

Cellular and Molecular Mechanisms Underlying Synaptic Subcellular Specificity


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Secreted Molecules
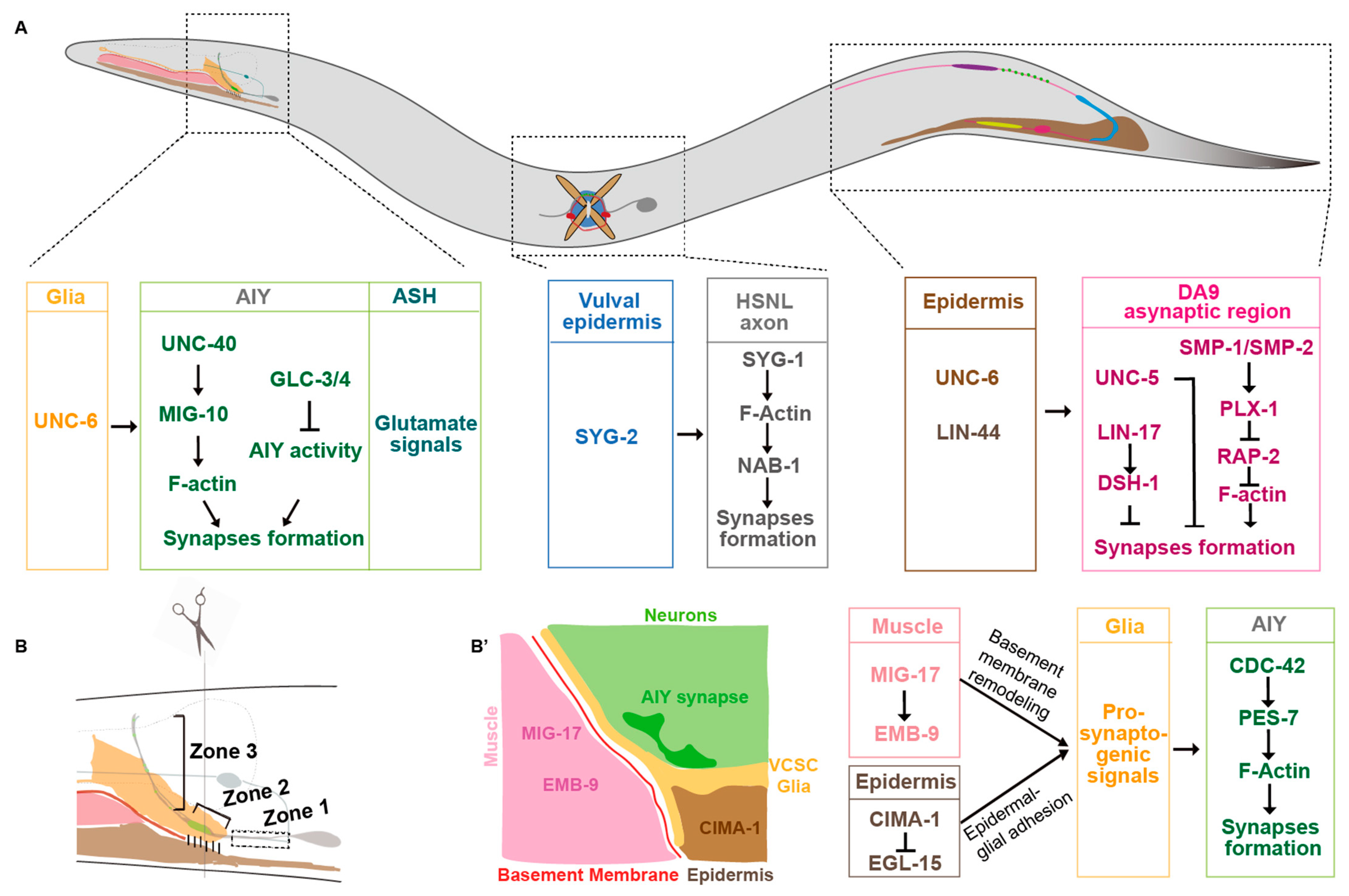
2.1. Netrin
2.2. Semaphorins
2.3. Wnts
2.4. Slit/Robo
3. Cell Adhesion/Transmembrane Molecules
4. Intracellular Molecules
5. Neuronal Activity-Dependent Mechanism
6. Glia
7. Non-Neural Cells
8. Conclusions and the Remaining Questions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hendi, A.; Kurashina, M.; Mizumoto, K. Intrinsic and extrinsic mechanisms of synapse formation and specificity in C. elegans. Cell. Mol. Life Sci. 2019, 76, 2719–2738. [Google Scholar] [CrossRef] [PubMed]
- Calahorro, F.; Izquierdo, P.G. The presynaptic machinery at the synapse of C. elegans. Invertebr. Neurosci. 2018, 18, 4. [Google Scholar] [CrossRef] [PubMed]
- Jung, N.; Haucke, V. Clathrin-mediated endocytosis at synapses. Traffic 2007, 8, 1129–1136. [Google Scholar] [CrossRef]
- Sudhof, T.C. The synaptic vesicle cycle: A cascade of protein-protein interactions. Nature 1995, 375, 645–653. [Google Scholar] [CrossRef]
- Sudhof, T.C. The synaptic vesicle cycle. Annu. Rev. Neurosci. 2004, 27, 509–547. [Google Scholar] [CrossRef] [PubMed]
- Holtmaat, A.; Svoboda, K. Experience-dependent structural synaptic plasticity in the mammalian brain. Nat. Rev. Neurosci. 2009, 10, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Christensen, R.; Shao, Z.; Colón-Ramos, D.A. The cell biology of synaptic specificity during development. Curr. Opin. Neurobiol. 2013, 23, 1018–1026. [Google Scholar] [CrossRef]
- Huang, Z.J. Subcellular organization of GABAergic synapses: Role of ankyrins and L1 cell adhesion molecules. Nat. Neurosci. 2006, 9, 163–166. [Google Scholar] [CrossRef]
- Sanes, J.R.; Yamagata, M. Many Paths to Synaptic Specificity. Annu. Rev. Cell Dev. Biol. 2009, 25, 161–195. [Google Scholar] [CrossRef]
- Shen, K.; Scheiffele, P. Genetics and Cell Biology of Building Specific Synaptic Connectivity. Annu. Rev. Neurosci. 2010, 33, 473–507. [Google Scholar] [CrossRef]
- Yogev, S.; Shen, K. Cellular and molecular mechanisms of synaptic specificity. Annu. Rev. Cell Dev. Biol. 2014, 30, 417–437. [Google Scholar] [CrossRef]
- Peters, A.; Feldman, M.L. The projection of the lateral geniculate nucleus to area 17 of the rat cerebral cortex. I. General description. J. Neurocytol. 1976, 5, 63–84. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.J.; Kalinski, C.A.; Hobert, O. Neuronal contact predicts connectivity in the C. elegans brain. Curr. Biol. 2023, 33, 2315–2320.e2. [Google Scholar] [CrossRef] [PubMed]
- Rees, C.L.; Moradi, K.; Ascoli, G.A. Weighing the Evidence in Peters’ Rule: Does Neuronal Morphology Predict Connectivity? Trends Neurosci. 2017, 40, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J. Synaptic configuration and reconfiguration in the neocortex are spatiotemporally selective. Anat. Sci. Int. 2024, 99, 17–33. [Google Scholar] [CrossRef]
- Dickson, B.J. Molecular mechanisms of axon guidance. Science 2002, 298, 1959–1964. [Google Scholar] [CrossRef]
- Kolodkin, A.L.; Tessier-Lavigne, M. Mechanisms and molecules of neuronal wiring: A primer. Cold Spring Harb. Perspect. Biol. 2011, 3, a001727. [Google Scholar] [CrossRef]
- Silhankova, M.; Korswagen, H.C. Migration of neuronal cells along the anterior-posterior body axis of C. elegans: Wnts are in control. Curr. Opin. Genet. Dev. 2007, 17, 320–325. [Google Scholar] [CrossRef]
- Stoeckli, E.T. Understanding axon guidance: Are we nearly there yet? Development 2018, 145, dev151415. [Google Scholar] [CrossRef]
- Colón-Ramos, D.A.; Margeta, M.A.; Kang, S. Glia promote local synaptogenesis through UNC-6 (netrin) signaling in C. elegans. Science 2007, 318, 103–106. [Google Scholar] [CrossRef]
