

Effects of Repetitive Transcranial Magnetic Stimulation on Pallidum GABAergic Neurons and Motor Function in Rat Models of Kernicterus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
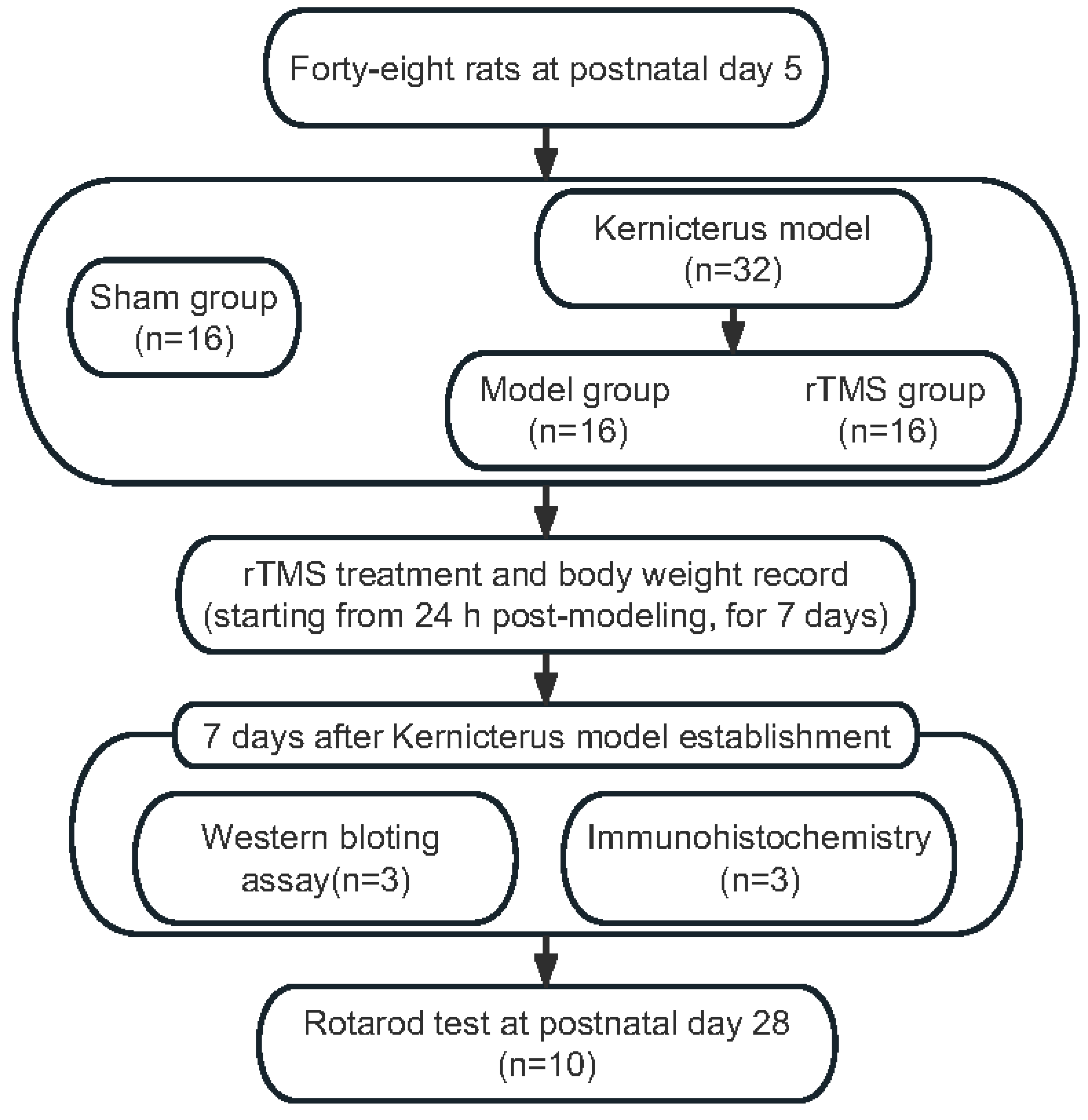
