
Glycoarray Technologies: Deciphering Interactions from Proteins to Live Cell Responses

Abstract
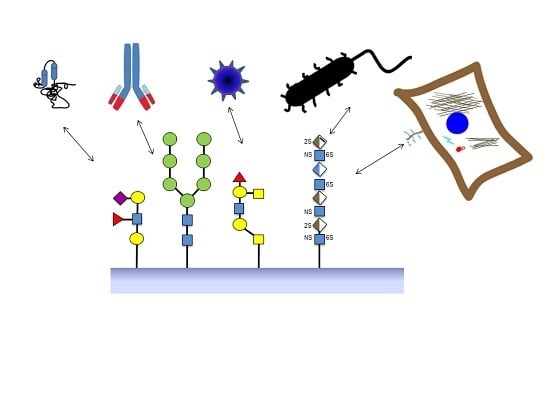
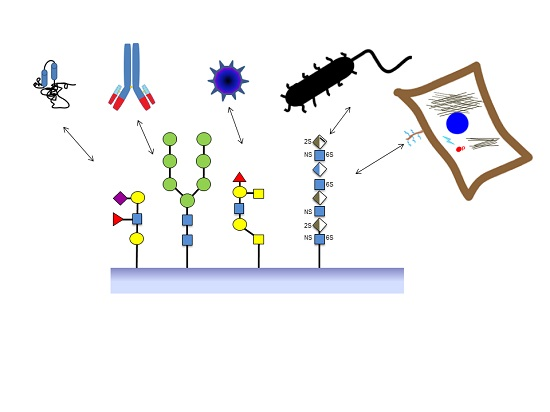
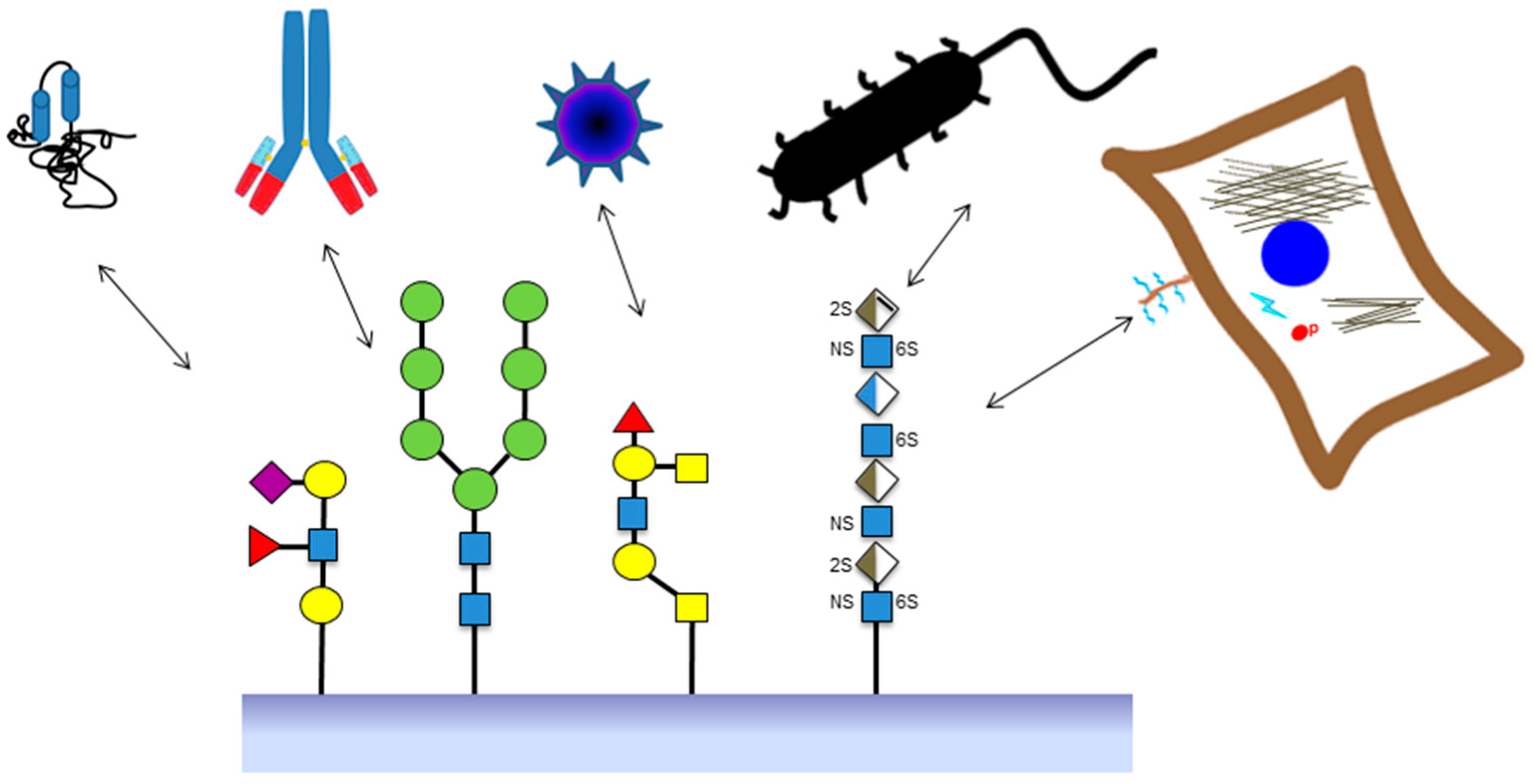
:
1. Introduction
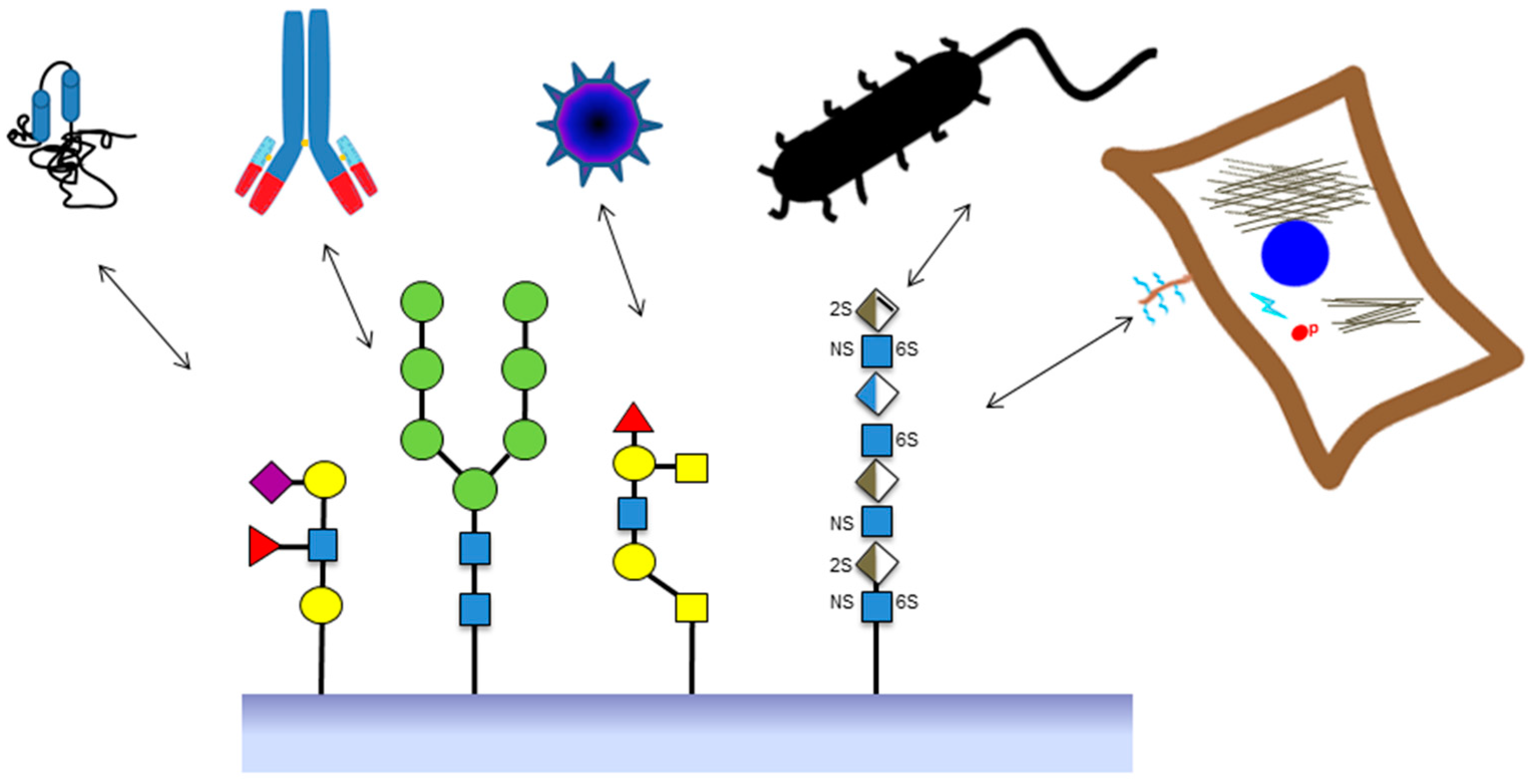
2. Sources of Glycan Structures
3. Choice of Solid Supports and Immobilization of Glycans onto Microarrays
3.1. Affinity Adsorption
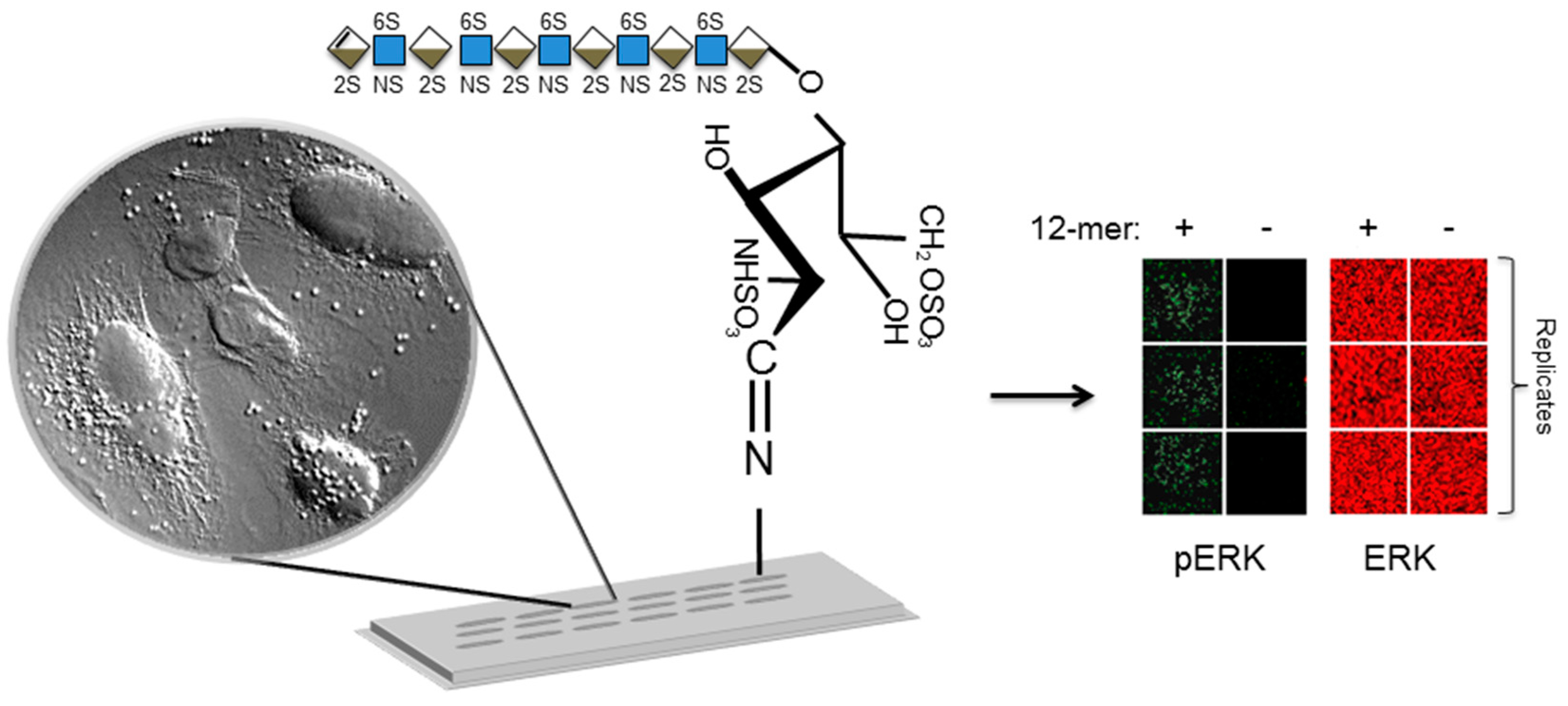
3.2. Covalent Immobilization of Glycans

{kind=link}
{kind=link}
{kind=link}
| Type of Interaction | Type of Reaction | Details of Immobilization | Reference |
|---|---|---|---|
| Covalent | Condensation | Unmodified carbohydrates onto hydrazide surfaces. | [22,40,41,42] |
| Covalent | Michael Addition | Malemide-linked carbohydrates and thiol-coated glass slides. | [27,43] |
| Covalent | Epoxide opening | Covalently attach carbohydrates, glycoproteins and neoglycoconjugates to glass slides. | [44,45,46] |
| Covalent | Amide coupling | – | – |
| Covalent | Diels-Alder reaction | Covalent immobilization of glycans by cycloaddition. | [31] |
| Covalent | Carbene | Covalent immobilization of glycans by insertion. | [35] |
| Covalent | Radical coupling | Covalent immobilization of unmodified glycans by insertion. | [47] |
3.3. Site-Specific Covalent Immobilization
3.4. Non-Covalent Immobilization
4. Techniques for Detection of Protein Binding to Glycoarrays
5. Use of Glycan Arrays for the High-Throughput Analysis of Glycan–Protein Interactions
6. Use of Glycan Arrays for Studying Heparin/Heparan Sulfate Interactions with Proteins
7. Glycoarrays for Measuring Glycan–Cell Interactions
8. Glycoarrays for Measuring Virus and Bacteria–Glycan Interactions
9. Glycoarrays for Reporting Live Cell Responses Including Cellular Signaling Pathways

10. Interrogation of Glycoarrays Using Mass Spectrometry
11. Conclusions and Future Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Prydz, K.; Dalen, K.T. Synthesis and sorting of proteoglycans. J. Cell Sci. 2000, 113, 193–205. [Google Scholar] [PubMed]
- Boehm, G.; Stahl, B. Oligosaccharides from milk. J. Nutr. 2007, 137 (Suppl. S2), 847S–849S. [Google Scholar] [PubMed]
- Haltiwanger, R.S.; Lowe, J.B. Role of glycosylation in development. Annu. Rev. Biochem. 2004, 73, 491–537. [Google Scholar] [CrossRef] [PubMed]
- Bertozzi, C.R.; Sasisekharan, R. Glycomics. In Essentials of Glycobiology, 2nd ed.; Ajit, V., Richard, D.C., Jeffrey, D.E., Hudson, H.F., Pamela, S., Carolyn, R.B., Gerald, W.H., Marilynn, E.E., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009. [Google Scholar]
- Turnbull, J.E.; Field, R.A. Emerging glycomics technologies. Nat. Chem. Biol. 2007, 3, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Aoki-Kinoshita, K.F. Glycome Informatics- Methods and Applications; CRC Press: New York, NY, USA, 2010. [Google Scholar]
