
Influence of Inherited Seizure Susceptibility on Intermittent Voluntary Alcohol Consumption and Alcohol Withdrawal Seizures in Genetically Epilepsy-Prone Rats (GEPR-3s)



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
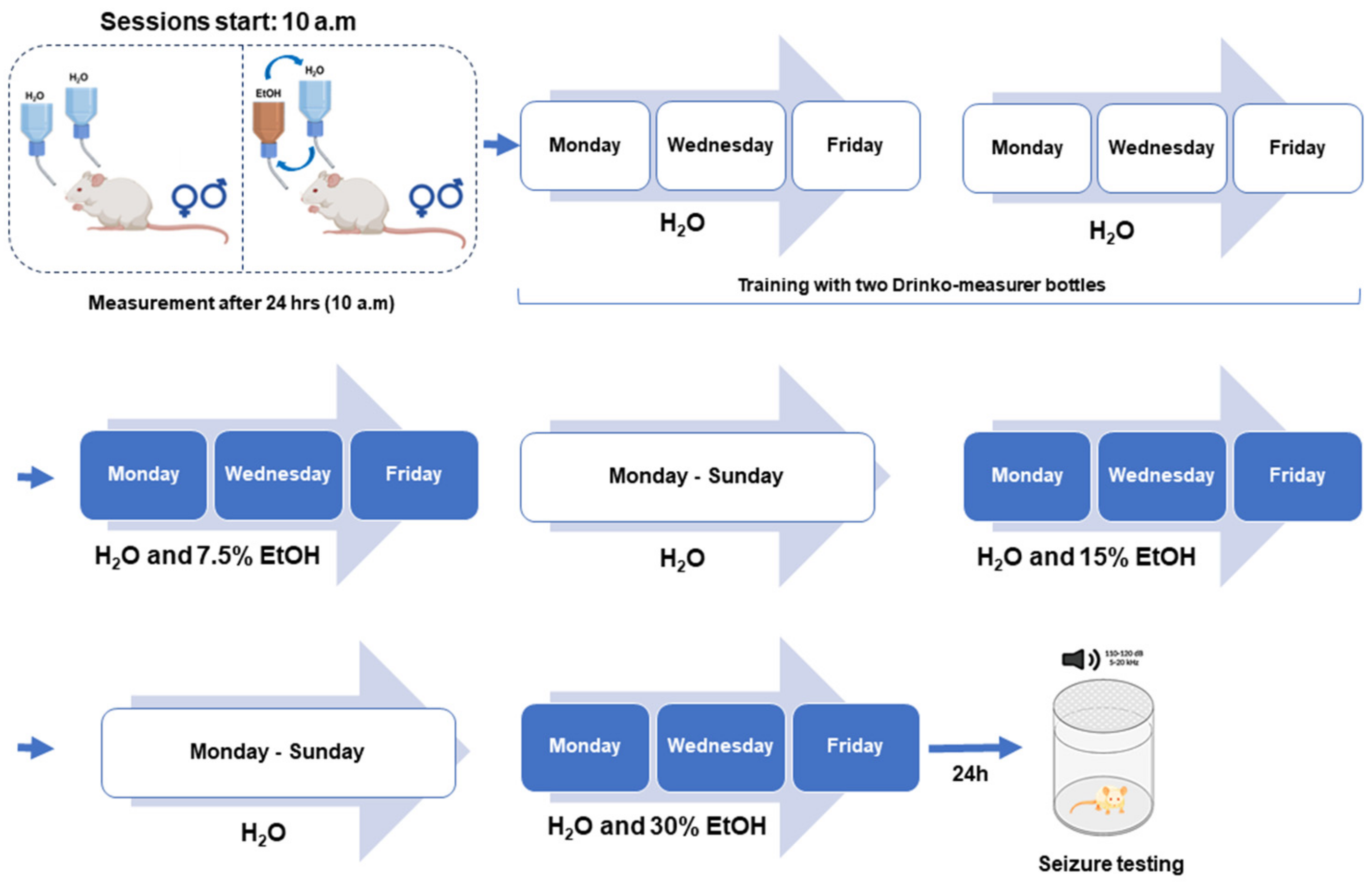
2.2. Voluntary Ethanol Consumption Paradigm: Intermittent Alcohol Access Two-Bottle Choice Paradigm
2.3. Acoustically Evoked Seizure Testing
2.4. Data Analysis
3. Results
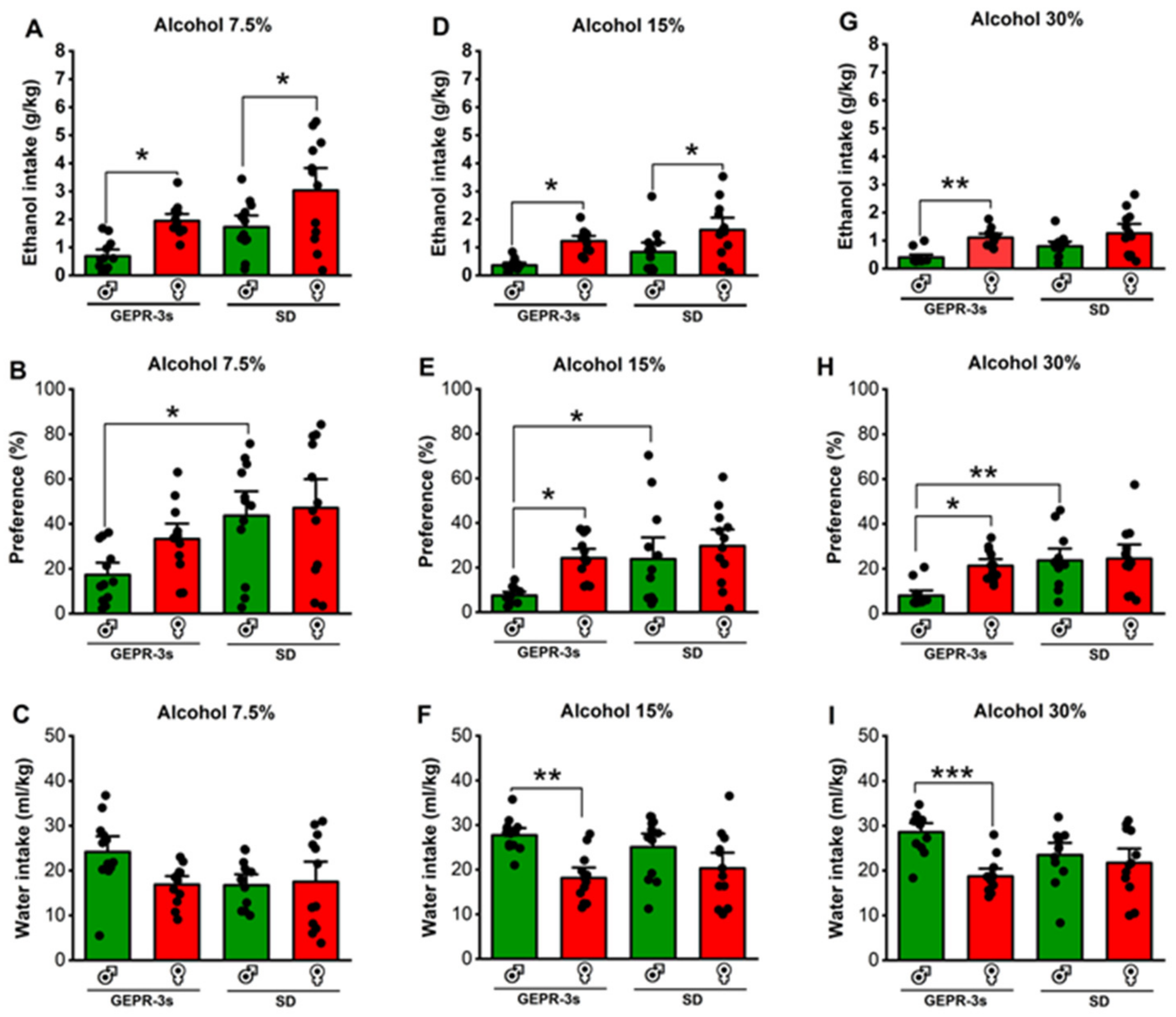
3.1. Effects of Strain, Sex, and Ethanol Concentrations on Voluntary Ethanol Consumption
3.2. Effects of Voluntary Ethanol Consumption: 7.5% Alcohol Concentration
3.3. Effects of Voluntary Ethanol Consumption: 15% Alcohol Concentration
3.4. Effects of Voluntary Ethanol Consumption: 30% Alcohol Concentration
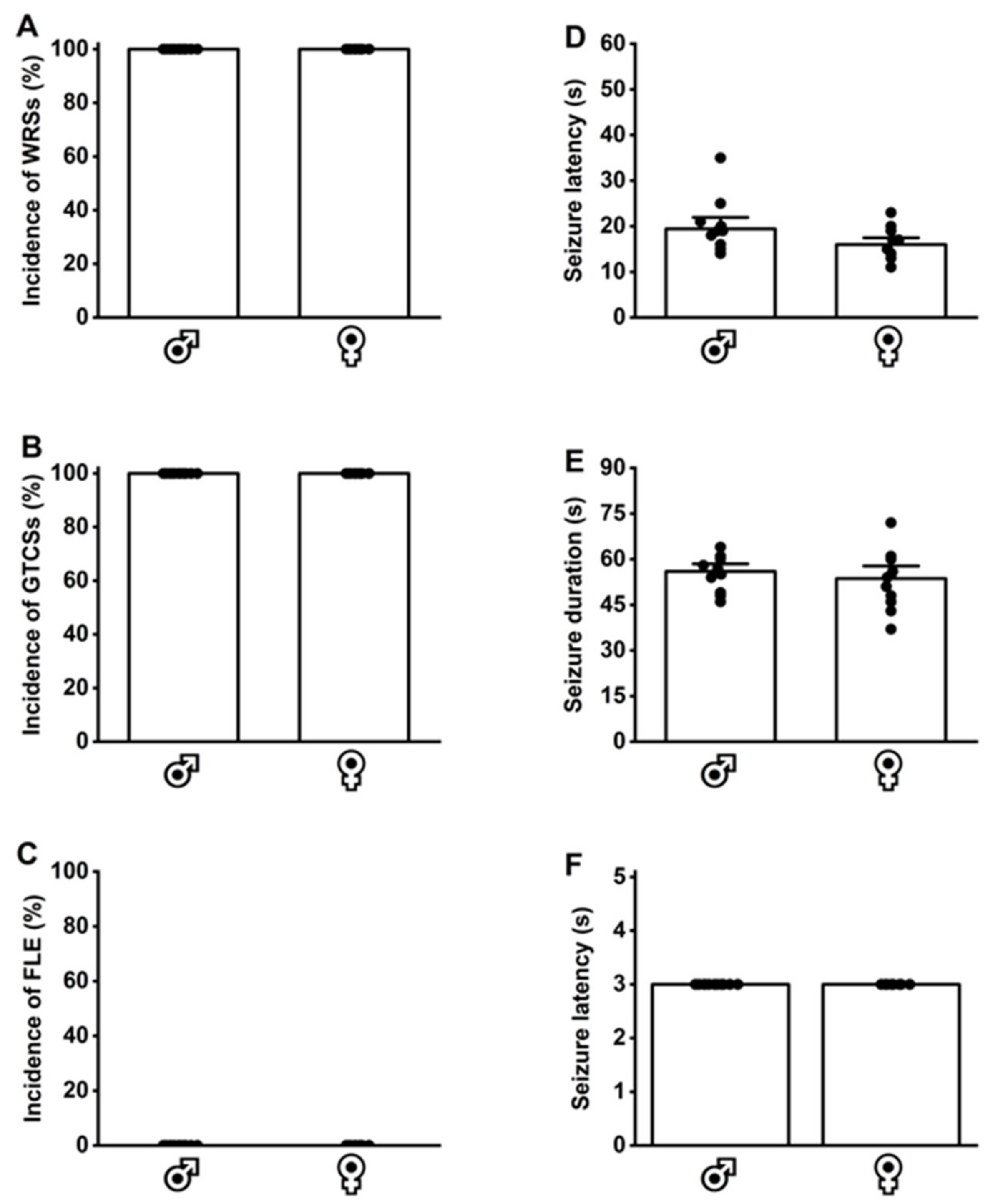
3.5. Effects of Withdrawal from Voluntary Ethanol Consumption on Acoustically Evoked Seizures in GEPR-3s and SD Rats
3.6. Acoustically Evoked Seizure Susceptibility at Postnatal Day 28 in Seizures in GEPR-3s and SD Rats
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cohen, S.M.; Alexander, R.S.; Holt, S.R. The Spectrum of Alcohol Use: Epidemiology, Diagnosis, and Treatment. Med. Clin. N. Am. 2022, 106, 43–60. [Google Scholar] [CrossRef] [PubMed]
- Freedland, E.S.; McMicken, D.B. Alcohol-Related Seizures, Part I: Pathophysiology, Differential Diagnosis, and Evaluation. J. Emerg. Med. 1993, 11, 463–473. [Google Scholar] [CrossRef]
- Hillbom, M.; Pieninkeroinen, I.; Leone, M. Seizures in Alcohol-Dependent Patients: Epidemiology, Pathophysiology and Management. CNS Drugs 2003, 17, 1013–1030. [Google Scholar] [CrossRef] [PubMed]
- Hamerle, M.; Ghaeni, L.; Kowski, A.; Weissinger, F.; Holtkamp, M. Alcohol Use and Alcohol-Related Seizures in Patients With Epilepsy. Front. Neurol. 2018, 9, 401. [Google Scholar] [CrossRef]
- Hauser, W.A.; Ng, S.K.; Brust, J.C. Alcohol, Seizures, and Epilepsy. Epilepsia 1988, 29 (Suppl. S2), S66–S78. [Google Scholar] [CrossRef]
- Scorza, F.A.; Cavalheiro, E.A.; Scorza, C.A.; de Almeida, A.-C.G. “I’m Afraid I Have Bad News for You …” Alcohol Contributes to the Occurrence of Sudden Unexpected Death in Epilepsy and Years Lost. Epilepsy Behav. 2014, 36, 131–132. [Google Scholar] [CrossRef] [PubMed]