2.1. Animals
2.2. Kernicterus Model
2.3. rTMS
2.4. Western Blotting (WB)
2.5. Immunohistochemistry
2.6. Body Weight
2.7. Rotarod Test
2.8. Statistical Analysis
3. Results
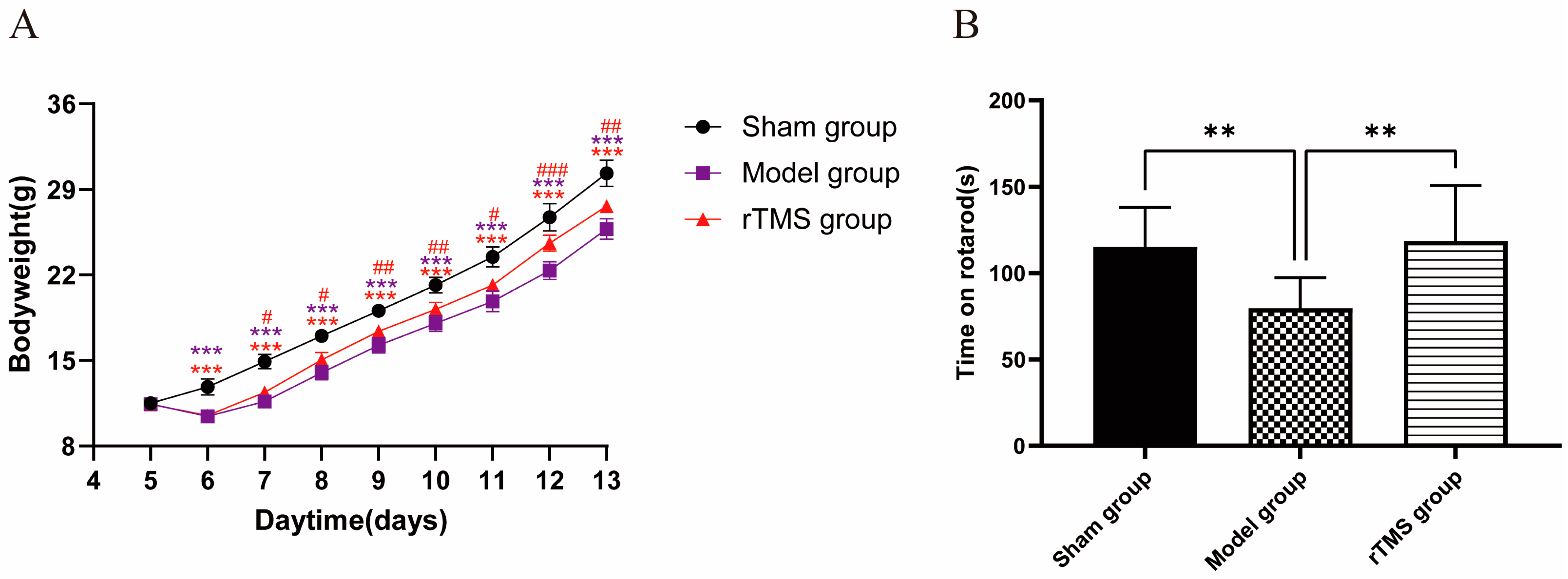
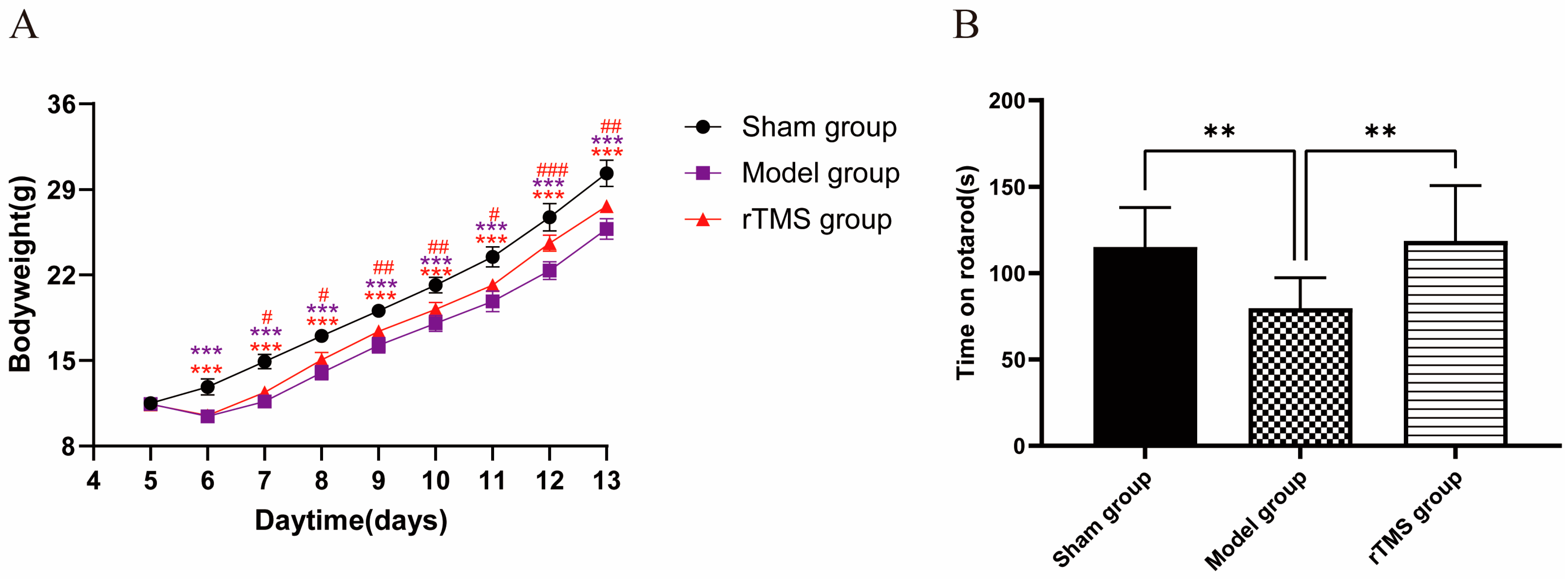
