Parkinsonisms and Glucocerebrosidase Deficiency: A Comprehensive Review for Molecular and Cellular Mechanism of Glucocerebrosidase Deficiency

Abstract
:1. Introduction
2. Methods
2.1. Search Strategy and Selection Criteria
2.2. Lysosomal Diseases and Gaucher Epidemiology
2.3. Phenotype/Genotype: Clinical Features
2.4. Biomarkers
2.4.1. Wet Biomarkers in GBA Mutation Carriers PD
2.4.2. Prodromal Signs in PD-GBA Patients
2.4.3. Cognition in GBA Homozygous and Heterozygous GBA Mutations Carriers
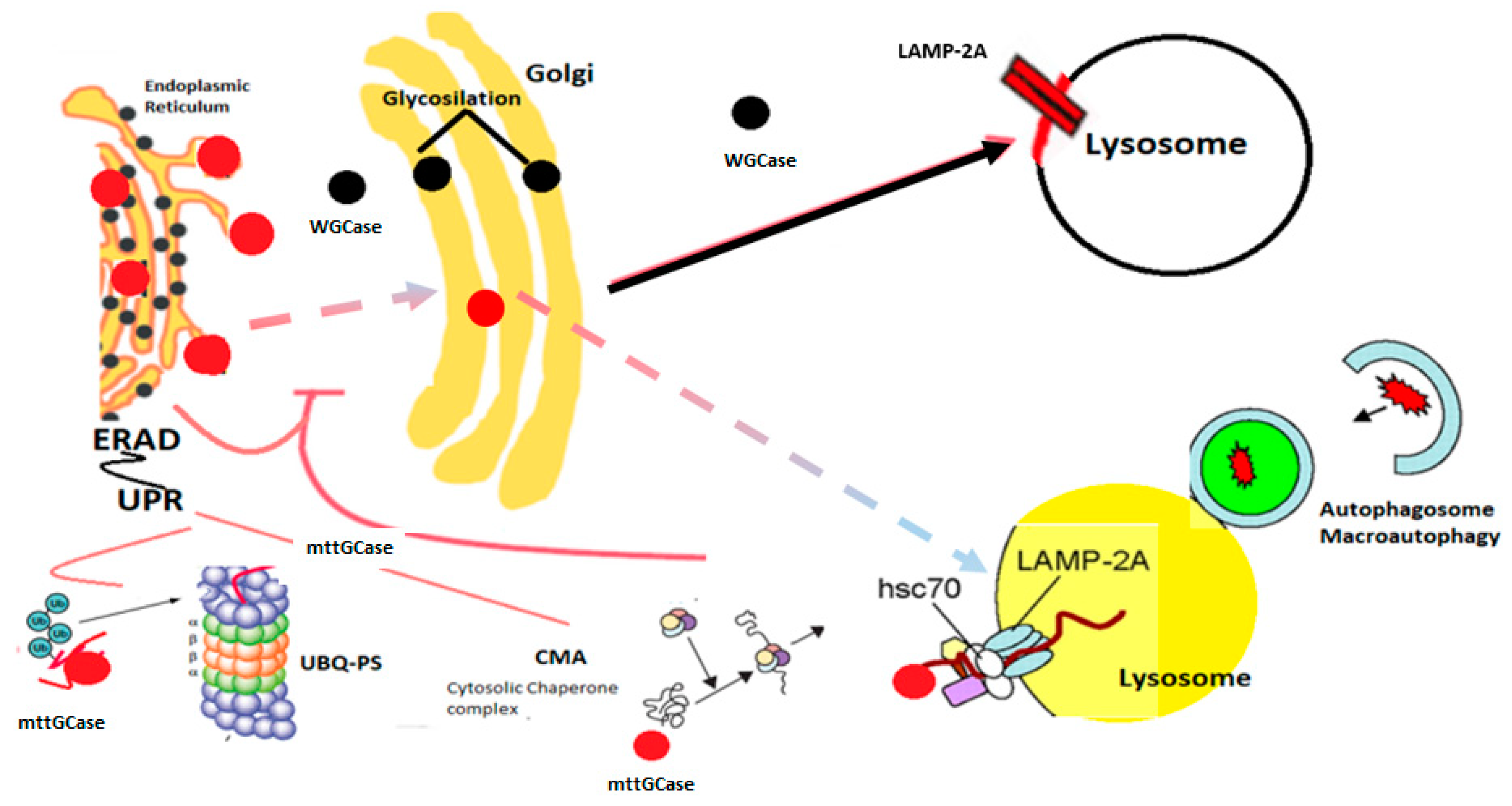
2.4.4. The Role of Autophagy in Lysosomal Diseases and Neurodegeneration (α-synuclein)
2.4.5. GBA Gene Mutations
2.5. Gaucher and PD: the Ethiopatogenic Link
2.6. α-Synuclein and GCase Link
2.7. Parkin-Pink1 Mitochondria and GCase
2.8. Therapeutic Implications
2.9. Future Therapies for GD
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moors, T.E.; Paciotti, S.; Ingrassia, A.; Quadri, M.; Breedveld, G.; Tasegian, A.; Chiasserini, D.; Eusebi, P.; Duran-Pacheco, G.; Kremer, T.; et al. Characterization of Brain Lysosomal Activities in GBA-Related and Sporadic Parkinson’s Disease and Dementia with Lewy Bodies. Mol. Neurobiol. 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 2015, 12, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Thibaudeau, T.A.; Anderson, R.T.; Smith, D.M. A common mechanism of proteasome impairment by neurodegenerative disease-associated oligomers. Nat. Commun. 2018, 9, 1097. [Google Scholar] [CrossRef] [PubMed]
- Balestrino, R.; Schapira, A. Glucocerebrosidase and Parkinson Disease: Molecular, Clinical and Therapeutic Implications. Neuroscientist 2018, 5, 540–559. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Le, W. Milestones of Parkinson’s Disease Research: 200 Years of History and Beyond. Neurosci. Bull. 2017, 33, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Alcalay, R.N. Genetic Forms of Parkinson’s Disease. Semin. Neurol. 2017, 37, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Domingo, A.; Klein, C. Genetics of Parkinson disease. Handb. Clin. Neurol. 2018, 147, 211–227. [Google Scholar] [CrossRef] [PubMed]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef]
- Xilouri, M.; Brekk, O.R.; Stefanis, L. Alpha-synuclein and protein degradation systems: A reciprocal relationship. Mol. Neurobiol. 2013, 47, 537–551. [Google Scholar] [CrossRef]
- Kett, L.R.; Boassa, D.; Ho, C.C.; Rideout, H.J.; Hu, J.; Terada, M.; Ellisman, M.; Dauer, W.T. LRRK2 Parkinson disease mutations enhance its microtubule association. Hum. Mol. Genet. 2012, 21, 890–899. [Google Scholar] [CrossRef]
- Harbour, M.E.; Breusegem, S.Y.A.; Antrobus, R.; Freeman, C.; Reid, E.; Seaman, M.N.J. The cargo-selective retromer complex is a recruiting hub for protein complexes that regulate endosomal tubule dynamics. J. Cell Sci. 2010, 123, 3703–3717. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Dodson, M.W.; Huang, H.; Guo, M. The Parkinson’s disease genes pink1 and parkin promote mitochondrial fission and/or inhibit fusion in Drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 14503–14508. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.R.; Gahl, W.A. Lysosomal storage diseases. Transl. Sci. Rare Dis. 2017, 2, 1–71. [Google Scholar] [CrossRef] [PubMed]
