 1.00 × 10−3 | −0.09 |
| ATP2A1 | Sarcoplasmic/Endoplasmic Reticulum Calcium ATPase 1 Isoform X1 | 1.00 × 10−4 | −0.17 |
| ANXA6 | Annexin A6 Isoform X1 | 1.00 × 10−4 | −0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henry, M.L.; Velez-Irizarry, D.; Pagan, J.D.; Sordillo, L.; Gandy, J.; Valberg, S.J. The Impact of N-Acetyl Cysteine and Coenzyme Q10 Supplementation on Skeletal Muscle Antioxidants and Proteome in Fit Thoroughbred Horses. Antioxidants 2021, 10, 1739. https://doi.org/10.3390/antiox10111739
Henry ML, Velez-Irizarry D, Pagan JD, Sordillo L, Gandy J, Valberg SJ. The Impact of N-Acetyl Cysteine and Coenzyme Q10 Supplementation on Skeletal Muscle Antioxidants and Proteome in Fit Thoroughbred Horses. Antioxidants. 2021; 10(11):1739. https://doi.org/10.3390/antiox10111739
Chicago/Turabian StyleHenry, Marisa L., Deborah Velez-Irizarry, Joe D. Pagan, Lorraine Sordillo, Jeff Gandy, and Stephanie J. Valberg. 2021. "The Impact of N-Acetyl Cysteine and Coenzyme Q10 Supplementation on Skeletal Muscle Antioxidants and Proteome in Fit Thoroughbred Horses" Antioxidants 10, no. 11: 1739. https://doi.org/10.3390/antiox10111739
APA StyleHenry, M. L., Velez-Irizarry, D., Pagan, J. D., Sordillo, L., Gandy, J., & Valberg, S. J. (2021). The Impact of N-Acetyl Cysteine and Coenzyme Q10 Supplementation on Skeletal Muscle Antioxidants and Proteome in Fit Thoroughbred Horses. Antioxidants, 10(11), 1739. https://doi.org/10.3390/antiox10111739