- Varki, A.; Cummings, R.D.; Esko, J.D.; Freeze, H.H.; Stanley, P.; Marth, J.D.; Bertozzi, C.R.; Hart, G.W.; Etzler, M.E. Symbol nomenclature for glycan representation. Proteomics 2009, 9, 5398–5399. [Google Scholar] [CrossRef] [PubMed]
- Wang, D. Carbohydrate microarrays. Proteomics 2003, 3, 2167–2175. [Google Scholar] [CrossRef] [PubMed]
- Fukui, S.; Feizi, T.; Galustian, C.; Lawson, A.M.; Chai, W. Oligosaccharide microarrays for high-throughput detection and specificity assignments of carbohydrate-protein interactions. Nat. Biotechnol. 2002, 20, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Nagasawa, K. Selective N-desulfation of heparin with dimethyl sulfoxide containing water or methanol. Carbohydr. Res. 1976, 46, 87–95. [Google Scholar] [CrossRef]
- Guimond, S.E.; Puvirajesinghe, T.M.; Skidmore, M.A.; Kalus, I.; Dierks, T.; Yates, E.A.; Turnbull, J.E. Rapid purification and high sensitivity analysis of heparan sulfate from cells and tissues: Toward glycomics profiling. J. Biol. Chem. 2009, 284, 25714–25722. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.K.; Ahmed, Y.A.; Yates, E.A.; Turnbull, J.E. Generating heparan sulfate saccharide libraries for glycomics applications. Nat. Protoc. 2010, 5, 821–833. [Google Scholar] [CrossRef] [PubMed]
- Grootenhuis, P.D.; Westerduin, P.; Meuleman, D.; Petitou, M.; van Boeckel, C.A. Rational design of synthetic heparin analogues with tailor-made coagulation factor inhibitory activity. Nat. Struct. Biol. 1995, 2, 736–739. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.K.; Yates, E.A.; Fernig, D.G.; Turnbull, J.E. Interactions of heparin/heparan sulfate with proteins: Appraisal of structural factors and experimental approaches. Glycobiology 2004, 14, 17R–30R. [Google Scholar] [CrossRef] [PubMed]
- Seeberger, P.H.; Finney, N.; Rabuka, D.; Bertozzi, C.R. Chemical and Enzymatic Synthesis of Glycans and Glycoconjugates; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009. [Google Scholar]
- Palcic, M.M. Glycosyltransferases as biocatalysts. Curr. Opin. Chem. Biol. 2011, 15, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Blixt, O.; Collins, B.E.; van den Nieuwenhof, I.M.; Crocker, P.R.; Paulson, J.C. Sialoside specificity of the siglec family assessed using novel multivalent probes: Identification of potent inhibitors of myelin-associated glycoprotein. J. Biol. Chem. 2003, 278, 31007–31019. [Google Scholar] [CrossRef] [PubMed]
- Merrifield, R.B. Solid Phase Peptide Synthesis. I. The Synthesis of a Tetrapeptide. J. Am. Chem. Soc. 1963, 85, 2149–2154. [Google Scholar] [CrossRef]
- De Paz, J.L.; Noti, C.; Seeberger, P.H. Microarrays of synthetic heparin oligosaccharides. J. Am. Chem. Soc. 2006, 128, 2766–2767. [Google Scholar] [CrossRef] [PubMed]