- Kielian, T. Lysosomal storage disorders: Pathology within the lysosome and beyond. J. Neurochem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kinghorn, K.J.; Asghari, A.M.; Castillo-Quan, J.I. The emerging role of autophagic-lysosomal dysfunction in Gaucher Disease and Parkinson’s disease. Neural Regen. Res. 2017, 12, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Thaler, A.; Bregman, N.; Gurevich, T.; Shiner, T.; Dror, Y.; Zmira, O.; Gan-Or, Z.; Bar-Shira, A.; Gana-Weisz, M.; Orr-Urtreger, A.; et al. Parkinson’s disease phenotype is influenced by the severity of the mutations in the GBA gene. Parkinsonism Relat. Disord. 2018, 55, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Beutler, E.; Gelbart, T. Mutation analysis in Gaucher disease. Am. J. Med. Genet. 1992, 44, 389–390. [Google Scholar] [CrossRef] [PubMed]
- Reczek, D.; Schwake, M.; Schröder, J.; Hughes, H.; Blanz, J.; Jin, X.; Brondyk, W.; Van Patten, S.; Edmunds, T.; Saftig, P. LIMP-2 is a receptor for lysosomal mannose-6-phosphate-independent targeting of beta-glucocerebrosidase. Cell 2007, 131, 770–783. [Google Scholar] [CrossRef]
- Jović, M.; Kean, M.J.; Szentpetery, Z.; Polevoy, G.; Gingras, A.C.; Brill, J.A.; Balla, T. Two phosphatidylinositol 4-kinases control lysosomal delivery of the Gaucher disease enzyme, β-glucocerebrosidase. Mol. Biol. Cell 2012, 23, 1533–1545. [Google Scholar] [CrossRef]
- Dandana, A.; Ben Khelifa, S.; Chahed, H.; Miled, A.; Ferchichi, S. Gaucher Disease: Clinical, Biological and Therapeutic Aspects. Pathobiology 2016, 83, 13–23. [Google Scholar] [CrossRef]
- Rodriguez-Porcel, F.; Espay, A.J.; Carecchio, M. Parkinson disease in Gaucher disease. J. Clin. Mov. Disord. 2017, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Siebert, M.; Sidransky, E.; Westbroek, W. Glucocerebrosidase is shaking up the synucleinopathies. Brain 2014, 137, 1304–1322. [Google Scholar] [CrossRef] [PubMed]
- Capablo Liesa, J.L.; de Cabezón, A.S.; Alarcia Alejos, R.; Ara Callizo, J.R. Clinical characteristics of the neurological forms of Gaucher’s disease. Med. Clin. 2011, 137 (Suppl. 1), 6–11. [Google Scholar] [CrossRef]
- Jinnah, H.A.; Albanese, A.; Bhatia, K.P.; Cardoso, F.; Da Prat, G.; de Koning, T.J.; Espay, A.J.; Fung, V.; Garcia-Ruiz, P.J.; Gershanik, O.; et al. International Parkinson’s Disease Movement Disorders Society Task Force on Rare Movement Disorders. Treatable inherited rare movement disorders. Mov. Disord. 2018, 33, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Drelichman, G.; Fernández Escobar, N.; Basack, N.; Aversa, L.; Kohan, R.; Watman, N.; Bolesina, M.; Elena, G.; Veber, S.E.; Dragosky, M.; et al. Enfermedad de Gaucher en Argentina Un informe del Registro Internacional de Gaucher y del Grupo Argentino de Diagnóstico y Tratamiento de la Enfermedad de Gaucher 12Revista de Hematologia; 17-Suplemento Enfermedad de Gaucher; Sociedad Argentina de Hematología HEMATOLOGÍA: Buenos Aires, Argentina, 2013; pp. 4–16. [Google Scholar]
- Ebrahimi-Fakhari, D.; Hildebrandt, C.; Davis, P.E.; Rodan, L.H.; Anselm, I.; Bodamer, O. The Spectrum of Movement Disorders in Childhood-Onset Lysosomal Storage Diseases. Mov. Disord. Clin. Pract. 2018, 5, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Sidransky, E.; Nalls, M.A.; Aasly, J.O.; Aharon-Peretz, J.; Annesi, G.; Barbosa, E.R.; Bar-Shira, A.; Berg, D.; Bras, J.; Brice, A.; et al. Multicenter analysis of glucocerebrosidase mutations in Parkinson’s disease. N. Engl. J. Med. 2009, 361, 1651–1661. [Google Scholar] [CrossRef] [PubMed]
