Transcriptome Analysis of Porphyrin-Accumulated and X-Ray-Irradiated Cell Cultures under Limited Proliferation and Non-Lethal Conditions



Abstract
:1. Introduction
2. Experimental Section
3. Results and Discussion
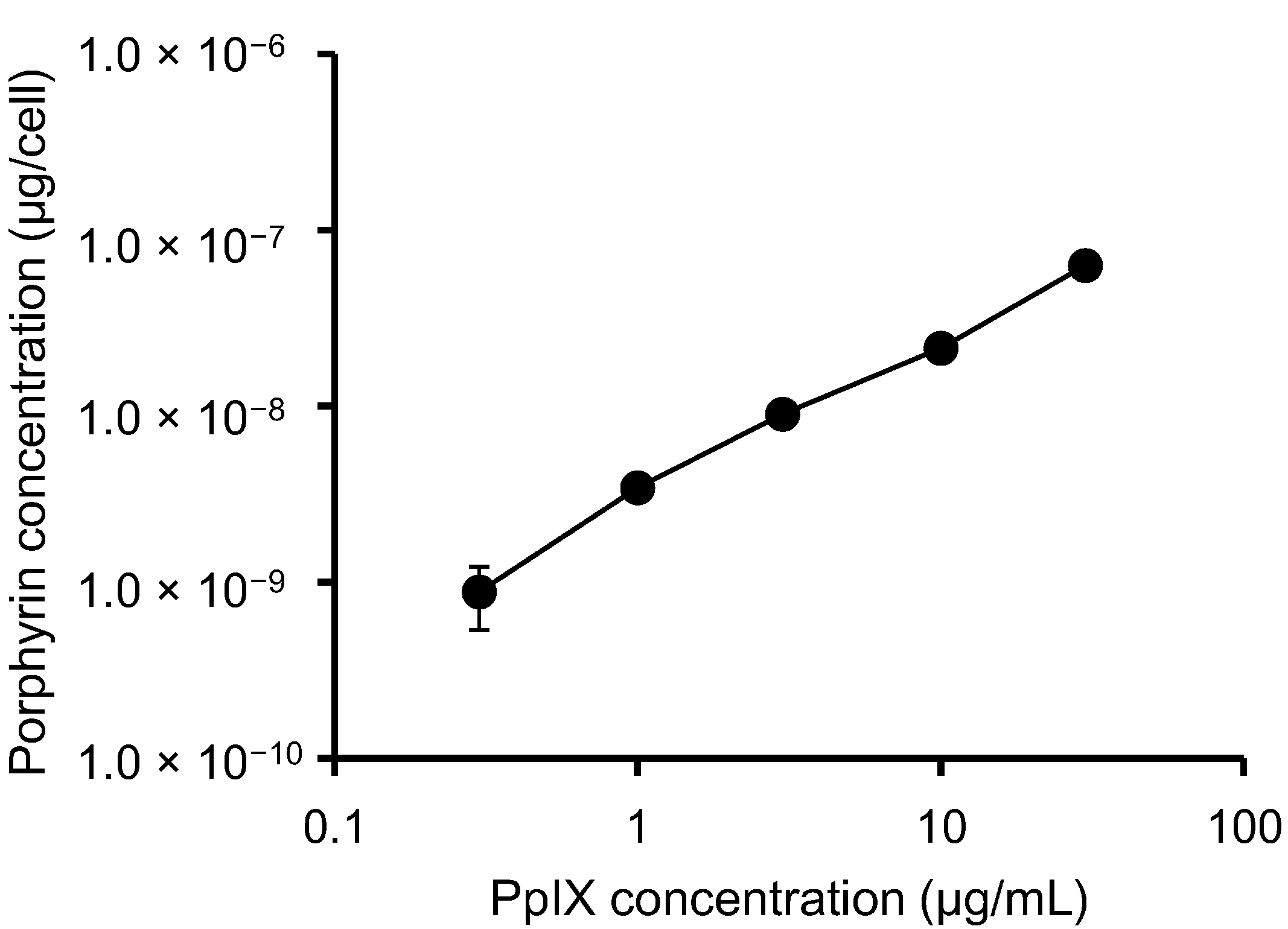
3.1. PpIX Uptake by HeLa Cells
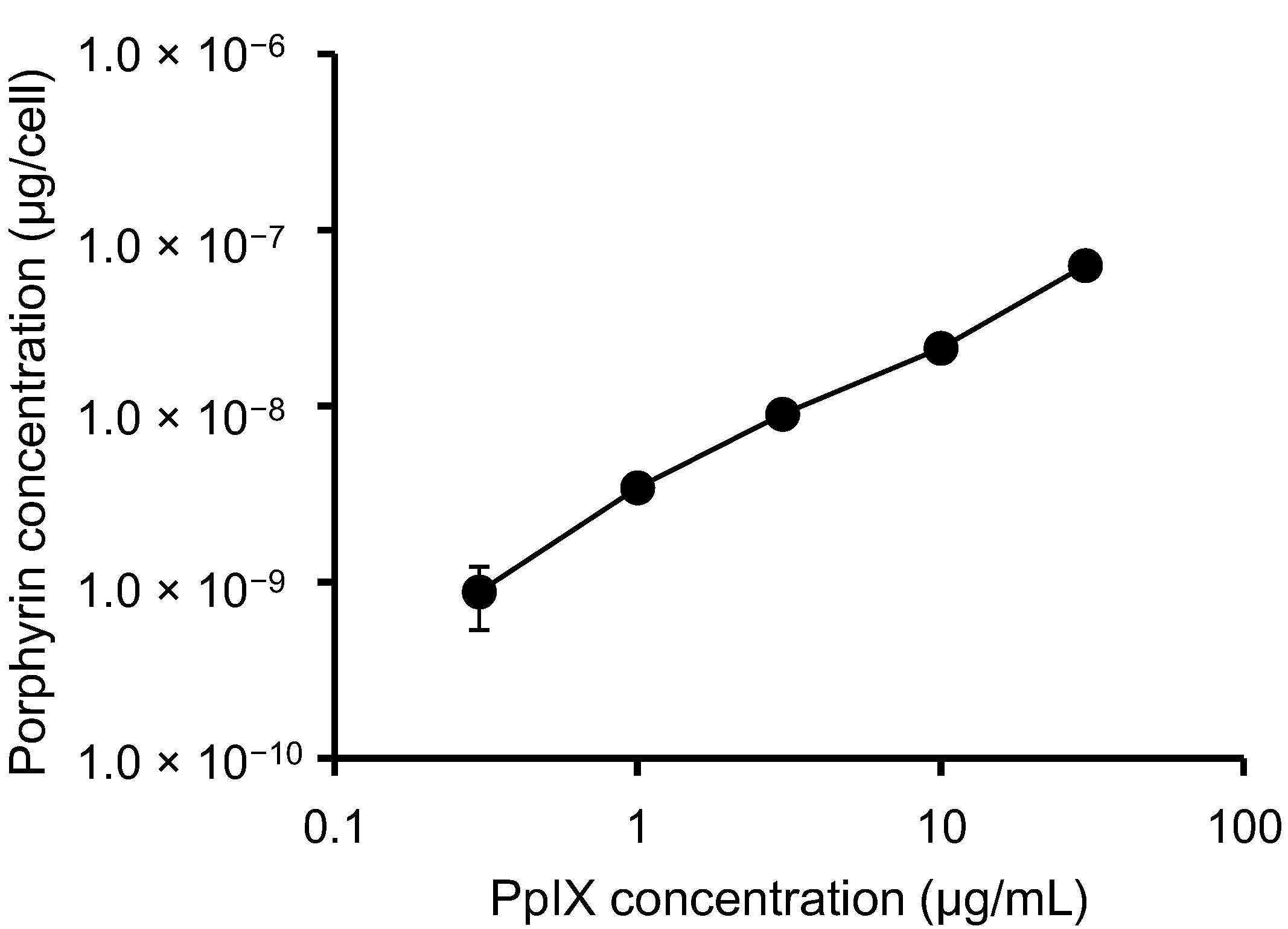
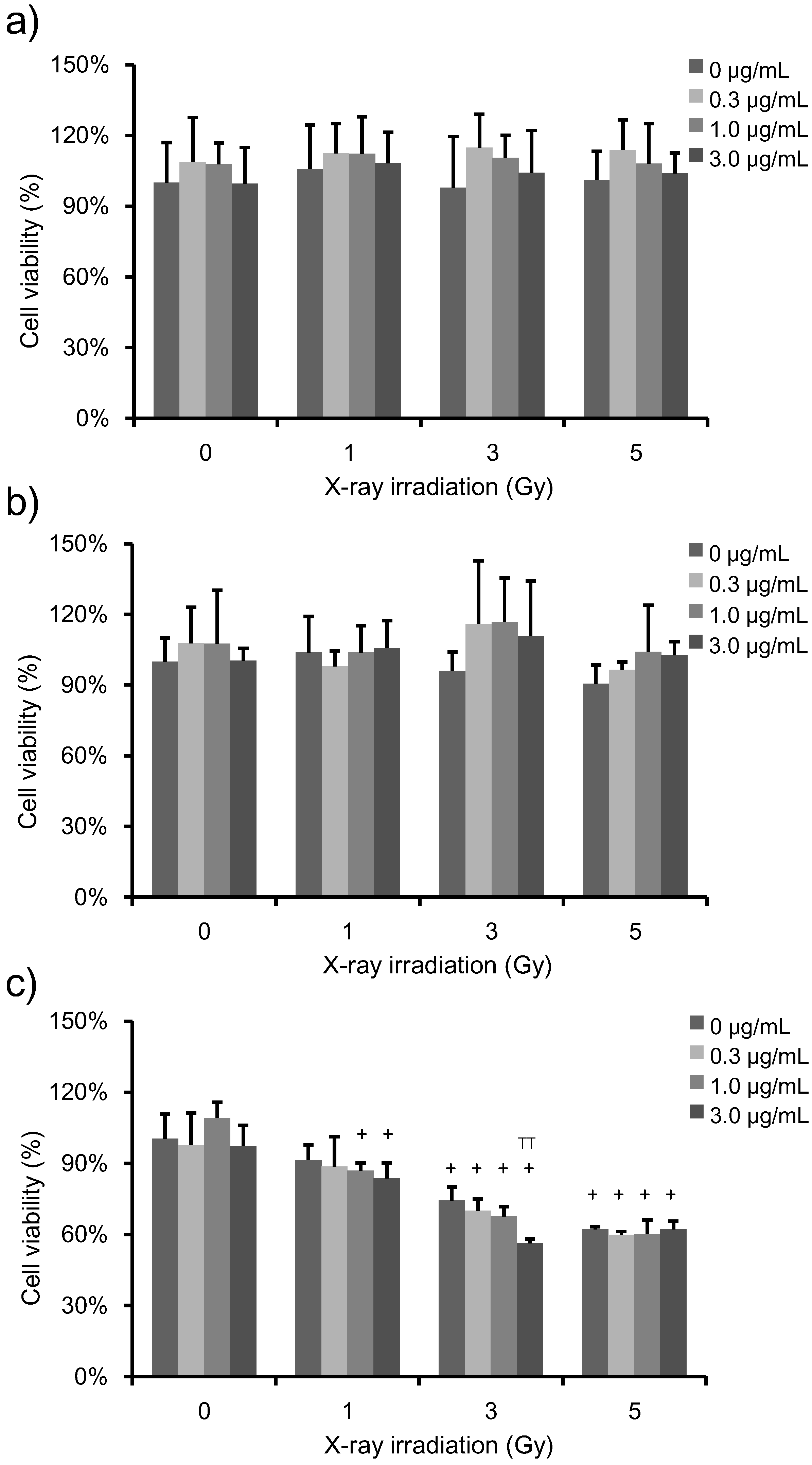
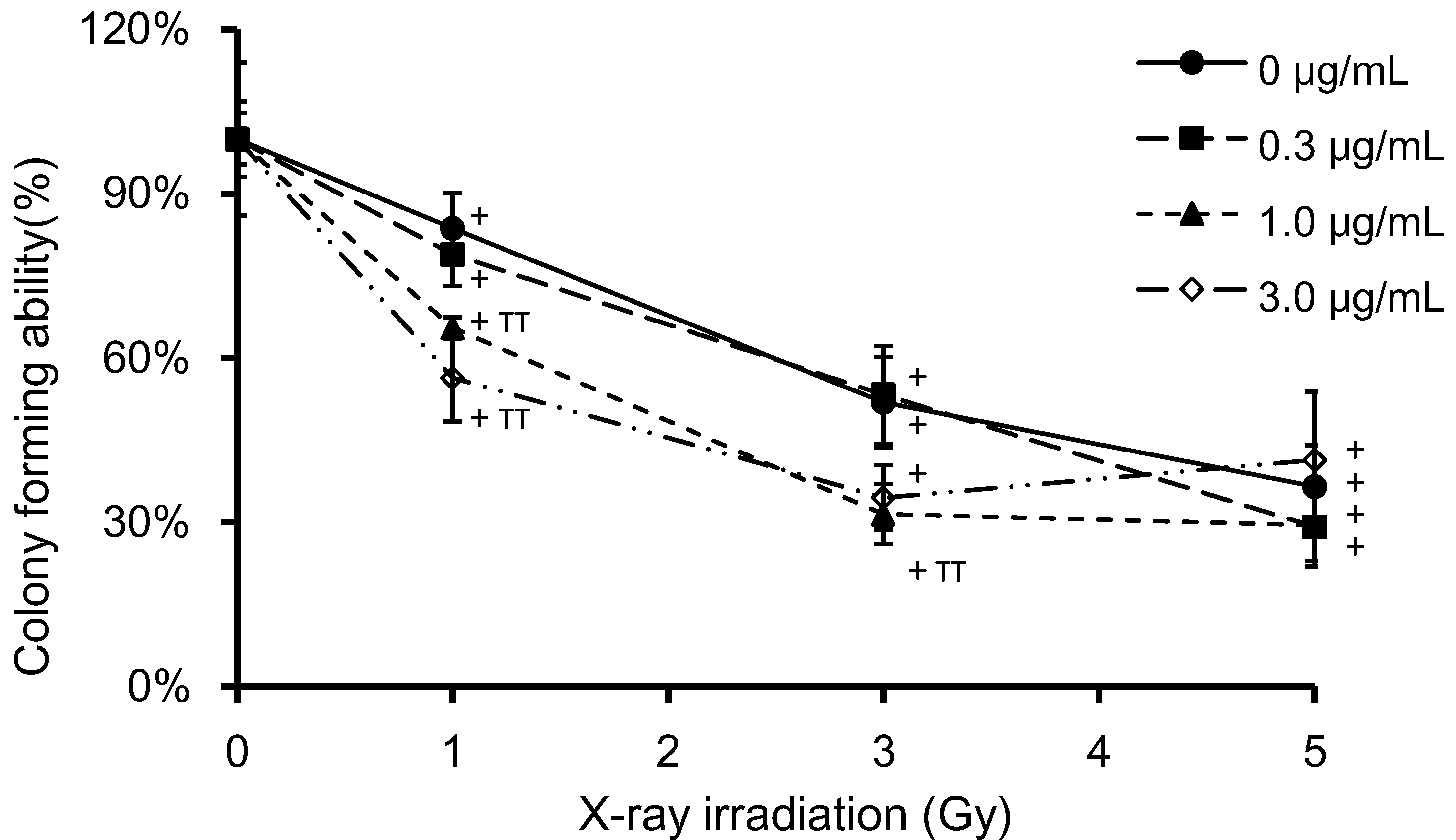
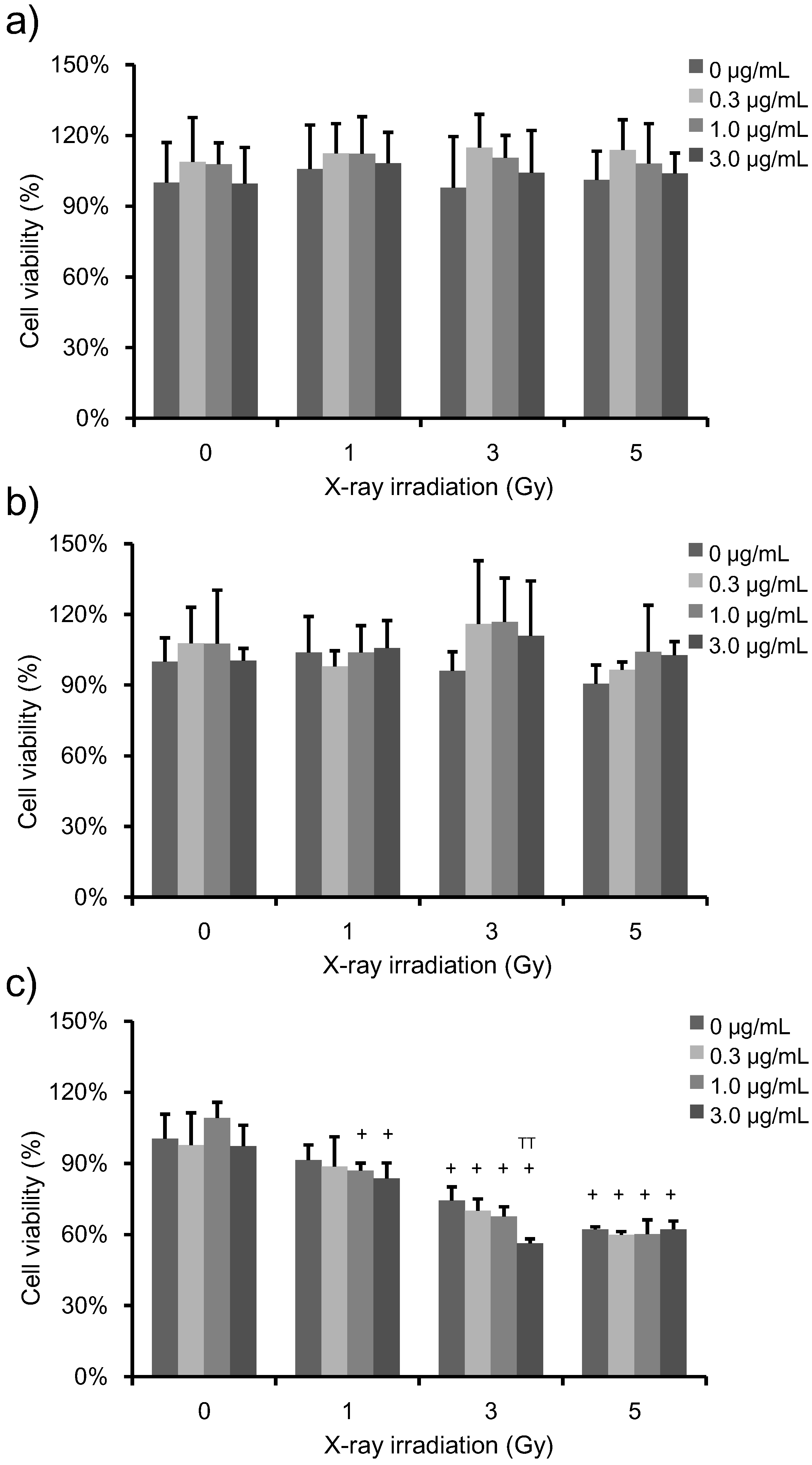
3.2. Determination of Cell Viability and Clonogenic Survival in Irradiated, PpIX-Treated HeLa Cells
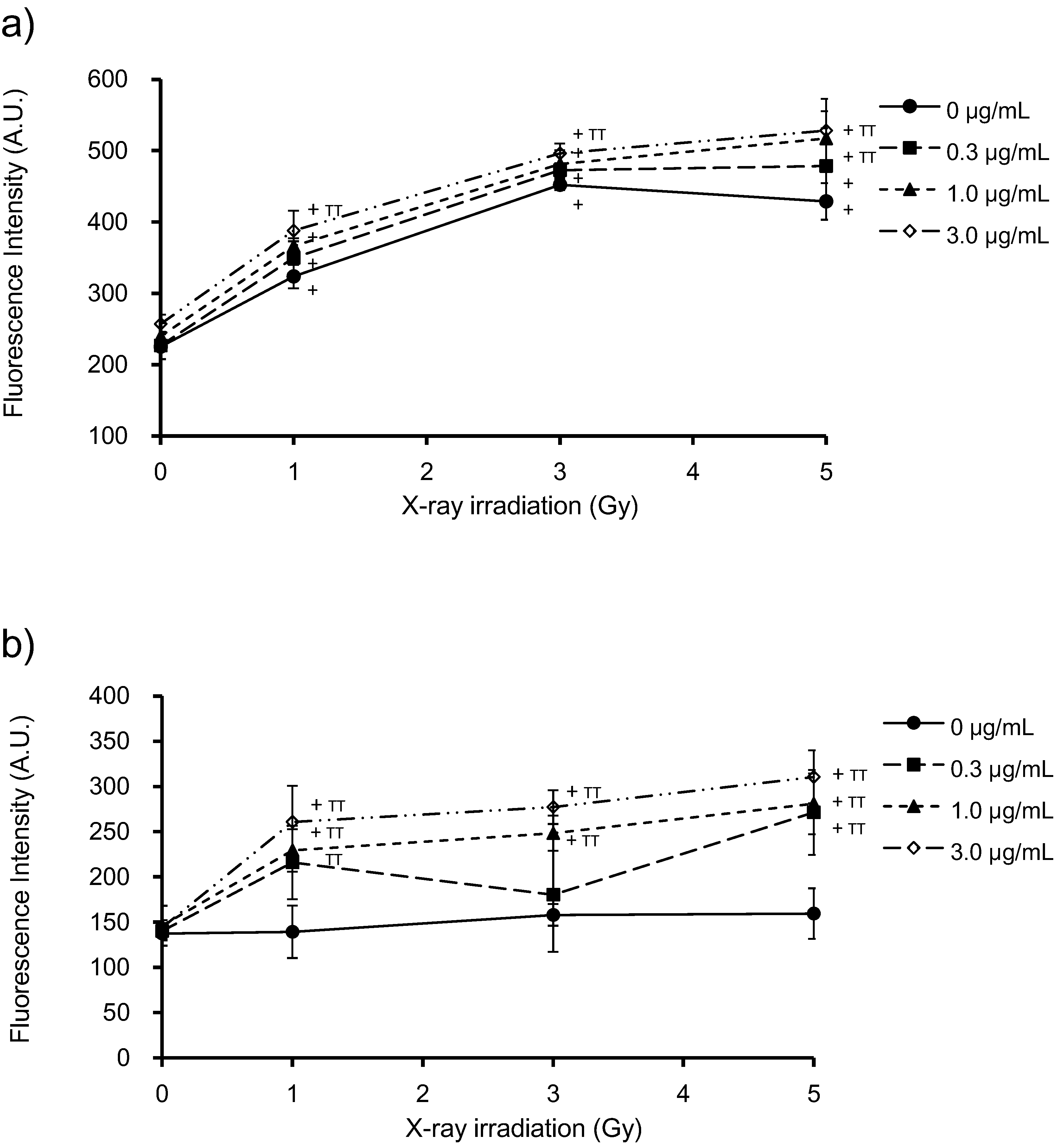
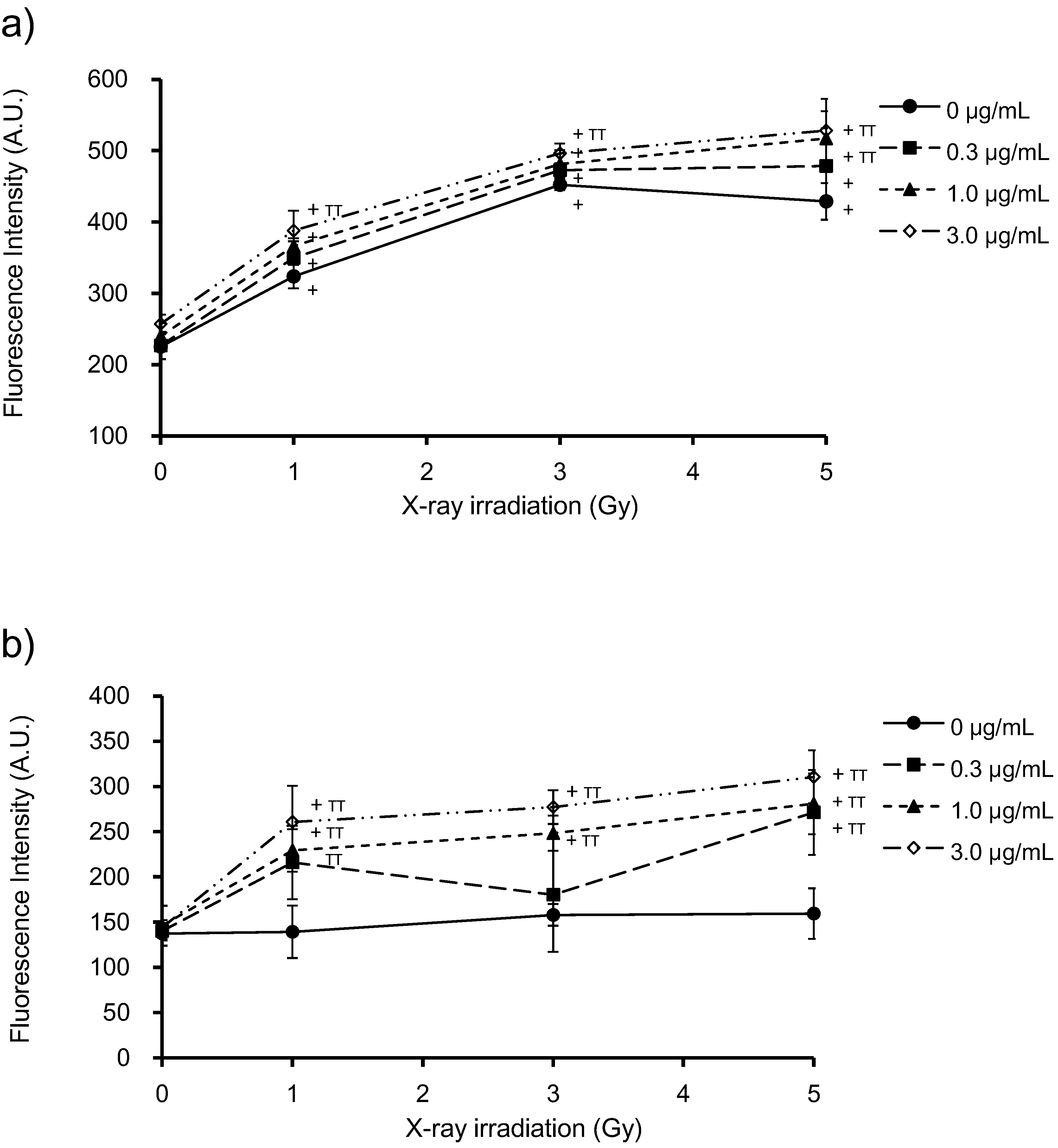
3.3. Generation of ROS by PpIX and X-Ray Irradiation
3.4. Microarray Gene Expression Analysis
3.4.1. Analysis of Microarray Gene Expression Profiles via the Classification of Upregulated and Downregulated Genes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regulation | Number of genes | ||