- Plante, O.J.; Palmacci, E.R.; Seeberger, P.H. Automated solid-phase synthesis of oligosaccharides. Science 2001, 291, 1523–1527. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Li, L.; Harvey, C.; Liu, J.; Linhardt, R.J. Towards the chemoenzymatic synthesis of heparan sulfate oligosaccharides: Oxidative cleavage of -nitrophenyl group with ceric ammonium salts. Tetrahedron Lett. 2013, 54, 4471–4474. [Google Scholar] [CrossRef] [PubMed]
- Zhi, Z.L.; Powell, A.K.; Turnbull, J.E. Fabrication of carbohydrate microarrays on gold surfaces: Direct attachment of nonderivatized oligosaccharides to hydrazide-modified self-assembled monolayers. Anal. Chem. 2006, 78, 4786–4793. [Google Scholar] [CrossRef] [PubMed]
- Wehner, J.W.; Weissenborn, M.J.; Hartmann, M.; Gray, C.J.; Sardzik, R.; Eyers, C.E.; Flitsch, S.L.; Lindhorst, T.K. Dual purpose S-trityl-linkers for glycoarray fabrication on both polystyrene and gold. Org. Biomol. Chem. 2012, 10, 8919–8926. [Google Scholar] [CrossRef] [PubMed]
- Ratner, D.M. Carbohydrate microarrays: Advancing the burgeoning field of glycomics. Biol. Tech. Int. 2005, 17, 8–11. [Google Scholar]
- Bryan, M.C.; Plettenburg, O.; Sears, P.; Rabuka, D.; Wacowich-Sgarbi, S.; Wong, C.H. Saccharide display on microtiter plates. Chem. Biol. 2002, 9, 713–720. [Google Scholar] [CrossRef]
- Fazio, F.; Bryan, M.C.; Blixt, O.; Paulson, J.C.; Wong, C.H. Synthesis of sugar arrays in microtiter plate. J. Am. Chem. Soc. 2002, 124, 14397–14402. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Shin, I. Fabrication of carbohydrate chips for studying protein-carbohydrate interactions. Angew. Chem. Int. Ed. Engl. 2002, 41, 3180–3182. [Google Scholar] [CrossRef]
- De Boer, A.R.; Hokke, C.H.; Deelder, A.M.; Wuhrer, M. General microarray technique for immobilization and screening of natural glycans. Anal. Chem. 2007, 79, 8107–8113. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Xia, B.; Stowell, S.R.; Lasanajak, Y.; Smith, D.F.; Cummings, R.D. Novel fluorescent glycan microarray strategy reveals ligands for galectins. Chem. Biol. 2009, 16, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, S.; Trummer, B.J.; Deng, C.; Wang, A. Carbohydrate microarrays for the recognition of cross-reactive molecular markers of microbes and host cells. Nat. Biotechnol. 2002, 20, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Houseman, B.T.; Mrksich, M. Carbohydrate arrays for the evaluation of protein binding and enzymatic modification. Chem. Biol. 2002, 9, 443–454. [Google Scholar] [CrossRef]
- Ma, X.; Mohammad, S.F.; Kim, S.W. Heparin removal from blood using poly(l-lysine) immobilized hollow fiber. Biotechnol. Bioeng. 1992, 40, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Barie, N.; Rapp, M.; Sigrist, H.; Ache, H.J. Covalent photolinker-mediated immobilization of an intermediate dextran layer to polymer-coated surfaces for biosensing applications. Biosens. Bioelectron. 1998, 13, 855–860. [Google Scholar] [CrossRef]