- Scorza, F.A.; Terra, V.C.; Arida, R.M.; Scorza, C.A.; Cavalheiro, E.A. Sudden Unexpected Death in Epilepsy and Winter Temperatures: It’s Important to Know That It’s c-c-c-c-Cold Outside. Epilepsy Behav. 2009, 14, 707, author reply 708. [Google Scholar] [CrossRef]
- Höppener, R.J.; Kuyer, A.; van der Lugt, P.J. Epilepsy and Alcohol: The Influence of Social Alcohol Intake on Seizures and Treatment in Epilepsy. Epilepsia 1983, 24, 459–471. [Google Scholar] [CrossRef]
- Woo, K.N.; Kim, K.; Ko, D.S.; Kim, H.-W.; Kim, Y.H. Alcohol Consumption on Unprovoked Seizure and Epilepsy: An Updated Meta-Analysis. Drug Alcohol. Depend. 2022, 232, 109305. [Google Scholar] [CrossRef]
- Scorza, F.A.; Arida, R.M.; Cysneiros, R.M.; Priel, M.R.; de Albuquerque, M.; Cavalheiro, E.A. The Effects of Alcohol Intake and Withdrawal on the Seizures Frequency and Hippocampal Morphology in Rats with Epilepsy. Neurosci. Res. 2003, 47, 323–328. [Google Scholar] [CrossRef]
- Kosobud, A.; Bodor, A.S.; Crabbe, J.C. Voluntary Consumption of Ethanol in WSP, WSC and WSR Selectively Bred Mouse Lines. Pharmacol. Biochem. Behav. 1988, 29, 601–607. [Google Scholar] [CrossRef]
- Crabbe, J.C.; Spence, S.E.; Huang, L.C.; Cameron, A.J.; Schlumbohm, J.P.; Barkley-Levenson, A.M.; Metten, P. Ethanol Drinking in Withdrawal Seizure-Prone and -Resistant Selected Mouse Lines. Alcohol 2013, 47, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Iannaccone, P.M.; Jacob, H.J. Rats! Dis. Models Mech. 2009, 2, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Nentwig, T.B.; Myers, K.P.; Grisel, J.E. Initial Subjective Reward to Alcohol in Sprague-Dawley Rats. Alcohol 2017, 58, 19–22. [Google Scholar] [CrossRef]
- Faingold, C.L. Neuronal Networks in the Genetically Epilepsy-Prone Rat. Adv. Neurol. 1999, 79, 311–321. [Google Scholar] [PubMed]
- Jobe, P.C.; Dailey, J.W. Genetically Epilepsy-Prone Rats (GEPRs) in Drug Research. CNS Drug Rev. 2000, 6, 241–260. [Google Scholar] [CrossRef]