- Klassen, M.P.; Shen, K. Wnt signaling positions neuromuscular connectivity by inhibiting synapse formation in C. elegans. Cell 2007, 130, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Mizumoto, K.; Shen, K. Interaxonal interaction defines tiled presynaptic innervation in C. elegans. Neuron 2013, 77, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Sales, E.C.; Heckman, E.L.; Warren, T.L.; Doe, C.Q. Regulation of subcellular dendritic synapse specificity by axon guidance cues. eLife 2019, 8, e43478. [Google Scholar] [CrossRef]
- Blockus, H.; Rolotti, S.V.; Szoboszlay, M.; Peze-Heidsieck, E.; Ming, T.; Schroeder, A.; Apostolo, N.; Vennekens, K.M.; Katsamba, P.S.; Bahna, F.; et al. Synaptogenic activity of the axon guidance molecule Robo2 underlies hippocampal circuit function. Cell Rep. 2021, 37, 109828. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.S.; Rubio, M.E.; Clem, R.L.; Johnson, D.; Case, L.; Tessier-Lavigne, M.; Huganir, R.L.; Ginty, D.D.; Kolodkin, A.L. Secreted semaphorins control spine distribution and morphogenesis in the postnatal CNS. Nature 2009, 462, 1065–1069. [Google Scholar] [CrossRef]
- Hedgecock, E.M.; Culotti, J.G.; Hall, D.H. The unc-5, unc-6, and unc-40 genes guide circumferential migrations of pioneer axons and mesodermal cells on the epidermis in C. elegans. Neuron 1990, 4, 61–85. [Google Scholar] [CrossRef]
- Keino-Masu, K.; Masu, M.; Hinck, L.; Leonardo, E.D.; Chan, S.S.; Culotti, J.G.; Tessier-Lavigne, M. Deleted in Colorectal Cancer (DCC) encodes a netrin receptor. Cell 1996, 87, 175–185. [Google Scholar] [CrossRef]
- Kennedy, T.E.; Serafini, T.; de la Torre, J.R.; Tessier-Lavigne, M. Netrins are diffusible chemotropic factors for commissural axons in the embryonic spinal cord. Cell 1994, 78, 425–435. [Google Scholar] [CrossRef]
- Leonardo, E.D.; Hinck, L.; Masu, M.; Keino-Masu, K.; Ackerman, S.L.; Tessier-Lavigne, M. Vertebrate homologues of C. elegans UNC-5 are candidate netrin receptors. Nature 1997, 386, 833–838. [Google Scholar] [CrossRef]
- Mitchell, K.J.; Doyle, J.L.; Serafini, T.; Kennedy, T.E.; Tessier-Lavigne, M.; Goodman, C.S.; Dickson, B.J. Genetic analysis of Netrin genes in Drosophila: Netrins guide CNS commissural axons and peripheral motor axons. Neuron 1996, 17, 203–215. [Google Scholar] [CrossRef]
- Serafini, T.; Colamarino, S.A.; Leonardo, E.D.; Wang, H.; Beddington, R.; Skarnes, W.C.; Tessier-Lavigne, M. Netrin-1 is required for commissural axon guidance in the developing vertebrate nervous system. Cell 1996, 87, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Serafini, T.; Kennedy, T.E.; Galko, M.J.; Mirzayan, C.; Jessell, T.M.; Tessier-Lavigne, M. The netrins define a family of axon outgrowth-promoting proteins homologous to C. elegans UNC-6. Cell 1994, 78, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Vosberg, D.E.; Leyton, M.; Flores, C. The Netrin-1/DCC guidance system: Dopamine pathway maturation and psychiatric disorders emerging in adolescence. Mol. Psychiatry 2020, 25, 297–307. [Google Scholar] [CrossRef]
- Wadsworth, W.G.; Bhatt, H.; Hedgecock, E.M. Neuroglia and pioneer neurons express UNC-6 to provide global and local netrin cues for guiding migrations in C. elegans. Neuron 1996, 16, 35–46. [Google Scholar] [CrossRef]
- Manitt, C.; Nikolakopoulou, A.M.; Almario, D.R.; Nguyen, S.A.; Cohen-Cory, S. Netrin participates in the development of retinotectal synaptic connectivity by modulating axon arborization and synapse formation in the developing brain. J. Neurosci. 2009, 29, 11065–11077. [Google Scholar] [CrossRef][Green Version]
- Nelson, J.C.; Colon-Ramos, D.A. Serotonergic neurosecretory synapse targeting is controlled by netrin-releasing guidepost neurons in Caenorhabditis elegans. J. Neurosci. 2013, 33, 1366–1376. [Google Scholar] [CrossRef]