|---|---|---|---|
| PpIXT | XT | PpIX-XT | |
| Upregulation | 28 | 196 | 290 |
| Downregulation | 39 | 146 | 203 |
| Total | 67 | 342 | 493 |
3.4.2. Functional Analysis
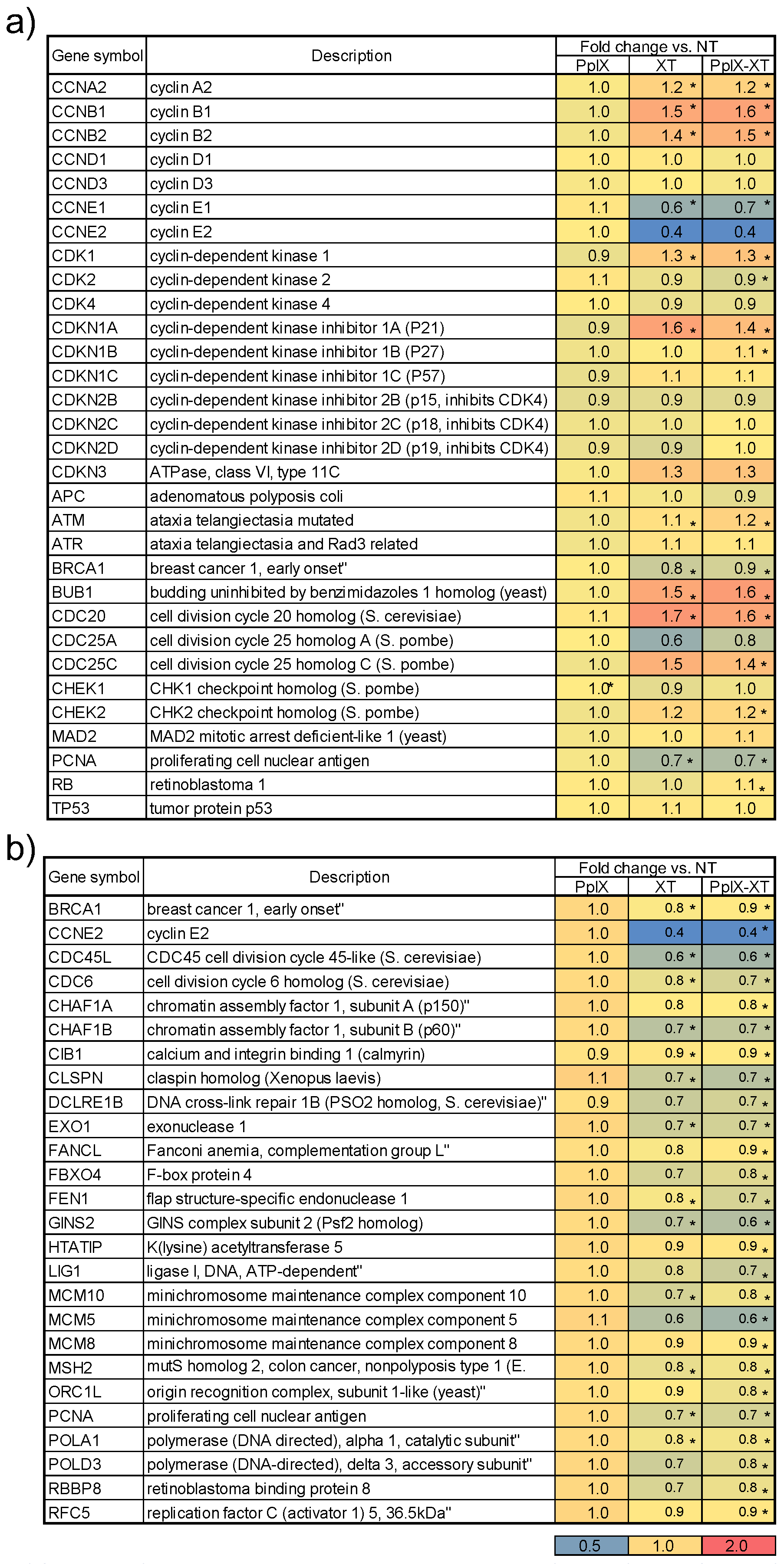
3.5. Functional Validation Using Marker Genes
| Function | Accession | Term | Count | FDR | Gene list of upregulation or downregulation | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| PpIXT | XT | PpIX-XT | PpIXT | XT | PpIX-XT | PpIXT | XT | PpIX-XT | |||
| cell cycle | GO:0000075 | cell cycle checkpoint | 11 | 3.9 × 10−4 | up | ||||||
| GO:0000278 | mitotic cell cycle | 14 | 38 | 4.5 × 10−3 | 9.5 × 10−20 | up | up | ||||
| GO:0007049 | cell cycle | 50 | 2.3 × 10−18 | up | |||||||
| GO:0007093 | mitotic cell cycle checkpoint | 8 | 1.8 × 10−3 | up | |||||||