- Velayati, A.; Yu, W.H.; Sidransky, E. The role of glucocerebrosidase mutations in Parkinson disease and Lewy body disorders. Curr. Neurol. Neurosci. Rep. 2010, 10, 190–198. [Google Scholar] [CrossRef]
- Gan-Or, Z.; Liong, C.; Alcalay, R.N. GBA-Associated Parkinson’s Disease and Other Synucleinopathies. Curr. Neurol. Neurosci. Rep. 2018, 18, 44. [Google Scholar] [CrossRef]
- Mitsui, J.; Mizuta, I.; Toyoda, A.; Ashida, R.; Takahashi, Y.; Goto, J.; Fukuda, Y.; Date, H.; Iwata, A.; Yamamoto, M.; et al. Mutations for Gaucher disease confer high susceptibility to Parkinson disease. Arch. Neurol. 2009, 66, 571–576. [Google Scholar] [CrossRef]
- Alcalay, R.N.; Dinur, T.; Quinn, T.; Sakanaka, K.; Levy, O.; Waters, C.; Fahn, S.; Dorovski, T.; Chung, W.K.; Pauciulo, M.; et al. Comparison of Parkinson risk in Ashkenazi Jewish patients with Gaucher disease and GBA heterozygotes. JAMA Neurol. 2014, 71, 752–757. [Google Scholar] [CrossRef]
- Grabowski, G.A.; Zimran, A.; Ida, H. Gaucher disease types 1 and 3: Phenotypic characterization of large populations from the ICGG Gaucher Registry. Am. J. Hematol. 2015, 90, 12–18. [Google Scholar] [CrossRef]
- Spitz, M.; Rozenberg, R.; Pereira Lda, V.; Reis Barbosa, E. Association between Parkinson’s disease and glucocerebrosidase mutations in Brazil. Parkinsonism Relat. Disord. 2008, 14, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Bultron, G.; Kacena, K.; Pearson, D.; Boxer, M.; Yang, R.; Sathe, S.; Pastores, G.; Mistry, P.K. The risk of Parkinson’s disease in type 1 Gaucher disease. J. Inherit. Metab. Dis. 2010, 33, 167–173. [Google Scholar] [CrossRef] [PubMed]
- McNeill, A.; Duran, R.; Proukakis, C.; Bras, J.; Hughes, D.; Mehta, A.; Hardy, J.; Wood, N.W.; Schapira, A.H. Hyposmia and cognitive impairment in Gaucher disease patients and carriers. Mov. Disord. 2012, 27, 526–532. [Google Scholar] [CrossRef]
- Mao, X.; Wang, T.; Peng, R.; Chang, X.; Li, N.; Gu, Y.; Zhao, D.; Liao, Q.; Liu, M. Mutations in GBA and risk of Parkinson’s disease: A meta-analysis based on 25 case-control studies. Neurol. Res. 2013, 35, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Blanz, J.; Saftig, P. Parkinson’s disease: Acid-glucocerebrosidase activity and alpha-synuclein clearance. J. Neurochem. 2016, 139 (Suppl. 1), 198–215. [Google Scholar] [CrossRef] [PubMed]
- Migdalska-Richards, A.; Schapira, A.H. The relationship between glucocerebrosidase mutations and Parkinson disease. J. Neurochem. 2016, 139 (Suppl. 1), 77–90. [Google Scholar] [CrossRef]
- Aflaki, E.; Westbroek, W.; Sidransky, E. The Complicated Relationship between Gaucher Disease and Parkinsonism: Insights from a Rare Disease. Neuron 2017, 93, 737–746. [Google Scholar] [CrossRef]
- O’Regan, G.; deSouza, R.M.; Balestrino, R.; Schapira, A.H. Glucocerebrosidase Mutations in Parkinson Disease. J. Parkinsons Dis. 2017, 7, 411–422. [Google Scholar] [CrossRef]
- Standaert, D.G. What would Dr. James parkinson think today? Mutations in beta-glucocerebrosidase and risk of Parkinson’s disease. Mov. Disord. 2017, 32, 1341–1342. [Google Scholar] [CrossRef]