- Poon, V.Y.; Klassen, M.P.; Shen, K. UNC-6/netrin and its receptor UNC-5 locally exclude presynaptic components from dendrites. Nature 2008, 455, 669–673. [Google Scholar] [CrossRef] [PubMed]
- White, J.G.; Southgate, E.; Thomson, J.N.; Brenner, S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philos. Trans. R. Soc. Lond. 1986, 314, 1–340. [Google Scholar]
- Goldman, J.S.; Ashour, M.A.; Magdesian, M.H.; Tritsch, N.X.; Harris, S.N.; Christofi, N.; Chemali, R.; Stern, Y.E.; Thompson-Steckel, G.; Gris, P.; et al. Netrin-1 promotes excitatory synaptogenesis between cortical neurons by initiating synapse assembly. J. Neurosci. 2013, 33, 17278–17289. [Google Scholar] [CrossRef]
- Cline, M.M.; Juarez, B.; Hunker, A.; Regiarto, E.G.; Hariadi, B.; Soden, M.E.; Zweifel, L.S. Netrin-1 regulates the balance of synaptic glutamate signaling in the adult ventral tegmental area. eLife 2023, 12, e83760. [Google Scholar] [CrossRef]
- Goodman, C.S.; Kolodkin, A.L.; Luo, Y.; Püschel, A.W.; Raper, J.A. Unified nomenclature for the semaphorins/collapsins. Semaphorin Nomenclature Committee. Cell 1999, 97, 551–552. [Google Scholar] [CrossRef] [PubMed]
- Limoni, G.; Niquille, M. Semaphorins and Plexins in central nervous system patterning: The key to it all? Curr. Opin. Neurobiol. 2021, 66, 224–232. [Google Scholar] [CrossRef]
- White, J.G.; Southgate, E.; Thomson, J.N.; Brenner, S. The structure of the ventral nerve cord of Caenorhabditis elegans. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1976, 275, 327–348. [Google Scholar] [CrossRef]
- Chen, X.; Shibata, A.C.; Hendi, A.; Kurashina, M.; Fortes, E.; Weilinger, N.L.; MacVicar, B.A.; Murakoshi, H.; Mizumoto, K. Rap2 and TNIK control Plexin-dependent tiled synaptic innervation in C. elegans. eLife 2018, 7, e38801. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, R.L.; Nguyen-Ba-Charvet, K.T.; Parray, A.; Badea, T.C.; Chedotal, A.; Kolodkin, A.L. Transmembrane semaphorin signalling controls laminar stratification in the mammalian retina. Nature 2011, 470, 259–263. [Google Scholar] [CrossRef]
- Adel, S.S.; Pranske, Z.J.; Kowalski, T.F.; Kanzler, N.; Ray, R.; Carmona, C.; Paradis, S. Plexin-B1 and Plexin-B2 play non-redundant roles in GABAergic synapse formation. bioRxiv 2023. [Google Scholar] [CrossRef]
- Duan, Y.; Wang, S.H.; Song, J.; Mironova, Y.; Ming, G.L.; Kolodkin, A.L.; Giger, R.J. Semaphorin 5A inhibits synaptogenesis in early postnatal- and adult-born hippocampal dentate granule cells. eLife 2014, 3, e04390. [Google Scholar] [CrossRef]
- Kuzirian, M.S.; Moore, A.R.; Staudenmaier, E.K.; Friedel, R.H.; Paradis, S. The class 4 semaphorin Sema4D promotes the rapid assembly of GABAergic synapses in rodent hippocampus. J. Neurosci. 2013, 33, 8961–8973. [Google Scholar] [CrossRef]
- McDermott, J.E.; Goldblatt, D.; Paradis, S. Class 4 Semaphorins and Plexin-B receptors regulate GABAergic and glutamatergic synapse development in the mammalian hippocampus. Mol. Cell. Neurosci. 2018, 92, 50–66. [Google Scholar] [CrossRef]
- Ciani, L.; Salinas, P.C. WNTs in the vertebrate nervous system: From patterning to neuronal connectivity. Nat. Rev. Neurosci. 2005, 6, 351–362. [Google Scholar] [CrossRef]
- Zwamborn, R.A.J.; Snijders, C.; An, N.; Thomson, A.; Rutten, B.P.F.; de Nijs, L. Wnt Signaling in the Hippocampus in Relation to Neurogenesis, Neuroplasticity, Stress and Epigenetics. Prog. Mol. Biol. Transl. Sci. 2018, 158, 129–157. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.A.; Khan, M.; V, M.; Kar, D. The Molecular Basis of Wnt/β-Catenin Signaling Pathways in Neurodegenerative Diseases. Int. J. Cell Biol. 2023, 2023, 9296092. [Google Scholar] [CrossRef]