- Jobe, P.C.; Browning, R.A. MODELS|Seizure Predisposition and Expression in the Genetic Epilepsies as Exemplified by the Genetically Epilepsy-Prone Rats. In Encyclopedia of Basic Epilepsy Research; Schwartzkroin, P.A., Ed.; Academic Press: Oxford, UK, 2009; pp. 792–798. ISBN 978-0-12-373961-2. [Google Scholar]
- Mishra, P.K.; Dailey, J.W.; Reigel, C.E.; Jobe, P.C. Audiogenic Convulsions in Moderate Seizure Genetically Epilepsy-Prone Rats (GEPR-3s). Epilepsy Res. 1989, 3, 191–198. [Google Scholar] [CrossRef]
- Faingold, C.L. The Majchrowicz Binge Alcohol Protocol: An Intubation Technique to Study Alcohol Dependence in Rats. CP Neurosci. 2008, 44, 9.28.1–9.28.12. [Google Scholar] [CrossRef]
- Linseman, M.A. Alcohol Consumption in Free-Feeding Rats: Procedural, Genetic and Pharmacokinetic Factors. Psychopharmacology 1987, 92, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Melchior, C.L.; Myers, R.D. Genetic Differences in Ethanol Drinking of the Rat Following Injection of 6-OHDA, 5,6-DHT or 5,7-DHT into the Cerebral Ventricles. Pharmacol. Biochem. Behav. 1976, 5, 63–72. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academies Collection: Reports Funded by National Institutes of Health; National Academies Press (US): Washington, DC, USA, 2011; ISBN 978-0-309-15400-0. [Google Scholar]
- Simms, J.A.; Steensland, P.; Medina, B.; Abernathy, K.E.; Chandler, L.J.; Wise, R.; Bartlett, S.E. Intermittent Access to 20% Ethanol Induces High Ethanol Consumption in Long-Evans and Wistar Rats. Alcohol. Clin. Exp. Res. 2008, 32, 1816–1823. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.A. Voluntary Ethanol Intake in Rats Following Exposure to Ethanol on Various Schedules. Psychopharmacologia 1973, 29, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Samokhvalov, A.V.; Irving, H.; Mohapatra, S.; Rehm, J. Alcohol Consumption, Unprovoked Seizures, and Epilepsy: A Systematic Review and Meta-Analysis. Epilepsia 2010, 51, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Kommajosyula, S.P.; Randall, M.E.; Tupal, S.; Faingold, C.L. Alcohol withdrawal in epileptic rats-effects on postictal depression, respiratory, and death. Epilepsy Behav. 2016, 64, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Turski, L.; Ikonomidou, C.; Turski, W.A.; Bortolotto, Z.A.; Cavalheiro, E.A. Review: Cholinergic Mechanisms and Epileptogenesis. The Seizures Induced by Pilocarpine: A Novel Experimental Model of Intractable Epilepsy. Synapse 1989, 3, 154–171. [Google Scholar] [CrossRef]