- Platz, M.; Admasu, A.S.; Kwiatkowski, S.; Crocker, P.J.; Imai, N.; Watt, D.S. Photolysis of 3-aryl-3-(trifluoromethyl)diazirines: A caveat regarding their use in photoaffinity probes. Bioconjug. Chem. 1991, 2, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Angeloni, S.; Ridet, J.L.; Kusy, N.; Gao, H.; Crevoisier, F.; Guinchard, S.; Kochhar, S.; Sigrist, H.; Sprenger, N. Glycoprofiling with micro-arrays of glycoconjugates and lectins. Glycobiology 2005, 15, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Yates, E.A.; Jones, M.O.; Clarke, C.E.; Powell, A.K.; Johnson, S.R.; Porch, A.; Edwards, P.P.; Turnbull, J.E. Microwave enhanced reaction of carbohydrates with amino-derivatised labels and glass surfaces. J. Mater. Chem. 2003, 13, 2061–2063. [Google Scholar] [CrossRef]
- Powell, A.K.; Zhi, Z.L.; Turnbull, J.E. Saccharide microarrays for high-throughput interrogation of glycan-protein binding interactions. Methods Mol. Biol. 2009, 534, 313–329. [Google Scholar] [PubMed]
- Zhi, Z.L.; Laurent, N.; Powell, A.K.; Karamanska, R.; Fais, M.; Voglmeir, J.; Wright, A.; Blackburn, J.M.; Crocker, P.R.; Russell, D.A.; et al. A versatile gold surface approach for fabrication and interrogation of glycoarrays. ChemBioChem 2008, 9, 1568–1575. [Google Scholar] [CrossRef] [PubMed]
- Puvirajesinghe, T.M.; Ahmed, Y.A.; Powell, A.K.; Fernig, D.G.; Guimond, S.E.; Turnbull, J.E. Array-based functional screening of heparin glycans. Chem. Biol. 2012, 19, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Shin, I. Facile preparation of carbohydrate microarrays by site-specific, covalent immobilization of unmodified carbohydrates on hydrazide-coated glass slides. Org. Lett. 2005, 7, 4269–4272. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Wong, D.T. Lectin microarray. Proteom. Clin. Appl. 2009, 3, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Houseman, B.T.; Gawalt, E.S.; Mrksich, M. Maleimide-functionalized self-assembled monolayers for the preparation of peptide and carbohydrate biochips. Langmuir 2003, 19, 1522–1531. [Google Scholar] [CrossRef]
- Park, S.; Lee, M.R.; Pyo, S.J.; Shin, I. Carbohydrate chips for studying high-throughput carbohydrate-protein interactions. J. Am. Chem. Soc. 2004, 126, 4812–4819. [Google Scholar] [CrossRef] [PubMed]
- Oyelaran, O.; Li, Q.; Farnsworth, D.; Gildersleeve, J.C. Microarrays with varying carbohydrate density reveal distinct subpopulations of serum antibodies. J. Proteome Res. 2009, 8, 3529–3538. [Google Scholar] [CrossRef] [PubMed]