- Dickins, E.M.; Salinas, P.C. Wnts in action: From synapse formation to synaptic maintenance. Front. Cell. Neurosci. 2013, 7, 162. [Google Scholar] [CrossRef]
- Fazel Darbandi, S.; Robinson Schwartz, S.E.; Pai, E.L.; Everitt, A.; Turner, M.L.; Cheyette, B.N.R.; Willsey, A.J.; State, M.W.; Sohal, V.S.; Rubenstein, J.L.R. Enhancing WNT Signaling Restores Cortical Neuronal Spine Maturation and Synaptogenesis in Tbr1 Mutants. Cell Rep. 2020, 31, 107495. [Google Scholar] [CrossRef]
- Mizumoto, K.; Shen, K. Two Wnts instruct topographic synaptic innervation in C. elegans. Cell Rep. 2013, 5, 389–396. [Google Scholar] [CrossRef]
- Davis, E.K.; Zou, Y.; Ghosh, A. Wnts acting through canonical and noncanonical signaling pathways exert opposite effects on hippocampal synapse formation. Neural Dev. 2008, 3, 32. [Google Scholar] [CrossRef]
- Park, M.; Shen, K. WNTs in synapse formation and neuronal circuitry. EMBO J. 2012, 31, 2697–2704. [Google Scholar] [CrossRef]
- Shi, Y.; Li, Q.; Shao, Z. Wnts Promote Synaptic Assembly Through T-Cell Specific Transcription Factors in Caenorhabditis elegans. Front. Mol. Neurosci. 2018, 11, 194. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.C.; Lucas, F.R.; Salinas, P.C. Axonal remodeling and synaptic differentiation in the cerebellum is regulated by WNT-7a signaling. Cell 2000, 100, 525–535. [Google Scholar] [CrossRef]
- Lucas, F.R.; Salinas, P.C. WNT-7a induces axonal remodeling and increases synapsin I levels in cerebellar neurons. Dev. Biol. 1997, 192, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Ciani, L.; Boyle, K.A.; Dickins, E.; Sahores, M.; Anane, D.; Lopes, D.M.; Gibb, A.J.; Salinas, P.C. Wnt7a signaling promotes dendritic spine growth and synaptic strength through Ca2+/Calmodulin-dependent protein kinase II. Proc. Natl. Acad. Sci. USA 2011, 108, 10732–10737. [Google Scholar] [CrossRef] [PubMed]
- Farias, G.G.; Alfaro, I.E.; Cerpa, W.; Grabowski, C.P.; Godoy, J.A.; Bonansco, C.; Inestrosa, N.C. Wnt-5a/JNK signaling promotes the clustering of PSD-95 in hippocampal neurons. J. Biol. Chem. 2009, 284, 15857–15866. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, D.; Lindsay, C.B.; Gonzalez-Billault, C.; Inestrosa, N.C. Wnt5a modulates dendritic spine dynamics through the regulation of Cofilin via small Rho GTPase activity in hippocampal neurons. J. Neurochem. 2021, 158, 673–693. [Google Scholar] [CrossRef] [PubMed]
- Hendi, A.; Niu, L.G.; Snow, A.W.; Ikegami, R.; Wang, Z.W.; Mizumoto, K. Channel-independent function of UNC-9/Innexin in spatial arrangement of GABAergic synapses in C. elegans. eLife 2022, 11, e80555. [Google Scholar] [CrossRef] [PubMed]
- Brose, K.; Bland, K.S.; Wang, K.H.; Arnott, D.; Henzel, W.; Goodman, C.S.; Tessier-Lavigne, M.; Kidd, T. Slit proteins bind Robo receptors and have an evolutionarily conserved role in repulsive axon guidance. Cell 1999, 96, 795–806. [Google Scholar] [CrossRef]
- Kidd, T.; Bland, K.S.; Goodman, C.S. Slit Is the Midline Repellent for the Robo Receptor in Drosophila. Cell 1999, 96, 785–794. [Google Scholar] [CrossRef]
- Kidd, T.; Brose, K.; Mitchell, K.J.; Fetter, R.D.; Tessier-Lavigne, M.; Goodman, C.S.; Tear, G. Roundabout controls axon crossing of the CNS midline and defines a novel subfamily of evolutionarily conserved guidance receptors. Cell 1998, 92, 205–215. [Google Scholar] [CrossRef]
- Blockus, H.; Chédotal, A. Slit-Robo signaling. Development 2016, 143, 3037–3044. [Google Scholar] [CrossRef]
- Kim, M.; Lee, C.H.; Barnum, S.J.; Watson, R.C.; Li, J.; Mastick, G.S. Slit/Robo signals prevent spinal motor neuron emigration by organizing the spinal cord basement membrane. Dev. Biol. 2019, 455, 449–457. [Google Scholar] [CrossRef]