- Naritoku, D.K.; Mecozzi, L.B.; Aiello, M.T.; Faingold, C.L. Repetition of Audiogenic Seizures in Genetically Epilepsy-Prone Rats Induces Cortical Epileptiform Activity and Additional Seizure Behaviors. Exp. Neurol. 1992, 115, 317–324. [Google Scholar] [CrossRef]
- Siciliano, C.A.; Noamany, H.; Chang, C.-J.; Brown, A.R.; Chen, X.; Leible, D.; Lee, J.J.; Wang, J.; Vernon, A.N.; Vander Weele, C.M.; et al. A Cortical-Brainstem Circuit Predicts and Governs Compulsive Alcohol Drinking. Science 2019, 366, 1008–1012. [Google Scholar] [CrossRef]
- Broadbent, J.; Muccino, K.J.; Cunningham, C.L. Ethanol-Induced Conditioned Taste Aversion in 15 Inbred Mouse Strains. Behav. Neurosci. 2002, 116, 138–148. [Google Scholar] [CrossRef]
- Cunningham, C.L. Genetic Relationships Between Ethanol-Induced Conditioned Place Aversion and Other Ethanol Phenotypes in 15 Inbred Mouse Strains. Brain Sci. 2019, 9, 209. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L. Genetic Relationship between Ethanol-Induced Conditioned Place Preference and Other Ethanol Phenotypes in 15 Inbred Mouse Strains. Behav. Neurosci. 2014, 128, 430–445. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L. Localization of Genes Influencing Ethanol-Induced Conditioned Place Preference and Locomotor Activity in BXD Recombinant Inbred Mice. Psychopharmacology 1995, 120, 28–41. [Google Scholar] [CrossRef]
- Risinger, F.O.; Cunningham, C.L. Ethanol-Induced Conditioned Taste Aversion in BXD Recombinant Inbred Mice. Alcohol. Clin. Exp. Res. 1998, 22, 1234–1244. [Google Scholar] [CrossRef] [PubMed]
- Roma, P.G.; Flint, W.W.; Higley, J.D.; Riley, A.L. Assessment of the Aversive and Rewarding Effects of Alcohol in Fischer and Lewis Rats. Psychopharmacology 2006, 189, 187–199. [Google Scholar] [CrossRef]
- Schramm-Sapyta, N.L.; DiFeliceantonio, A.G.; Foscue, E.; Glowacz, S.; Haseeb, N.; Wang, N.; Zhou, C.; Kuhn, C.M. Aversive Effects of Ethanol in Adolescent versus Adult Rats: Potential Causes and Implication for Future Drinking. Alcohol. Clin. Exp. Res. 2010, 34, 2061–2069. [Google Scholar] [CrossRef] [PubMed]