| GO:0007346 | regulation of mitotic cell cycle | 15 | 1.4 × 10−5 | up | |||||||
| GO:0010564 | regulation of cell cycle process | 11 | 3.2 × 10−3 | up | |||||||
| GO:0022402 | cell cycle process | 18 | 44 | 1.0 × 10−3 | 9.2 × 10−19 | up | up | ||||
| GO:0051726 | regulation of cell cycle | 22 | 2.4 × 10−6 | up | |||||||
| GO:0000280 | nuclear division | 12 | 28 | 9.2 × 10−4 | 5.6 × 10−16 | up | up | ||||
| GO:0007067 | mitotic nuclear division | 12 | 28 | 9.2 × 10−4 | 5.6 × 10−16 | up | up | ||||
| GO:0051301 | cell division | 13 | 29 | 2.4 × 10−3 | 1.8 × 10−13 | up | up | ||||
| GO:0048285 | organelle fission | 12 | 28 | 1.4 × 10−3 | 1.6 × 10−15 | up | up | ||||
| GO:0000087 | M phase of mitotic cell cycle | 12 | 28 | 1.1 × 10−3 | 9.0 × 10−16 | up | up | ||||
| GO:0000279 | M phase | 14 | 34 | 1.2 × 10−3 | 2.5 × 10−17 | ||||||