- Oyelaran, O.; McShane, L.M.; Dodd, L.; Gildersleeve, J.C. Profiling human serum antibodies with a carbohydrate antigen microarray. J. Proteome Res. 2009, 8, 4301–4310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Campbell, C.; Li, Q.; Gildersleeve, J.C. Multidimensional glycan arrays for enhanced antibody profiling. Mol. Biosyst. 2010, 6, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Carroll, G.T.; Wang, D.; Turro, N.J.; Koberstein, J.T. Photochemical micropatterning of carbohydrates on a surface. Langmuir 2006, 22, 2899–2905. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Adachi, K.; Lee, B.K.; Kang, D.G.; Kim, Y.K.; Kim, K.R.; Lee, H.Y.; Kawai, T.; Cha, H.J. Facile and rapid direct gold surface immobilization with controlled orientation for carbohydrates. Bioconjug. Chem. 2007, 18, 2197–2201. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Ruiz, A.; Serna, S.; Ruiz, N.; Martin-Lomas, M.; Reichardt, N.C. MALDI-TOF mass spectrometric analysis of enzyme activity and lectin trapping on an array of N-glycans. Angew. Chem. Int. Ed. Engl. 2011, 50, 1801–1804. [Google Scholar] [CrossRef] [PubMed]
- Vora, G.J.; Meador, C.E.; Anderson, G.P.; Taitt, C.R. Comparison of detection and signal amplification methods for DNA microarrays. Mol. Cell Probes 2008, 22, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Fero, M.; Pogliano, K. Automated quantitative live cell fluorescence microscopy. Cold Spring Harb. Perspect. Biol. 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Horstmeyer, R.; Yang, C. Wide-field, high-resolution Fourier ptychographic microscopy. Nat. Photonics 2013, 7, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Varki, A. Modulation of glycan recognition by clustered saccharide patches. Int. Rev. Cell Mol. Biol. 2014, 308, 75–125. [Google Scholar] [PubMed]
- Leppanen, A.; Penttila, L.; Renkonen, O.; McEver, R.P.; Cummings, R.D. Glycosulfopeptides with O-glycans containing sialylated and polyfucosylated polylactosamine bind with low affinity to P-selectin. J. Biol. Chem. 2002, 277, 39749–39759. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Liu, Y.; Zhang, H.; Zhang, Y.; Fukuda, M.N.; Palma, A.S.; Kozak, R.P.; Childs, R.A.; Nonaka, M.; Li, Z.; et al. Carbohydrate sequence of the prostate cancer-associated antigen F77 assigned by a mucin O-glycome designer array. J. Biol. Chem. 2014, 289, 16462–16477. [Google Scholar] [CrossRef] [PubMed]
- Palma, A.S.; Liu, Y.; Zhang, H.; Zhang, Y.; McCleary, B.V.; Yu, G.; Huang, Q.; Guidolin, L.S.; Ciocchini, A.E.; Torosantucci, A.; et al. Unravelling glucan recognition systems by glycome microarrays using the designer approach and mass spectrometry. Mol. Cell Proteom. 2015, 14, 974–988. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, S.; Brennan, K.M.; Willison, H.J. Heteromeric glycolipid complexes as modulators of autoantibody and lectin binding. Prog. Lipid Res. 2010, 49, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Galban-Horcajo, F.; Halstead, S.K.; McGonigal, R.; Willison, H.J. The application of glycosphingolipid arrays to autoantibody detection in neuroimmunological disorders. Curr. Opin. Chem. Biol. 2014, 18, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Van Horssen, J.; Wesseling, P.; van den Heuvel, L.P.; de Waal, R.M.; Verbeek, M.M. Heparan sulphate proteoglycans in Alzheimer’s disease and amyloid-related disorders. Lancet Neurol. 