- Giraldo, P.; Capablo, J.L.; Alfonso, P.; Garcia-Rodriguez, B.; Latre, P.; Irun, P.; de Cabezon, A.S.; Pocovi, M. Neurological manifestations in patients with Gaucher disease and their relatives, it is just a coincidence? J. Inherit. Metab. Dis. 2011, 34, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Setó-Salvia, N.; Pagonabarraga, J.; Houlden, H.; Pascual-Sedano, B.; Dols-Icardo, O.; Tucci, A.; Paisán-Ruiz, C.; Campolongo, A.; Antón-Aguirre, S.; Martín, I.; et al. Glucocerebrosidase mutations confer a greater risk of dementia during Parkinson’s disease course. Mov. Disord. 2012, 27, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Grabowski, G.A. Impaired autophagosomes and lysosomes in neuronopathic Gaucher disease. Autophagy 2010, 6, 648–649. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.W.; Ye, J.; Qiu, W.J.; Han, L.S.; Gu, X.F. Comparison and clinical application of two methods for determination of plasma chitotriosidase activity. Zhonghua Er Ke Za Zhi 2012, 50, 834–838. (In Chinese) [Google Scholar]
- Choi, J.M.; Kim, W.C.; Lyoo, C.H.; Kang, S.Y.; Lee, P.H.; Baik, J.S.; Koh, S.B.; Ma, H.I.; Sohn, Y.H.; Lee, M.S.; et al. Association of mutations in the glucocerebrosidase gene with Parkinson disease in a Korean population. Neurosci. Lett. 2012, 514, 12–15. [Google Scholar] [CrossRef]
- Biegstraaten, M.; Wesnes, K.A.; Luzy, C.; Petakov, M.; Mrsic, M.; Niederau, C.; Giraldo, P.; Hughes, D.; Mehta, A.; Mengel, K.E.; et al. The cognitive profile of type 1 Gaucher disease patients. J. Inherit. Metab. Dis. 2012, 35, 1093–1099. [Google Scholar] [CrossRef] [PubMed]
- Schapira, A.H. Glucocerebrosidase and Parkinson disease: Recent advances. Mol. Cell Neurosci. 2015, 66, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Barrett, M.J.; Hagenah, J.; Dhawan, V.; Peng, S.; Stanley, K.; Raymond, D.; Deik, A.; Gross, S.J.; Schreiber-Agus, N.; Mirelman, A.; et al. LRRK2 Ashkenazi Jewish Consortium. Transcranial sonography and functional imaging in glucocerebrosidase mutation Parkinson disease. Parkinsonism Relat. Disord. 2013, 19, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Saunders-Pullman, R.; Hagenah, J.; Dhawan, V.; Stanley, K.; Pastores, G.; Sathe, S.; Tagliati, M.; Condefer, K.; Palmese, C.; Brüggemann, N.; et al. Gaucher disease ascertained through a Parkinson’s center: Imaging and clinical characterization. Mov. Disord. 2010, 25, 1364–1372. [Google Scholar] [CrossRef]
- Goker-Alpan, O.; Masdeu, J.C.; Kohn, P.D.; Ianni, A.; Lopez, G.; Groden, C.; Chapman, M.C.; Cropp, B.; Eisenberg, D.P.; Maniwang, E.D.; et al. The neurobiology of glucocerebrosidase-associated parkinsonism: A positron emission tomography study of dopamine synthesis and regional cerebral blood flow. Brain 2012, 135, 2440–2448. [Google Scholar] [CrossRef]
- Agosta, F.; Sarasso, E.; Filippi, M. Functional MRI in Atypical Parkinsonisms. Int. Rev. Neurobiol. 2018, 142, 149–173. [Google Scholar] [CrossRef] [PubMed]
- Kraoua, I.; Stirnemann, J.; Ribeiro, M.J.; Rouaud, T.; Verin, M.; Annic, A.; Rose, C.; Defebvre, L.; Réménieras, L.; Schüpbach, M.; et al. Parkinsonism in Gaucher’s disease type 1: Ten new cases and a review of the literature. Mov. Disord. 2009, 24, 1524–1530. [Google Scholar] [CrossRef] [PubMed]