| GO:0022403 | cell cycle phase | 15 | 39 | 2.8 × 10−3 | 4.9 × 10−19 | Up | up | ||||
| DNA metabolic process | GO:0006259 | DNA metabolic process | 18 | 29 | 7.5 × 10−6 | 3.6 × 10−13 | down | down | |||
| GO:0006260 | DNA replication | 15 | 19 | 7.5 × 10−9 | 1.8 × 10−11 | down | down | ||||
| GO:0006281 | DNA repair | 11 | 18 | 9.9 × 10−3 | 1.7 × 10−7 | down | down | ||||
| GO:0006323 | DNA packaging | 12 | 1.3 × 10−6 | up | |||||||
| GO:0051052 | regulation of DNA metabolic process | 8 | 9 | 7.5 × 10−3 | 5.9 × 10−3 | down | down | ||||
| chromosome organization | GO:0006325 | chromatin organization | 14 | 5.6 × 10−3 | up | ||||||
| GO:0006333 | chromatin assembly or disassembly | 11 | 4.4 × 10−5 | up | |||||||
| GO:0006334 | nucleosome assembly | 11 | 7.6 × 10−7 | up | |||||||
| GO:0031497 | chromatin assembly | 11 | 1.1 × 10−6 | up | |||||||
| GO:0034728 | nucleosome organization | 11 | 2.1 × 10−6 | up | |||||||