2003, 2, 482–492. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, H.; Jiang, L.; Zhou, X.; Lu, C.; Zhang, X. Positive association of heparanase expression with tumor invasion and lymphatic metastasis in gastric carcinoma. Mod. Pathol. 2004, 18, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Sasisekharan, C.R. Essentials of Glycobiology; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009. [Google Scholar]
- McFeters, A.G.; Yu, F.P.; Pyle, B.H.; Stewart, P.S. Physiological assessment of bacteria using fluorochromes. J. Microbiol. Methods 1995, 21, 1–13. [Google Scholar] [CrossRef]
- De Paz, J.L.; Angulo, J.; Lassaletta, J.M.; Nieto, P.M.; Redondo-Horcajo, M.; Lozano, R.M.; Gimenez-Gallego, G.; Martin-Lomas, M. The activation of fibroblast growth factors by heparin: synthesis, structure, and biological activity of heparin-like oligosaccharides. ChemBioChem 2001, 2, 673–685. [Google Scholar] [CrossRef]
- Esko, J.D.; Lindahl, U. Molecular diversity of heparan sulfate. J. Clin. Investig. 2001, 108, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, U.; Thunberg, L.; Backstrom, G.; Riesenfeld, J.; Nordling, K.; Bjork, I. Extension and structural variability of the antithrombin-binding sequence in heparin. J. Biol. Chem. 1984, 259, 12368–12376. [Google Scholar] [PubMed]
- Guimond, S.; Maccarana, M.; Olwin, B.B.; Lindahl, U.; Rapraeger, A.C. Activating and inhibitory heparin sequences for FGF-2 (basic FGF). Distinct requirements for FGF-1, FGF-2, and FGF-4. J. Biol. Chem. 1993, 268, 23906–23914. [Google Scholar] [PubMed]
- Pye, D.A.; Vives, R.R.; Turnbull, J.E.; Hyde, P.; Gallagher, J.T. Heparan sulfate oligosaccharides require 6-O-sulfation for promotion of basic fibroblast growth factor mitogenic activity. J. Biol. Chem. 1998, 273, 22936–22942. [Google Scholar] [CrossRef] [PubMed]
- Deakin, J.A.; Lyon, M. Differential regulation of hepatocyte growth factor/scatter factor by cell surface proteoglycans and free glycosaminoglycan chains. J. Cell Sci. 1999, 112, 1999–2009. [Google Scholar] [PubMed]
- Yates, E.A.; Guimond, S.E.; Turnbull, J.E. Highly diverse heparan sulfate analogue libraries: Providing access to expanded areas of sequence space for bioactivity screening. J. Med. Chem. 2004, 47, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Orgueira, H.A.; Bartolozzi, A.; Schell, P.; Litjens, R.E.; Palmacci, E.R.; Seeberger, P.H. Modular synthesis of heparin oligosaccharides. Chemistry 2003, 9, 140–169. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Gildersleeve, J.C.; Blixt, O.; Shin, I. Carbohydrate microarrays. Chem. Soc. Rev. 2013, 42, 4310–4326. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Shin, I. Carbohydrate microarrays for assaying galactosyltransferase activity. Org. Lett. 2007, 9, 1675–1678. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.H.; Wang, S.K.; Wong, C.H. Quantitative analysis of carbohydrate-protein interactions using glycan microarrays: Determination of surface and solution dissociation constants. J. Am. Chem. Soc. 2007, 129, 11177–11184. [Google Scholar] [CrossRef] [PubMed]