- Nienhuis, R.; Olds, J. Changes in Unit Responses to Tones after Food Reinforcement in the Auditory Pathway of the Rat: Intertrial Arousal. Exp. Neurol. 1978, 59, 229–242. [Google Scholar] [CrossRef]
- Ruth, R.E.; Rosenfeld, J.P.; Harris, D.M.; Birkel, P. Effects of Aversive and Rewarding Electrical Brain Stimulation on Auditory Evoked Responses in Albino Rat Tectum. Physiol. Behav. 1974, 13, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Vander Weele, C.M.; Siciliano, C.A.; Matthews, G.A.; Namburi, P.; Izadmehr, E.M.; Espinel, I.C.; Nieh, E.H.; Schut, E.H.S.; Padilla-Coreano, N.; Burgos-Robles, A.; et al. Dopamine Enhances Signal-to-Noise Ratio in Cortical-Brainstem Encoding of Aversive Stimuli. Nature 2018, 563, 397–401. [Google Scholar] [CrossRef]
- Zhou, Z.; Metten, P.; Yuan, Q.; Sun, H.; Hodgkinson, C.A.; Shen, P.-H.; Marietta, C.; Crabbe, J.C.; Goldman, D. Genetic and Genomic Signatures in Ethanol Withdrawal Seizure-Prone and Seizure-Resistant Mice Implicate Genes Involved in Epilepsy and Neuronal Excitability. Mol. Psychiatry 2022, 27, 4611–4623. [Google Scholar] [CrossRef] [PubMed]
- Mulholland, P.J. K(Ca)2 Channels: Novel Therapeutic Targets for Treating Alcohol Withdrawal and Escalation of Alcohol Consumption. Alcohol 2012, 46, 309–315. [Google Scholar] [CrossRef]
- Rinker, J.A.; Mulholland, P.J. Promising Pharmacogenetic Targets for Treating Alcohol Use Disorder: Evidence from Preclinical Models. Pharmacogenomics 2017, 18, 555–570. [Google Scholar] [CrossRef]
- Hopf, F.W.; Simms, J.A.; Chang, S.-J.; Seif, T.; Bartlett, S.E.; Bonci, A. Chlorzoxazone, an SK Channel Activator Used in Humans, Reduces Excessive Alcohol Intake in Rats. Biol. Psychiatry 2011, 69, 618–624. [Google Scholar] [CrossRef]
- Hopf, F.W.; Bowers, M.S.; Chang, S.-J.; Chen, B.T.; Martin, M.; Seif, T.; Cho, S.L.; Tye, K.; Bonci, A. Reduced Nucleus Accumbens SK Channel Activity Enhances Alcohol Seeking during Abstinence. Neuron 2010, 65, 682–694. [Google Scholar] [CrossRef]