- Parnetti, L.; Paciotti, S.; Eusebi, P.; Dardis, A.; Zampieri, S.; Chiasserini, D.; Tasegian, A.; Tambasco, N.; Bembi, B.; Calabresi, P.; et al. Cerebrospinal fluid β-glucocerebrosidase activity is reduced in parkinson’s disease patients. Mov. Disord. 2017, 32, 1423–1431. [Google Scholar] [CrossRef] [PubMed]
- Parnetti, L.; Chiasserini, D.; Persichetti, E.; Eusebi, P.; Varghese, S.; Qureshi, M.M.; Dardis, A.; Deganuto, M.; De Carlo, C.; Castrioto, A.; et al. Cerebrospinal fluid lysosomal enzymes and alpha-synuclein in Parkinson’s disease. Mov. Disord. 2014, 29, 1019–1027. [Google Scholar] [CrossRef] [PubMed]
- Alcalay, R.N.; Levy, O.A.; Waters, C.C.; Fahn, S.; Ford, B.; Kuo, S.-H.; Mazzoni, P.; Pauciulo, M.W.; Nichols, W.C.; Gan-Or, Z.; et al. Glucocerebrosidase activity in Parkinson’s disease with and without GBA mutations. Brain 2015, 138, 2648–2658. [Google Scholar] [CrossRef]
- Gatto, E.M.; Etcheverry, J.L.; Sanguinetti, A.; Cesarini, M.; Fernandez Escobar, N.; Drelichman, G. Prodromal Clinical Markers of Parkinson disease in Gaucher Disease Individuals. Eur. Neurol. 2016, 76, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Litvan, I.; Goldman, J.G.; Tröster, A.I.; Schmand, B.A.; Weintraub, D.; Petersen, R.C.; Mollenhauer, B.; Adler, C.H.; Marder, K.; Williams-Gray, C.H.; et al. Diagnostic criteria for mild cognitive impairment in Parkinson’s disease: Movement Disorder Society Task Force guidelines. Mov. Disord. 2012, 27, 349–356. [Google Scholar] [CrossRef]
- Liu, G.; Locascio, J.J.; Corvol, J.C.; Boot, B.; Liao, Z.; Page, K.; Franco, D.; Burke, K.; Jansen, I.E.; Trisini-Lipsanopoulos, A.; et al. Prediction of cognition in Parkinson’s disease with a clinical-genetic score: A longitudinal analysis of nine cohorts. Lancet Neurol. 2017, 16, 620–629. [Google Scholar] [CrossRef]
- Mata, I.F.; Leverenz, J.B.; Weintraub, D.; Trojanowski, J.Q.; Chen-Plotkin, A.; Van Deerlin, V.M.; Ritz, B.; Rausch, R.; Factor, S.A.; Wood-Siverio, C.; et al. GBA Variants are associated with a distinct pattern of cognitive deficits in Parkinson’s disease. Mov. Disord. 2016, 31, 95–102. [Google Scholar] [CrossRef]
- Bregman, N.; Thaler, A.; Mirelman, A.; Helmich, R.C.; Gurevich, T.; Orr-Urtreger, A.; Marder, K.; Bressman, S.; Bloem, B.R.; Giladi, N. LRRK2 Ashkenazi Jewish consortium. A cognitive fMRI study in non-manifesting LRRK2 and GBA carriers. Brain Struct. Funct. 2017, 222, 1207–1218. [Google Scholar] [CrossRef]
- Chahine, L.M.; Urbe, L.; Caspell-Garcia, C.; Aarsland, D.; Alcalay, R.; Barone, P.; Burn, D.; Espay, A.J.; Hamilton, J.L.; Hawkins, K.A.; et al. Parkinson’s Progression Markers Initiative. Cognition among individuals along a spectrum of increased risk for Parkinson’s disease. PLoS ONE 2018, 20, e0201964. [Google Scholar] [CrossRef]
- Taguchi, Y.V.; Liu, J.; Ruan, J.; Pacheco, J.; Zhang, X.; Abbasi, J.; Keutzer, J.; Mistry, P.K.; Chandra, S.S. Glucosylsphingosine Promotes α-Synuclein Pathology in MutantGBA-Associated Parkinson’s Disease. J. Neurosci. 2017, 37, 9617–9631. [Google Scholar] [CrossRef] [PubMed]
- Chun, Y.; Kim, J. Autophagy: An Essential Degradation Program for Cellular Homeostasis and Life. Cells 2018, 7, 278. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Kroemer, G. Biological Functions of Autophagy Genes: A Disease Perspective. Cell 2019, 176, 11–42. [Google Scholar] [CrossRef] [PubMed]