- Vigerust, D.J.; Shepherd, V.L. Virus glycosylation: Role in virulence and immune interactions. Trends Microbiol. 2007, 15, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; Kwon, D.S.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Middel, J.; Cornelissen, I.L.; Nottet, H.S.; KewalRamani, V.N.; Littman, D.R.; et al. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell 2000, 100, 587–597. [Google Scholar] [CrossRef]
- Engering, A.; Geijtenbeek, T.B.; van Vliet, S.J.; Wijers, M.; van Liempt, E.; Demaurex, N.; Lanzavecchia, A.; Fransen, J.; Figdor, C.G.; Piguet, V.; et al. The dendritic cell-specific adhesion receptor DC-SIGN internalizes antigen for presentation to T cells. J. Immunol. 2002, 168, 2118–2126. [Google Scholar] [CrossRef] [PubMed]
- Powlesland, A.S.; Ward, E.M.; Sadhu, S.K.; Guo, Y.; Taylor, M.E.; Drickamer, K. Widely divergent biochemical properties of the complete set of mouse DC-SIGN-related proteins. J. Biol. Chem. 2006, 281, 20440–20449. [Google Scholar] [CrossRef] [PubMed]
- Nimrichter, L.; Gargir, A.; Gortler, M.; Altstock, R.T.; Shtevi, A.; Weisshaus, O.; Fire, E.; Dotan, N.; Schnaar, R.L. Intact cell adhesion to glycan microarrays. Glycobiology 2004, 14, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Bewley, C.A.; Otero-Quintero, S. The potent anti-HIV protein cyanovirin-N contains two novel carbohydrate binding sites that selectively bind to Man8 D1D3 and Man9 with nanomolar affinity: Implications for binding to the HIV envelope protein gp120. J. Am. Chem. Soc. 2001, 123, 3892–3902. [Google Scholar] [CrossRef] [PubMed]
- Ratner, D.M.; Adams, E.W.; Su, J.; O’Keefe, B.R.; Mrksich, M.; Seeberger, P.H. Probing protein-carbohydrate interactions with microarrays of synthetic oligosaccharides. ChemBioChem 2004, 5, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.E.; Ratner, D.M.; Bokesch, H.R.; McMahon, J.B.; O’Keefe, B.R.; Seeberger, P.H. Oligosaccharide and glycoprotein microarrays as tools in HIV glycobiology; glycan-dependent gp120/protein interactions. Chem. Biol. 2004, 11, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Gordon, J.I. Glycans as legislators of host-microbial interactions: Spanning the spectrum from symbiosis to pathogenicity. Glycobiology 2001, 11, 1R–10R. [Google Scholar] [CrossRef] [PubMed]
- CodeLink Activated slides. Available online: http://www.surmodics.com/assets/uploads/documents/CodeLink_User_Guide.pdf (accessed on 24 December 2015).