- Knapp, C.M.; O’Malley, M.; Datta, S.; Ciraulo, D.A. The Kv7 Potassium Channel Activator Retigabine Decreases Alcohol Consumption in Rats. Am. J. Drug Alcohol Abus. 2014, 40, 244–250. [Google Scholar] [CrossRef] [PubMed]
- McGuier, N.S.; Griffin, W.C.; Gass, J.T.; Padula, A.E.; Chesler, E.J.; Mulholland, P.J. Kv7 Channels in the Nucleus Accumbens Are Altered by Chronic Drinking and Are Targets for Reducing Alcohol Consumption. Addict. Biol. 2016, 21, 1097–1112. [Google Scholar] [CrossRef] [PubMed]
- Herman, M.A.; Sidhu, H.; Stouffer, D.G.; Kreifeldt, M.; Le, D.; Cates-Gatto, C.; Munoz, M.B.; Roberts, A.J.; Parsons, L.H.; Roberto, M.; et al. GIRK3 Gates Activation of the Mesolimbic Dopaminergic Pathway by Ethanol. Proc. Natl. Acad. Sci. USA 2015, 112, 7091–7096. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Luypaert, C.; Sáez-Cortez, F.; Quintanilla, M.E.; Herrera-Marschitz, M.; Rivera-Meza, M. Gene Knockdown of HCN2 Ion Channels in the Ventral Tegmental Area Reduces Ethanol Consumption in Alcohol Preferring Rats. Am. J. Drug Alcohol. Abus. 2022, 48, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Rassnick, S.; Pulvirenti, L.; Koob, G.F. Oral Ethanol Self-Administration in Rats Is Reduced by the Administration of Dopamine and Glutamate Receptor Antagonists into the Nucleus Accumbens. Psychopharmacology 1992, 109, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Szot, P.; Reigel, C.E.; White, S.S.; Veith, R.C. Alterations in mRNA Expression of Systems That Regulate Neurotransmitter Synaptic Content in Seizure-Naive Genetically Epilepsy-Prone Rat (GEPR): Transporter Proteins and Rate-Limiting Synthesizing Enzymes for Norepinephrine, Dopamine and Serotonin. Brain Res. Mol. Brain Res. 1996, 43, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Tsyba, E.T.; Midzyanovskaya, I.S.; Birioukova, L.M.; Tuomisto, L.M.; van Luijtelaar, G.; Abbasova, K.R. Striatal Patchwork of D1-like and D2-like Receptors Binding Densities in Rats with Genetic Audiogenic and Absence Epilepsies. Diagnostics 2023, 13, 587. [Google Scholar] [CrossRef]