- Mastick, G.S.; Farmer, W.T.; Altick, A.L.; Nural, H.F.; Dugan, J.P.; Kidd, T.; Charron, F. Longitudinal axons are guided by Slit/Robo signals from the floor plate. Cell Adhes. Migr. 2010, 4, 337–341. [Google Scholar] [CrossRef][Green Version]
- Kim, H.N.; Ruan, Y.; Ogana, H.; Kim, Y.M. Cadherins, Selectins, and Integrins in CAM-DR in Leukemia. Front. Oncol. 2020, 10, 592733. [Google Scholar] [CrossRef]
- De Arce, K.P.; Ribic, A.; Chowdhury, D.; Watters, K.; Thompson, G.J.; Sanganahalli, B.G.; Lippard, E.T.C.; Rohlmann, A.; Strittmatter, S.M.; Missler, M.; et al. Concerted roles of LRRTM1 and SynCAM 1 in organizing prefrontal cortex synapses and cognitive functions. Nat. Commun. 2023, 14, 459. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, X.J.; Yang, J.; Li, S.; Li, L.; Lv, X.; Ma, J.; Shi, S.H. Developmental neuronal origin regulates neocortical map formation. Cell Rep. 2023, 42, 112170. [Google Scholar] [CrossRef]
- Sudhof, T.C. The cell biology of synapse formation. J. Cell Biol. 2021, 220, e202103052. [Google Scholar] [CrossRef]
- Dalva, M.B.; McClelland, A.C.; Kayser, M.S. Cell adhesion molecules: Signalling functions at the synapse. Nat. Rev. Neurosci. 2007, 8, 206–220. [Google Scholar] [CrossRef]
- Galindo, S.E.; Shin, G.J.; Millard, S.S.; Grueber, W.B. Regulated alternative splicing of Dscam2 is required for somatosensory circuit wiring. bioRxiv 2023. [Google Scholar] [CrossRef]
- Gerrow, K.; El-Husseini, A. Cell adhesion molecules at the synapse. Front. Biosci. 2006, 11, 2400–2419. [Google Scholar] [CrossRef][Green Version]
- Obst-Pernberg, K.; Redies, C. Cadherins and synaptic specificity. J. Neurosci. Res. 1999, 58, 130–138. [Google Scholar] [CrossRef]
- Prigge, C.L.; Dembla, M.; Sharma, A.; El-Quessny, M.; Kozlowski, C.; Paisley, C.E.; Miltner, A.M.; Johnson, T.M.; Della Santina, L.; Feller, M.B.; et al. Rejection of inappropriate synaptic partners in mouse retina mediated by transcellular FLRT2-UNC5 signaling. Dev. Cell 2023, 58, 2080–2096.e7. [Google Scholar] [CrossRef] [PubMed]
- Sytnyk, V.; Leshchyns’ka, I.; Schachner, M. Neural Cell Adhesion Molecules of the Immunoglobulin Superfamily Regulate Synapse Formation, Maintenance, and Function. Trends Neurosci. 2017, 40, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.; Dombrovski, M.; Mirshahidi, P.; Nern, A.; LoCascio, S.A.; Zipursky, S.L.; Kurmangaliyev, Y.Z. Brain wiring determinants uncovered by integrating connectomes and transcriptomes. Curr. Biol. 2023, 33, 3998–4005.e6. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.H.; Walker, D.I.; Lin, Y.; Smith, M.R.; Mandl, K.D.; Jones, D.P.; Kong, S.W. Association between Neuroligin-1 polymorphism and plasma glutamine levels in individuals with autism spectrum disorder. eBioMedicine 2023, 95, 104746. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, X. SynCAMs in Normal Vertebrate Neural Development and Neuropsychiatric Disorders: From the Perspective of the OCAs. Mol. Neurobiol. 2023. [Google Scholar] [CrossRef]
- Biederer, T.; Sara, Y.; Mozhayeva, M.; Atasoy, D.; Liu, X.; Kavalali, E.T.; Südhof, T.C. SynCAM, a synaptic adhesion molecule that drives synapse assembly. Science 2002, 297, 1525–1531. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, M.; Weiner, J.A.; Sanes, J.R. Sidekicks: Synaptic Adhesion Molecules that Promote Lamina-Specific Connectivity in the Retina. Cell 2002, 110, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.; Bargmann, C.I. The immunoglobulin superfamily protein SYG-1 determines the location of specific synapses in C. elegans. Cell 2003, 112, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.; Fetter, R.D.; Bargmann, C.I. Synaptic Specificity Is Generated by the Synaptic Guidepost Protein SYG-2 and Its Receptor, SYG-1. Cell 2004, 116, 869–881. [Google Scholar] [CrossRef]