- Bengoa-Vergniory, N.; Roberts, R.F.; Wade-Martins, R.; Alegre-Abarrategui, J. Alpha-synuclein oligomers: A new hope. Acta Neuropathol. 2017, 134, 819–838. [Google Scholar] [CrossRef] [PubMed]
- Mor, D.E.; Daniels, M.J.; Ischiropoulos, H. The usual suspects, dopamine and alpha-synuclein, conspire to cause neurodegeneration. Mov. Disord. 2019. [Google Scholar] [CrossRef]
- Blandini, F.; Cilia, R.; Cerri, S.; Pezzoli, G.; Schapira, A.H.V.; Mullin, S.; Lanciego, J.L. Glucocerebrosidase mutations and synucleinopathies: Toward a model of precision medicine. Mov. Disord. 2018. [Google Scholar] [CrossRef]
- Ruskey, J.A.; Greenbaum, L.; Roncière, L.; Alam, A.; Spiegelman, D.; Liong, C.; Levy, O.A.; Waters, C.; Fahn, S.; Marder, K.S.; et al. Increased yield of full GBA sequencing in Ashkenazi Jews with Parkinson’s disease. Eur. J. Med. Genet. 2018, S1769–S7212. [Google Scholar] [CrossRef]
- Stirnemann, J.; Vigan, M.; Hamroun, D.; Heraoui, D.; Rossi-Semerano, L.; Berger, M.G.; Rose, C.; Camou, F.; de Roux-Serratrice, C.; Grosbois, B.; et al. The French Gaucher’s disease registry: Clinical characteristics, complications and treatment of 562 patients. Orphanet J. Rare Dis. 2012, 7, 77. [Google Scholar] [CrossRef]
- Liu, G.; Boot, B.; Locascio, J.J.; Jansen, I.E.; Winder-Rhodes, S.; Eberly, S.; Elbaz, A.; Brice, A.; Ravina, B.; van Hilten, J.J.; et al. Specifically neuropathic Gaucher’s mutations accelerate cognitive decline in Parkinson’s. Ann. Neurol. 2016, 80, 674–685. [Google Scholar] [CrossRef]
- Zhang, Y.; Shu, L.; Sun, Q.; Zhou, X.; Pan, H.; Guo, J.; Tang, B. Integrated Genetic Analysis of Racial Differences of Common GBA Variants in Parkinson’s Disease: A Meta-Analysis. Front. Mol. Neurosci. 2018, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Deng, L.; Zhong, Y.; Yi, M. The Association between E326K of GBA and the Risk of Parkinson’s Disease. Parkinsons Dis. 2018, 2018, 1048084. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, Q.Y.; Zhao, Y.W.; Shu, L.; Guo, J.F.; Xu, Q.; Yan, X.X.; Tang, B.S. Effect of GBA Mutations on Phenotype of Parkinson’s Disease: A Study on Chinese Population and a Meta-Analysis. Parkinsons Dis. 2015, 2015, 916971. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, H.A.; González, A.E.; Cerda-Troncoso, C.; Shaughnessy, R.; Otth, C.; Soza, A.; Burgos, P.V. Interplay Between the Autophagy-Lysosomal Pathway and the Ubiquitin-Proteasome System: A Target for Therapeutic Development in Alzheimer’s Disease. Front. Cell. Neurosci. 2018, 12, 126. [Google Scholar] [CrossRef] [PubMed]
- Pitcairn, C.; Wani, W.Y.; Mazzolli, J. Dysregulation of the autophagic-lysosomal pathway in Gaucher Disease and Parkinson’s Disease. Neurobiol. Dis. 2018, 122, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.; Jansen Steur, E.N.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Gegg, M.E.; Schapira, A.H.V. The role of glucocerebrosidase in Parkinson disease pathogenesis. FEBS J. 2018, 285, 3591–3603. [Google Scholar] [CrossRef] [PubMed]
- Zunke, F.; Andresen, L.; Wesseler, S.; Groth, J.; Arnold, P.; Rothaug, M.; Mazzulli, J.R.; Krainc, D.; Blanz, J.; Saftig, P.; et al. Characterization of the complex formed by β-glucocerebrosidase and the lysosomal integral membrane protein type-2. Proc. Natl. Acad. Sci. USA 2016, 113, 3791–3796. [Google Scholar] [CrossRef] [PubMed]