- Cornelius, J.R.; Bukstein, O.; Salloum, I.; Clark, D. Alcohol and Psychiatric Comorbidity. In Recent Developments in Alcoholism: Research on Alcoholism Treatment; Galanter, M., Begleiter, H., Deitrich, R., Gallant, D., Gottheil, E., Paredes, A., Rothschild, M., Van Theil, D., Lagressa, D., Allen, J.P., et al., Eds.; Recent Developments in Alcoholism; Springer US: Boston, MA, USA, 2002; pp. 361–374. ISBN 978-0-306-47939-7. [Google Scholar]
- Aguilar, B.L.; Malkova, L.; N’Gouemo, P.; Forcelli, P.A. Genetically Epilepsy-Prone Rats Display Anxiety-Like Behaviors and Neuropsychiatric Comorbidities of Epilepsy. Front. Neurol. 2018, 9, 476. [Google Scholar] [CrossRef]
- Flores-Bonilla, A.; De Oliveira, B.; Silva-Gotay, A.; Lucier, K.W.; Richardson, H.N. Shortening Time for Access to Alcohol Drives up Front-Loading Behavior, Bringing Consumption in Male Rats to the Level of Females. Biol. Sex. Differ. 2021, 12, 51. [Google Scholar] [CrossRef] [PubMed]
- Foo, J.C.; Skorodumov, I.; Spanagel, R.; Meinhardt, M.W. Sex- and Age-Specific Effects on the Development of Addiction and Compulsive-like Drinking in Rats. Biol. Sex. Differ. 2023, 14, 44. [Google Scholar] [CrossRef] [PubMed]
- Radke, A.K.; Sneddon, E.A.; Frasier, R.M.; Hopf, F.W. Recent Perspectives on Sex Differences in Compulsion-Like and Binge Alcohol Drinking. Int. J. Mol. Sci. 2021, 22, 3788. [Google Scholar] [CrossRef] [PubMed]
- Carroll, M.R.; Rodd, Z.A.; Murphy, J.M.; Simon, J.R. Chronic Ethanol Consumption Increases Dopamine Uptake in the Nucleus Accumbens of High Alcohol Drinking Rats. Alcohol 2006, 40, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Quigley, J.A.; Logsdon, M.K.; Turner, C.A.; Gonzalez, I.L.; Leonardo, N.B.; Becker, J.B. Sex Differences in Vulnerability to Addiction. Neuropharmacology 2021, 187, 108491. [Google Scholar] [CrossRef] [PubMed]
- Satta, R.; Hilderbrand, E.R.; Lasek, A.W. Ovarian Hormones Contribute to High Levels of Binge-Like Drinking by Female Mice. Alcohol. Clin. Exp. Res. 2018, 42, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Zachry, J.E.; Nolan, S.O.; Brady, L.J.; Kelly, S.J.; Siciliano, C.A.; Calipari, E.S. Sex Differences in Dopamine Release Regulation in the Striatum. Neuropsychopharmacology 2021, 46, 491–499. [Google Scholar] [CrossRef]
- Hilderbrand, E.R.; Lasek, A.W. Estradiol Enhances Ethanol Reward in Female Mice through Activation of ERα and ERβ. Horm. Behav. 2018, 98, 159–164. [Google Scholar] [CrossRef]