- Chia, P.H.; Patel, M.R.; Shen, K. NAB-1 instructs synapse assembly by linking adhesion molecules and F-actin to active zone proteins. Nat. Neurosci. 2012, 15, 234–242. [Google Scholar] [CrossRef]
- Ango, F.; Cristo, G.d.; Higashiyama, H.; Bennett, V.; Wu, P.; Huang, Z.J. Ankyrin-Based Subcellular Gradient of Neurofascin, an Immunoglobulin Family Protein, Directs GABAergic Innervation at Purkinje Axon Initial Segment. Cell 2004, 119, 257–272. [Google Scholar] [CrossRef]
- Telley, L.; Cadilhac, C.; Cioni, J.M.; Saywell, V.; Jahannault-Talignani, C.; Huettl, R.E.; Sarrailh-Faivre, C.; Dayer, A.; Huber, A.B.; Ango, F. Dual Function of NRP1 in Axon Guidance and Subcellular Target Recognition in Cerebellum. Neuron 2016, 91, 1276–1291. [Google Scholar] [CrossRef]
- Tai, Y.; Gallo, N.B.; Wang, M.; Yu, J.R.; Van Aelst, L. Axo-axonic Innervation of Neocortical Pyramidal Neurons by GABAergic Chandelier Cells Requires AnkyrinG-Associated L1CAM. Neuron 2019, 102, 358–372.e9. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Maness, P.F. Perisomatic GABAergic innervation in prefrontal cortex is regulated by ankyrin interaction with the L1 cell adhesion molecule. Cereb. Cortex 2010, 20, 2684–2693. [Google Scholar] [CrossRef]
- Ashrafi, S.; Betley, J.N.; Comer, J.D.; Brenner-Morton, S.; Bar, V.; Shimoda, Y.; Watanabe, K.; Peles, E.; Jessell, T.M.; Kaltschmidt, J.A. Neuronal Ig/Caspr Recognition Promotes the Formation of Axoaxonic Synapses in Mouse Spinal Cord. Neuron 2014, 81, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Betley, J.N.; Wright, C.V.; Kawaguchi, Y.; Erdelyi, F.; Szabo, G.; Jessell, T.M.; Kaltschmidt, J.A. Stringent specificity in the construction of a GABAergic presynaptic inhibitory circuit. Cell 2009, 139, 161–174. [Google Scholar] [CrossRef]
- Dong, X.; Jin, S.; Shao, Z. Glia Promote Synaptogenesis through an IQGAP PES-7 in C. elegans. Cell Rep. 2020, 30, 2614–2626.e2. [Google Scholar] [CrossRef]
- Stavoe, A.K.H.; Colon-Ramos, D.A. Netrin instructs synaptic vesicle clustering through Rac GTPase, MIG-10, and the actin cytoskeleton. J. Cell Biol. 2012, 197, 75–88. [Google Scholar] [CrossRef]
- Favuzzi, E.; Deogracias, R.; Marques-Smith, A.; Maeso, P.; Jezequel, J.; Exposito-Alonso, D.; Balia, M.; Kroon, T.; Hinojosa, A.J.; Maraver, E.F.; et al. Distinct molecular programs regulate synapse specificity in cortical inhibitory circuits. Science 2019, 363, 413–417. [Google Scholar] [CrossRef]
- Anderson, G.R.; Maxeiner, S.; Sando, R.; Tsetsenis, T.; Malenka, R.C.; Südhof, T.C. Postsynaptic adhesion GPCR latrophilin-2 mediates target recognition in entorhinal-hippocampal synapse assembly. J. Cell Biol. 2017, 216, 3831–3846. [Google Scholar] [CrossRef]
- Sando, R.; Jiang, X.; Südhof, T.C. Latrophilin GPCRs direct synapse specificity by coincident binding of FLRTs and teneurins. Science 2019, 363, eaav7969. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, A.; Vanderlinden, J.; Vints, K.; Ribeiro, L.F.; Vennekens, K.M.; Gounko, N.V.; Wierda, K.D.; de Wit, J. A Modular Organization of LRR Protein-Mediated Synaptic Adhesion Defines Synapse Identity. Neuron 2018, 99, 329–344.e7. [Google Scholar] [CrossRef]
- Exposito-Alonso, D.; Osório, C.; Bernard, C.; Pascual-García, S.; del Pino, I.; Marín, O.; Rico, B. Subcellular sorting of neuregulins controls the assembly of excitatory-inhibitory cortical circuits. eLife 2020, 9, e57000. [Google Scholar] [CrossRef]
- Lin, W.; Burgess, R.W.; Dominguez, B.; Pfaff, S.L.; Sanes, J.R.; Lee, K.F. Distinct roles of nerve and muscle in postsynaptic differentiation of the neuromuscular synapse. Nature 2001, 410, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Arber, S.; William, C.; Li, L.; Tanabe, Y.; Jessell, T.M.; Birchmeier, C.; Burden, S.J. Patterning of muscle acetylcholine receptor gene expression in the absence of motor innervation. Neuron 2001, 30, 399–410. [Google Scholar] [CrossRef]