- Mazzulli, J.R.; Xu, Y.H.; Sun, Y.; Knight, A.L.; McLean, P.J.; Caldwell, G.A.; Sidransky, E.; Grabowski, G.A.; Krainc, D. Gaucher disease glucocerebrosidase and α-synuclein form a bidirectional pathogenic loop in synucleinopathies. Cell 2011, 146, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, J.; Gegg, M.E.; Migdalska-Richards, A.; Doherty, M.K.; Whitfield, P.D.; Schapira, A.H. Autophagic lysosome reformation dysfunction in glucocerebrosidase deficient cells: Relevance to Parkinson disease. Hum. Mol. Genet. 2016, 25, 3432–3445. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.E.; Vincow, E.S.; Merrihew, G.E.; MacCoss, M.J.; Davis, M.Y.; Pallanck, L.J. Glucocerebrosidase deficiency promotes protein aggregation through dysregulation of extracellular vesicles. PLoS Genet. 2018, 14, e1007694. [Google Scholar] [CrossRef] [PubMed]
- Berthet, A.; Margolis, E.B.; Zhang, J.; Hsieh, I.; Zhang, J.; Hnasko, T.S.; Ahmad, J.; Edwards, R.H.; Sesaki, H.; Huang, E.J.; et al. Loss of mitochondrial fission depletes axonal mitochondria in midbrain dopamine neurons. J. Neurosci. 2014, 34, 14304–14317. [Google Scholar] [CrossRef] [PubMed]
- Poole, A.C.; Thomas, R.E.; Andrews, L.A.; McBride, H.M.; Whitworth, A.J.; Pallanck, L.J. The PINK1/Parkin pathway regulates mitochondrial morphology. Proc. Natl. Acad. Sci. USA 2008, 105, 1638–1643. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Petrie, T.G.; Liu, Y.; Liu, J.; Fujioka, H.; Zhu, X. Parkinson’s disease-associated DJ-1 mutations impair mitochondrial dynamics and cause mitochondrial dysfunction. J. Neurochem. 2012, 121, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yan, M.H.; Fujioka, H.; Liu, J.; Wilson-Delfosse, A.; Chen, S.G.; Perry, G.; Casadesus, G.; Zhu, X. LRRK2 regulates mitochondrial dynamics and function through direct interaction with DLP1. Hum. Mol. Genet. 2012, 21, 1931–1944. [Google Scholar] [CrossRef] [PubMed]
- Kamp, F.; Exner, N.; Lutz, A.K.; Wender, N.; Hegermann, J.; Brunner, B.; Nuscher, B.; Bartels, T.; Giese, A.; Beyer, K.; et al. Inhibition of mitochondrial fusion by α-synuclein is rescued by PINK1, Parkin and DJ-1. EMBO J. 2010, 29, 3571–3589. [Google Scholar] [CrossRef]
- Yun, S.P.; Kim, D.; Kim, S.; Kim, S.; Karuppagounder, S.S.; Kwon, S.H.; Lee, S.; Kam, T.I.; Lee, S.; Ham, S.; et al. α-Synuclein accumulation and GBA deficiency due to L444P GBA mutation contributes to MPTP-induced parkinsonism. Mol. Neurodegener. 2018, 13. [Google Scholar] [CrossRef]
- Bendikov-Bar, I.; Rapaport, D.; Larisch, S.; Horowitz, M. Parkin-mediated ubiquitination of mutant glucocerebrosidase leads to competition with its substrates PARIS and ARTS. Orphanet J. Rare Dis. 2014, 9, 86. [Google Scholar] [CrossRef]
- Li, H.; Ham, A.; Ma, T.C.; Kuo, S.H.; Kanter, E.; Kim, D.; Ko, H.S.; Quan, Y.; Sardi, S.P.; Li, A.; et al. Mitochondrial dysfunction and mitophagy defect triggered by heterozygous GBA mutations. Autophagy 2019, 15, 113–130. [Google Scholar] [CrossRef]
- Stirnemann, J.; Belmatoug, N.; Camou, F.; Serratrice, C.; Froissart, R.; Caillaud, C.; Levade, T.; Astudillo, L.; Serratrice, J.; Brassier, A.; et al. A Review of Gaucher Disease Pathophysiology, Clinical Presentation and Treatments. Int. J. Mol. Sci. 2017, 18, 441. [Google Scholar] [CrossRef]
- Extracted from Clinical Trials. Available online: clinicaltrials.gov/ct2/show/NCT02906020 (accessed on 11 December 2018).