- Thomasson, H.R.; Beard, J.D.; Li, T.-K. ADH2 Gene Polymorphisms Are Determinants of Alcohol Pharmacokinetics. Alcohol. Clin. Exp. Res. 1995, 19, 1494–1499. [Google Scholar] [CrossRef]
- Chappell, A.M.; Carter, E.; McCool, B.A.; Weiner, J.L. Adolescent Rearing Conditions Influence the Relationship between Initial Anxiety-like Behavior and Ethanol Drinking in Male Long Evans Rats. Alcohol. Clin. Exp. Res. 2013, 37 (Suppl. S1), E394–E403. [Google Scholar] [CrossRef]
- Moench, K.M.; Logrip, M.L. Housing Condition Differentially Impacts Escalation of Alcohol Intake, Relapse-Like Drinking, Anxiety-Like Behavior, and Stress History Effects by Sex. Alcohol. Clin. Exp. Res. 2021, 45, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Scott, H.; Tjernström, N.; Roman, E. Effects of Pair Housing on Voluntary Alcohol Intake in Male and Female Wistar Rats. Alcohol. 2020, 86, 121–128. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva-Cardoso, G.K.; N’Gouemo, P. Influence of Inherited Seizure Susceptibility on Intermittent Voluntary Alcohol Consumption and Alcohol Withdrawal Seizures in Genetically Epilepsy-Prone Rats (GEPR-3s). Brain Sci. 2024, 14, 188. https://doi.org/10.3390/brainsci14020188
Silva-Cardoso GK, N’Gouemo P. Influence of Inherited Seizure Susceptibility on Intermittent Voluntary Alcohol Consumption and Alcohol Withdrawal Seizures in Genetically Epilepsy-Prone Rats (GEPR-3s). Brain Sciences. 2024; 14(2):188. https://doi.org/10.3390/brainsci14020188
Chicago/Turabian StyleSilva-Cardoso, Gleice Kelli, and Prosper N’Gouemo. 2024. "Influence of Inherited Seizure Susceptibility on Intermittent Voluntary Alcohol Consumption and Alcohol Withdrawal Seizures in Genetically Epilepsy-Prone Rats (GEPR-3s)" Brain Sciences 14, no. 2: 188. https://doi.org/10.3390/brainsci14020188
APA StyleSilva-Cardoso, G. K., & N’Gouemo, P. (2024). Influence of Inherited Seizure Susceptibility on Intermittent Voluntary Alcohol Consumption and Alcohol Withdrawal Seizures in Genetically Epilepsy-Prone Rats (GEPR-3s). Brain Sciences, 14(2), 188. https://doi.org/10.3390/brainsci14020188