- Yang, X.; Li, W.; Prescott, E.D.; Burden, S.J.; Wang, J.C. DNA topoisomerase IIbeta and neural development. Science 2000, 287, 131–134. [Google Scholar] [CrossRef]
- Kim, N.; Burden, S.J. MuSK controls where motor axons grow and form synapses. Nat. Neurosci. 2008, 11, 19–27. [Google Scholar] [CrossRef]
- Glock, C.; Heumüller, M.; Schuman, E.M. mRNA transport & local translation in neurons. Curr. Opin. Neurobiol. 2017, 45, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.E.; Martin, K.C.; Schuman, E.M. Local translation in neurons: Visualization and function. Nat. Struct. Mol. Biol. 2019, 26, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.C.; Zukin, R.S. RNA trafficking and local protein synthesis in dendrites: An overview. J. Neurosci. 2006, 26, 7131–7134. [Google Scholar] [CrossRef]
- Luhmann, H.J.; Khazipov, R. Neuronal activity patterns in the developing barrel cortex. Neuroscience 2018, 368, 256–267. [Google Scholar] [CrossRef]
- Penn, A.A. Early Brain Wiring: Activity-Dependent Processes. Schizophr. Bull. 2001, 27, 337–347. [Google Scholar] [CrossRef]
- Spitzer, N.C. Electrical activity in early neuronal development. Nature 2006, 444, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Wamsley, B.; Fishell, G. Genetic and activity-dependent mechanisms underlying interneuron diversity. Nat. Rev. Neurosci. 2017, 18, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Bramham, C.R.; Messaoudi, E. BDNF function in adult synaptic plasticity: The synaptic consolidation hypothesis. Prog. Neurobiol. 2005, 76, 99–125. [Google Scholar] [CrossRef]
- Golovin, R.M.; Broadie, K. Developmental experience-dependent plasticity in the first synapse of the Drosophila olfactory circuit. J. Neurophysiol. 2016, 116, 2730–2738. [Google Scholar] [CrossRef] [PubMed]
- Grunwald Kadow, I.C. State-dependent plasticity of innate behavior in fruit flies. Curr. Opin. Neurobiol. 2019, 54, 60–65. [Google Scholar] [CrossRef]
- Haas, K.F.; Broadie, K. Roles of ubiquitination at the synapse. Biochim. Biophys. Acta 2008, 1779, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Hart, M.P.; Hobert, O. Neurexin controls plasticity of a mature, sexually dimorphic neuron. Nature 2018, 553, 165–170. [Google Scholar] [CrossRef]
- Mansvelder, H.D.; Verhoog, M.B.; Goriounova, N.A. Synaptic plasticity in human cortical circuits: Cellular mechanisms of learning and memory in the human brain? Curr. Opin. Neurobiol. 2019, 54, 186–193. [Google Scholar] [CrossRef]
- Neves, G.; Cooke, S.F.; Bliss, T.V. Synaptic plasticity, memory and the hippocampus: A neural network approach to causality. Nat. Rev. Neurosci. 2008, 9, 65–75. [Google Scholar] [CrossRef]
- Sachse, S.; Rueckert, E.; Keller, A.; Okada, R.; Tanaka, N.K.; Ito, K.; Vosshall, L.B. Activity-dependent plasticity in an olfactory circuit. Neuron 2007, 56, 838–850. [Google Scholar] [CrossRef]
- Zhao, H.; Nonet, M.L. A retrograde signal is involved in activity-dependent remodeling at a C. elegans neuromuscular junction. Development 2000, 127, 1253–1266. [Google Scholar] [CrossRef] [PubMed]
- Bloodgood, B.L.; Sharma, N.; Browne, H.A.; Trepman, A.Z.; Greenberg, M.E. The activity-dependent transcription factor NPAS4 regulates domain-specific inhibition. Nature 2013, 503, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Witvliet, D.; Wu, M.; Kang, L.; Shao, Z. Temperature regulates synaptic subcellular specificity mediated by inhibitory glutamate signaling. PLoS Genet. 2021, 17, e1009295. [Google Scholar] [CrossRef] [PubMed]