- Sardi, S.P.; Viel, C.; Clarke, J.; Treleaven, C.M.; Richards, A.M.; Park, H.; Olszewski, M.A.; Dodge, J.C.; Marshall, J.; Makino, E.; et al. GCS inhibitor treats synucleinopathy models. Proc. Natl. Acad. Sci. USA 2017, 114, 2699–2704. [Google Scholar] [CrossRef] [PubMed]
- McNeill, A.; Magalhaes, J.; Shen, C.; Chau, K.Y.; Hughes, D.; Mehta, A.; Foltynie, T.; Cooper, J.M.; Abramov, A.Y.; Gegg, M.; et al. Ambroxol improves lysosomal biochemistry in glucocerebrosidase mutation-linked Parkinson disease cells. Brain 2014, 137, 1481–1495. [Google Scholar] [CrossRef] [PubMed]
- Aflaki, E.; Borger, D.K.; Moaven, N.; Stubblefield, B.K.; Rogers, S.A.; Patnaik, S.; Schoenen, F.J.; Westbroek, W.; Zheng, W.; Sullivan, P.; et al. A New Glucocerebrosidase Chaperone Reduces α-Synuclein and Glycolipid Levels in iPSC-Derived Dopaminergic Neurons from Patients with Gaucher Disease and Parkinsonism. J. Neurosci. 2016, 36, 7441–7452. [Google Scholar] [CrossRef] [PubMed]
- Mistry, P.K.; Lopez, G.; Schiffmann, R.; Barton, N.W.; Weinreb, N.J.; Sidransky, E. 27 Progress and ongoing challenges. Mol. Genet. Metab. 2016, 120, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Zunke, F.; Moise, A.C.; Belur, N.R.; Gelyana, E.; Stojkovska, I.; Dzaferbegovic, H.; Toker, N.J.; Jeon, S.; Fredriksen, K.; Mazzulli, J.R. Reversible Conformational Conversion of α-Synuclein into Toxic Assemblies by Glucosylceramide. Neuron 2018, 97, 92–107. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Jeon, S.; Burbulla, L.F.; Krainc, D. Acid ceramidase inhibition ameliorates α-synuclein accumulation upon loss of GBA1 function. Hum. Mol. Genet. 2018, 27, 1972–1988. [Google Scholar] [CrossRef] [PubMed]
- Espay, A.J.; Brundin, P.; Lang, A.E. Precision medicine for disease modification in Parkinson disease. Nat. Rev. Neurol. 2017, 13, 119–126. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Biomarker | Observation | References |
|---|---|---|
| Clinical biomarker | Early multidomain cognitive impairment. More severe Levodopa induced dyskinesias. | |
| Transcranial sonography | Nigral hyperechogenicity. | [49] |
| PET 1 8F dopa | Decreased striatal dopamine synthesis, similar to iPD. Bilateral asymmetric reduction in striatal uptake. | [50,51] |
| fMRI | Significant hypometabolism in glucose metabolism in supplementary motor area and parieto-occipital cortices. Hypermetabolism of the lentiform nuclei and thalamus. Decrease in the parieto-occipital and to a certain degree anteromedial frontal cortex. | [52] |
| Diffusion tensor MRI | Decreased frontal cortico-cortical and parahippocampal tracts in GBA-PD. Decreased fractional anisotropy of the corpus callosum, olfactory tract, anterior limb of the internal capsule, cingulum, external capsule bilaterally, and left superior longitudinal fasciculus. | [52] |
| Postsynaptic DA 11 C-Raclopride | Postsynaptic dopamine terminal persistence of higher putaminal uptake in advanced disease. | [53] |
| Null or Severe GBAmtt | Mild GD | |
|---|---|---|
| L444P | N307S | |
| Phenotype | Onset infancy and childhood, rapid progression shorter life expectancy, and appearance of more severe neurologic features (GD2, GD3) | 50% GD-PD homozygous for N307S 90% GD-PD carry at least one N307S mutation |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gatto, E.M.; Da Prat, G.; Etcheverry, J.L.; Drelichman, G.; Cesarini, M. Parkinsonisms and Glucocerebrosidase Deficiency: A Comprehensive Review for Molecular and Cellular Mechanism of Glucocerebrosidase Deficiency. Brain Sci. 2019, 9, 30. https://doi.org/10.3390/brainsci9020030
Gatto EM, Da Prat G, Etcheverry JL, Drelichman G, Cesarini M. Parkinsonisms and Glucocerebrosidase Deficiency: A Comprehensive Review for Molecular and Cellular Mechanism of Glucocerebrosidase Deficiency. Brain Sciences. 2019; 9(2):30. https://doi.org/10.3390/brainsci9020030
Chicago/Turabian StyleGatto, Emilia M., Gustavo Da Prat, Jose Luis Etcheverry, Guillermo Drelichman, and Martin Cesarini. 2019. "Parkinsonisms and Glucocerebrosidase Deficiency: A Comprehensive Review for Molecular and Cellular Mechanism of Glucocerebrosidase Deficiency" Brain Sciences 9, no. 2: 30. https://doi.org/10.3390/brainsci9020030
APA StyleGatto, E. M., Da Prat, G., Etcheverry, J. L., Drelichman, G., & Cesarini, M. (2019). Parkinsonisms and Glucocerebrosidase Deficiency: A Comprehensive Review for Molecular and Cellular Mechanism of Glucocerebrosidase Deficiency. Brain Sciences, 9(2), 30. https://doi.org/10.3390/brainsci9020030