| GO:0034622 | cellular macromolecular complex assembly | 15 | 1.2 × 10−4 | up | |||||||
| GO:0043933 | macromolecular complex subunit organization | 19 | 5.2 × 10−3 | up | |||||||
| GO:0051276 | chromosome organization | 19 | 22 | 1.9 × 10−5 | 1.6 × 10−3 | up | up | ||||
| GO:0065003 | macromolecular complex assembly | 19 | 2.1 × 10−3 | up | |||||||
| GO:0065004 | protein-DNA complex assembly | 12 | 8.3 × 10−8 | up | |||||||
| cellular response to stress | GO:0006974 | response to DNA damage stimulus | 18 | 1.1 × 10−5 | down | ||||||
| GO:0033554 | cellular response to stress | 18 | 4.2 × 10−3 | down | |||||||
| other | GO:0007017 | microtubule-based process | 17 | 2.5 × 10−4 | up | ||||||
| GO:0007059 | chromosome segregation | 10 | 1.4 × 10−3 | up | |||||||
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ding, H.; Yu, H.; Dong, Y.; Tian, R.; Huang, G.; Boothman, D.A.; Sumer, B.D.; Gao, J. Photoactivation switch from type II to type I reactions by electron-rich micelles for improved photodynamic therapy of cancer cells under hypoxia. J. Contr. Release 2011, 156, 276–280. [Google Scholar] [CrossRef]
- Pass, H.I. Photodynamic therapy in oncology—Mechanisms and clinical use. J. Natl. Cancer Inst. 1993, 85, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.; Misawa, M. Characterization of reactive oxygen species generated by protoporphyrin IX under X-ray irradiation. Rad. Phys. Chem. 2009, 78, 889–898. [Google Scholar] [CrossRef]
- Peng, Q.; Berg, K.; Moan, J; Kongshaug, M.; Nesland, J.M. 5-Aminolevulinic acid-based photodynamic therapy: Principles and experimental research. Photochem. Photobiol. 1997, 65, 235–251. [Google Scholar]
- Zhao, B.; He, Y.Y. Recent advances in the prevention and treatment of skin cancer using photodynamic therapy. Expert Rev. Anticancer Ther. 2010, 10, 1797–1809. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.; Misawa, M.; Murakami, M.; Mori, T.; Nomura, K.; Iwahashi, H. 5-Aminolevulinic acid enhances cancer radiotherapy in a mouse tumor model. Springerplus 2013, 2, 602. [Google Scholar] [CrossRef] [PubMed]
- Doss, M.; Schmidt, A. Rapid determination of urinary total porphyrins by ion exchange chromatography. Z. Klin. Chem. Klin. Biochem. 1971, 9, 415–418. [Google Scholar] [PubMed]
- Franke, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.; Misawa, M.; Iwahashi, H. Characterization of protoporphytin IX effects on X-ray irradiation in Hela cells in vitro. Available online: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE61805 (accessed on 21 January 2015).