- Bosworth, A.P.; Allen, N.J. The diverse actions of astrocytes during synaptic development. Curr. Opin. Neurobiol. 2017, 47, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Farizatto, K.L.G.; Baldwin, K.T. Astrocyte-synapse interactions during brain development. Curr. Opin. Neurobiol. 2023, 80, 102704. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Watanabe, S.; Christensen, R.; Jorgensen, E.M.; Colon-Ramos, D.A. Synapse location during growth depends on glia location. Cell 2013, 154, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Ango, F.; Wu, C.; Van der Want, J.J.; Wu, P.; Schachner, M.; Huang, Z.J.; Ghosh, A. Bergmann Glia and the Recognition Molecule CHL1 Organize GABAergic Axons and Direct Innervation of Purkinje Cell Dendrites. PLoS Biol. 2008, 6, e103. [Google Scholar] [CrossRef]
- Reshef, R.; Kudryavitskaya, E.; Shani-Narkiss, H.; Isaacson, B.; Rimmerman, N.; Mizrahi, A.; Yirmiya, R. The role of microglia and their CX3CR1 signaling in adult neurogenesis in the olfactory bulb. eLife 2017, 6, e30809. [Google Scholar] [CrossRef]
- Baalman, K.; Marin, M.A.; Ho, T.S.; Godoy, M.; Cherian, L.; Robertson, C.; Rasband, M.N. Axon initial segment-associated microglia. J. Neurosci. 2015, 35, 2283–2292. [Google Scholar] [CrossRef]
- Favuzzi, E.; Huang, S.; Saldi, G.A.; Binan, L.; Ibrahim, L.A.; Fernandez-Otero, M.; Cao, Y.; Zeine, A.; Sefah, A.; Zheng, K.; et al. GABA-receptive microglia selectively sculpt developing inhibitory circuits. Cell 2021, 184, 4048–4063.e32. [Google Scholar] [CrossRef]
- Fan, J.; Ji, T.; Wang, K.; Huang, J.; Wang, M.; Manning, L.; Dong, X.; Shi, Y.; Zhang, X.; Shao, Z.; et al. A muscle-epidermis-glia signaling axis sustains synaptic specificity during allometric growth in Caenorhabditis elegans. eLife 2020, 9, e55890. [Google Scholar] [CrossRef]
- Chisholm, A.D.; Hardin, J. Epidermal morphogenesis. WormBook; The C. elegans Research Community, Ed.; WormBook Research Community: New York, NY, USA, 2005; pp. 1–22. [Google Scholar] [CrossRef]
- Chisholm, A.D.; Hsiao, T.I. The Caenorhabditis elegans epidermis as a model skin. I: Development, patterning, and growth. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 861–878. [Google Scholar] [CrossRef] [PubMed]
- Vuong-Brender, T.T.; Yang, X.; Labouesse, M.C. elegans Embryonic Morphogenesis. Curr. Top. Dev. Biol. 2016, 116, 597–616. [Google Scholar] [CrossRef]
- Shi, Y.; Qin, L.; Wu, M.; Zheng, J.; Xie, T.; Shao, Z. Gut neuroendocrine signaling regulates synaptic assembly in C. elegans. EMBO Rep. 2022, 23, e53267. [Google Scholar] [CrossRef] [PubMed]
- Apóstolo, N.; de Wit, J. Compartmentalized distributions of neuronal and glial cell-surface proteins pattern the synaptic network. Curr. Opin. Neurobiol. 2019, 57, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Qi, Y.B. Building stereotypic connectivity: Mechanistic insights into structural plasticity from C. elegans. Curr. Opin. Neurobiol. 2018, 48, 97–105. [Google Scholar] [CrossRef]
- Kurup, N.; Jin, Y. Neural circuit rewiring: Insights from DD synapse remodeling. Worm 2016, 5, e1129486. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Fan, J.; Shao, Z. Cellular and Molecular Mechanisms Underlying Synaptic Subcellular Specificity. Brain Sci. 2024, 14, 155. https://doi.org/10.3390/brainsci14020155
Wang M, Fan J, Shao Z. Cellular and Molecular Mechanisms Underlying Synaptic Subcellular Specificity. Brain Sciences. 2024; 14(2):155. https://doi.org/10.3390/brainsci14020155
Chicago/Turabian StyleWang, Mengqing, Jiale Fan, and Zhiyong Shao. 2024. "Cellular and Molecular Mechanisms Underlying Synaptic Subcellular Specificity" Brain Sciences 14, no. 2: 155. https://doi.org/10.3390/brainsci14020155
APA StyleWang, M., Fan, J., & Shao, Z. (2024). Cellular and Molecular Mechanisms Underlying Synaptic Subcellular Specificity. Brain Sciences, 14(2), 155. https://doi.org/10.3390/brainsci14020155