- De Paz, J.L.; Horlacher, T.; Seeberger, P.H. Oligosaccharide Microarrays to Map Interactions of Carbohydrates in Biological Systems; Elsevier Inc.: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Mahal, L.K. Catching bacteria with sugar. Chem. Biol. 2004, 11, 1602–1604. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.C.; Ping-Sheng, L.; Zhai, H.X.; Shen, T.Y.; Mensa-Wilmot, K. Phosphatidylinositol phospholipase C is activated allosterically by the aminoglycoside G418. 2-deoxy-2-fluoro-scyllo-inositol-1-O-dodecylphosphonate and its analogs inhibit glycosylphosphatidylinositol phospholipase C. J. Biol. Chem. 1996, 271, 15468–15477. [Google Scholar] [PubMed]
- Ren, Y.G.; Martinez, J.; Kirsebom, L.A.; Virtanen, A. Inhibition of Klenow DNA polymerase and poly(A)-specific ribonuclease by aminoglycosides. RNA 2002, 8, 1393–1400. [Google Scholar] [CrossRef] [PubMed]
- Moazed, D.; Noller, H.F. Interaction of antibiotics with functional sites in 16S ribosomal RNA. Nature 1987, 327, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Walter, F.; Vicens, Q.; Westhof, E. Aminoglycoside-RNA interactions. Curr. Opin. Chem. Biol. 1999, 3, 694–704. [Google Scholar] [CrossRef]
- Jana, S.; Deb, J.K. Molecular understanding of aminoglycoside action and resistance. Appl. Microbiol. Biotechnol. 2006, 70, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Delehedde, M.; Sergeant, N.; Lyon, M.; Rudland, P.S.; Fernig, D.G. Hepatocyte growth factor/scatter factor stimulates migration of rat mammary fibroblasts through both mitogen-activated protein kinase and phosphatidylinositol 3-kinase/Akt pathways. Eur. J. Biochem. 2001, 268, 4423–4429. [Google Scholar] [CrossRef] [PubMed]
- Lundin, L.; Larsson, H.; Kreuger, J.; Kanda, S.; Lindahl, U.; Salmivirta, M.; Claesson-Welsh, L. Selectively desulfated heparin inhibits fibroblast growth factor-induced mitogenicity and angiogenesis. J. Biol. Chem. 2000, 275, 24653–24660. [Google Scholar] [CrossRef] [PubMed]
- Hazzalin, C.A.; Mahadevan, L.C. MAPK-regulated transcription: A continuously variable gene switch? Nat. Rev. Mol. Cell Biol. 2002, 3, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Raman, M.; Chen, W.; Cobb, M.H. Differential regulation and properties of MAPKs. Oncogene 2007, 26, 3100–3112. [Google Scholar] [CrossRef] [PubMed]
- Guimond, S.E.; Turnbull, J.E. Fibroblast growth factor receptor signalling is dictated by specific heparan sulphate saccharides. Curr. Biol. 1999, 9, 1343–1346. [Google Scholar] [CrossRef]
- Delehedde, M.; Lyon, M.; Gallagher, J.T.; Rudland, P.S.; Fernig, D.G. Fibroblast growth factor-2 binds to small heparin-derived oligosaccharides and stimulates a sustained phosphorylation of p42/44 mitogen-activated protein kinase and proliferation of rat mammary fibroblasts. Biochem. J. 2002, 366, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, J.E.; Fernig, D.G.; Ke, Y.; Wilkinson, M.C.; Gallagher, J.T. Identification of the basic fibroblast growth factor binding sequence in fibroblast heparan sulfate. J. Biol. Chem. 1992, 267, 10337–10341. [Google Scholar] [PubMed]
- Ishihara, M.; Shaklee, P.N.; Yang, Z.; Liang, W.; Wei, Z.; Stack, R.J.; Holme, K. Structural features in heparin which modulate specific biological activities mediated by basic fibroblast growth factor. Glycobiology 1994, 4, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Sterner, E.; Meli, L.; Kwon, S.J.; Dordick, J.S.; Linhardt, R.J. FGF-FGFR signaling mediated through glycosaminoglycans in microtiter plate and cell-based microarray platforms. Biochemistry 2013, 52, 9009–9019. [Google Scholar] [CrossRef] [PubMed]
- Ornitz, D.M.; Xu, J.; Colvin, J.S.; McEwen, D.G.; MacArthur, C.A.; Coulier, F.; Gao, G.; Goldfarb, M. Receptor specificity of the fibroblast growth factor family. J. Biol. Chem. 1996, 271, 15292–15297. [Google Scholar] [PubMed]
- Yu, F.P.; Pyle, B.H.; McFeters, G.A. A direct viable count method for the enumeration of attached bacteria and assessment of biofilm disinfection. J. Microbiol. Methods 1993, 17, 167–180. [Google Scholar] [CrossRef]
- Papadopoulos, N.G.; Dedoussis, G.V.; Spanakos, G.; Gritzapis, A.D.; Baxevanis, C.N.; Papamichail, M. An improved fluorescence assay for the determination of lymphocyte-mediated cytotoxicity using flow cytometry. J. Immunol. Methods 1994, 177, 101–111. [Google Scholar] [CrossRef]
- Hayes, A.W. Principles and Methods of Toxicology, 5th ed.; Hayes, A.W., Ed.; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Feizi, T.; Chai, W. Oligosaccharide microarrays to decipher the glyco code. Nat. Rev. Mol. Cell Biol. 2004, 5, 582–588. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puvirajesinghe, T.M.; Turnbull, J.E. Glycoarray Technologies: Deciphering Interactions from Proteins to Live Cell Responses. Microarrays 2016, 5, 3. https://doi.org/10.3390/microarrays5010003
Puvirajesinghe TM, Turnbull JE. Glycoarray Technologies: Deciphering Interactions from Proteins to Live Cell Responses. Microarrays. 2016; 5(1):3. https://doi.org/10.3390/microarrays5010003
Chicago/Turabian StylePuvirajesinghe, Tania M., and Jeremy. E. Turnbull. 2016. "Glycoarray Technologies: Deciphering Interactions from Proteins to Live Cell Responses" Microarrays 5, no. 1: 3. https://doi.org/10.3390/microarrays5010003
APA StylePuvirajesinghe, T. M., & Turnbull, J. E. (2016). Glycoarray Technologies: Deciphering Interactions from Proteins to Live Cell Responses. Microarrays, 5(1), 3. https://doi.org/10.3390/microarrays5010003