- Agilent Technologies. eArray. Available online: https://earray.chem.agilent.com/earray/ (accessed on 30 October 2014).
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Dennis, G., Jr.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, P3. [Google Scholar] [CrossRef] [PubMed]
- National Institute of Allergy and Infectious Diseases (NIAID), NIH. DAVID Bioinformatics Resources 6.7. Available online: http://david.abcc.ncifcrf.gov/home.jsp (accessed on 30 October 2014).
- Allison, R.R.; Downie, G.H.; Cuenca, R.; Hu, X.H.; Childs, C.J.; Sibata, C.H. Photosensitizers in clinical PDT. Photodiagnosis Photodyn. Ther. 2004, 1, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, C.; Batz, J.; Bolsen, K.; Schulte, K.W.; Zumdick, M.; Ruzicka, T.; Goerz, G. Ex vivo application of delta-aminolevulinic acid induces high and specific porphyrin levels in human skin tumors: Possible basis for selective photodynamic therapy. Photochem. Photobiol. 1997, 66, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Ding, K.K.; Shang, Z.F.; Hao, C.; Xu, Q.Z.; Shen, J.J.; Yang, C.J.; Xie, Y.H.; Qiao, C.; Wang, Y.; Xu, L.L.; et al. Induced expression of the IER5 gene by gamma-ray irradiation and its involvement in cell cycle checkpoint control and survival. Radiat. Environ. Biophys. 2009, 48, 205–213. [Google Scholar]
- Hardmeier, R.; Hoeger, H.; Fang-Kircher, S.; Khoschsorur, A.; Lubec, G. Transcription and activity of antioxidant enzymes after ionizing irradiation in radiation-resistant and radiation-sensitive mice. Proc. Natl. Acad. Sci. USA 1997, 94, 7572–7576. [Google Scholar] [CrossRef] [PubMed]
- Suetens, A.; Moreels, M.; Quintens, R.; Soors, E.; Buset, J.; Chiriotti, S.; Tabury, K.; Gregoire, V.; Baatout, S. Dose- and time-dependent gene expression alterations in prostate and colon cancer cells after in vitro exposure to carbon ion and X-irradiation. J. Radiat. Res. 2015, 56, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Kari, V.; Karpiuk, O.; Tieg, B.; Kriegs, M.; Dikomey, E.; Krebber, H.; Begus-Nahrmann, Y.; Johnsen, S.A. A subset of histone H2B genes produces polyadenylated mRNAs under a variety of cellular conditions. PLoS One 2013, 8, e63745. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, M.A.; Chodosh, L.A.; McKenna, W.G.; Muschel, R.J. Gene expression profile of human cells irradiated in G1 and G2 phases of cell cycle. Cancer Lett. 2003, 195, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Park, W.Y.; Hwang, C.I.; Im, C.N.; Kang, M.J.; Woo, J.H.; Kim, J.H.; Kim, Y.S.; Kim, J.H.; Kim, H.; Kim, K.A.; et al. Identification of radiation-specific responses from gene expression profile. Oncogene 2003, 21, 8521–8528. [Google Scholar]
- Kis, E.; Szatmári, T.; Keszei, M.; Farkas, R.; Esik, O.; Lumniczky, K.; Falus, A.; Sáfrány, G. Microarray analysis of radiation response genes in primary human fibroblasts. Int. J. Radiat. Oncol. Biol. Phys. 2006, 66, 1506–1514. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.S.; Bonner, W.M. Separation of basal histone synthesis from S-phase histone synthesis in dividing cells. Cell 1981, 27, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.M.; Ye, X.; Hall, C.; Santos, H.; Ma, T.; Kao, G.D.; Yen, T.J.; Harper, J.W.; Adams, P.D. Coupling of DNA Synthesis and Histone Synthesis in S Phase Independent of Cyclin/cdk2 Activity. Mol. Cell Biol. 2002, 22, 7459–7472. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takahashi, J.; Misawa, M.; Iwahashi, H. Transcriptome Analysis of Porphyrin-Accumulated and X-Ray-Irradiated Cell Cultures under Limited Proliferation and Non-Lethal Conditions. Microarrays 2015, 4, 25-40. https://doi.org/10.3390/microarrays4010025
Takahashi J, Misawa M, Iwahashi H. Transcriptome Analysis of Porphyrin-Accumulated and X-Ray-Irradiated Cell Cultures under Limited Proliferation and Non-Lethal Conditions. Microarrays. 2015; 4(1):25-40. https://doi.org/10.3390/microarrays4010025
Chicago/Turabian StyleTakahashi, Junko, Masaki Misawa, and Hitoshi Iwahashi. 2015. "Transcriptome Analysis of Porphyrin-Accumulated and X-Ray-Irradiated Cell Cultures under Limited Proliferation and Non-Lethal Conditions" Microarrays 4, no. 1: 25-40. https://doi.org/10.3390/microarrays4010025
APA StyleTakahashi, J., Misawa, M., & Iwahashi, H. (2015). Transcriptome Analysis of Porphyrin-Accumulated and X-Ray-Irradiated Cell Cultures under Limited Proliferation and Non-Lethal Conditions. Microarrays, 4(1), 25-40. https://doi.org/10.3390/microarrays4010025