3.1. rTMS Improved the General Health and Motor Ability of Rats with Kernicterus
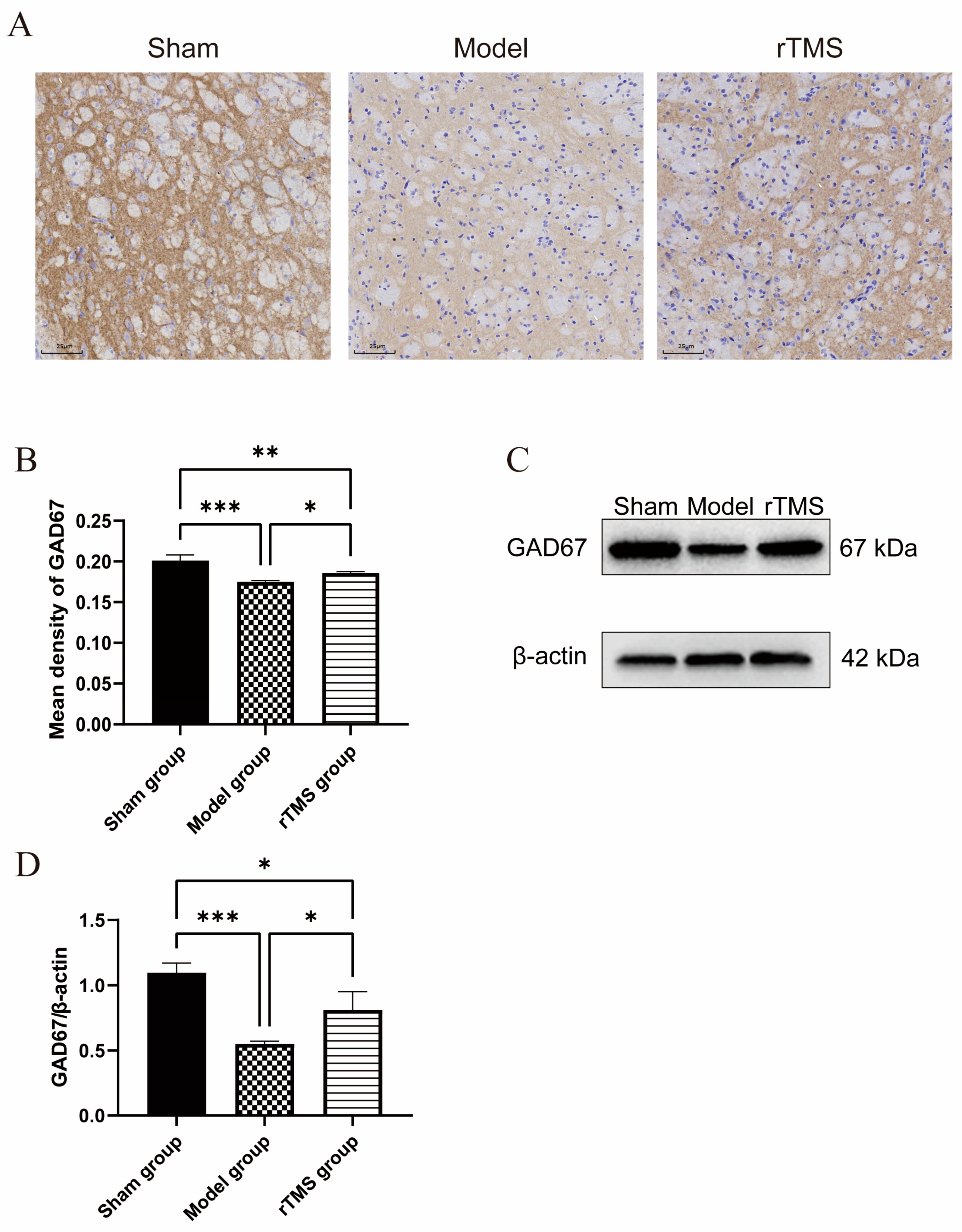
3.2. rTMS Increased the Expression of GAD67 in the Globi Pallidi of Rats with Kernicterus
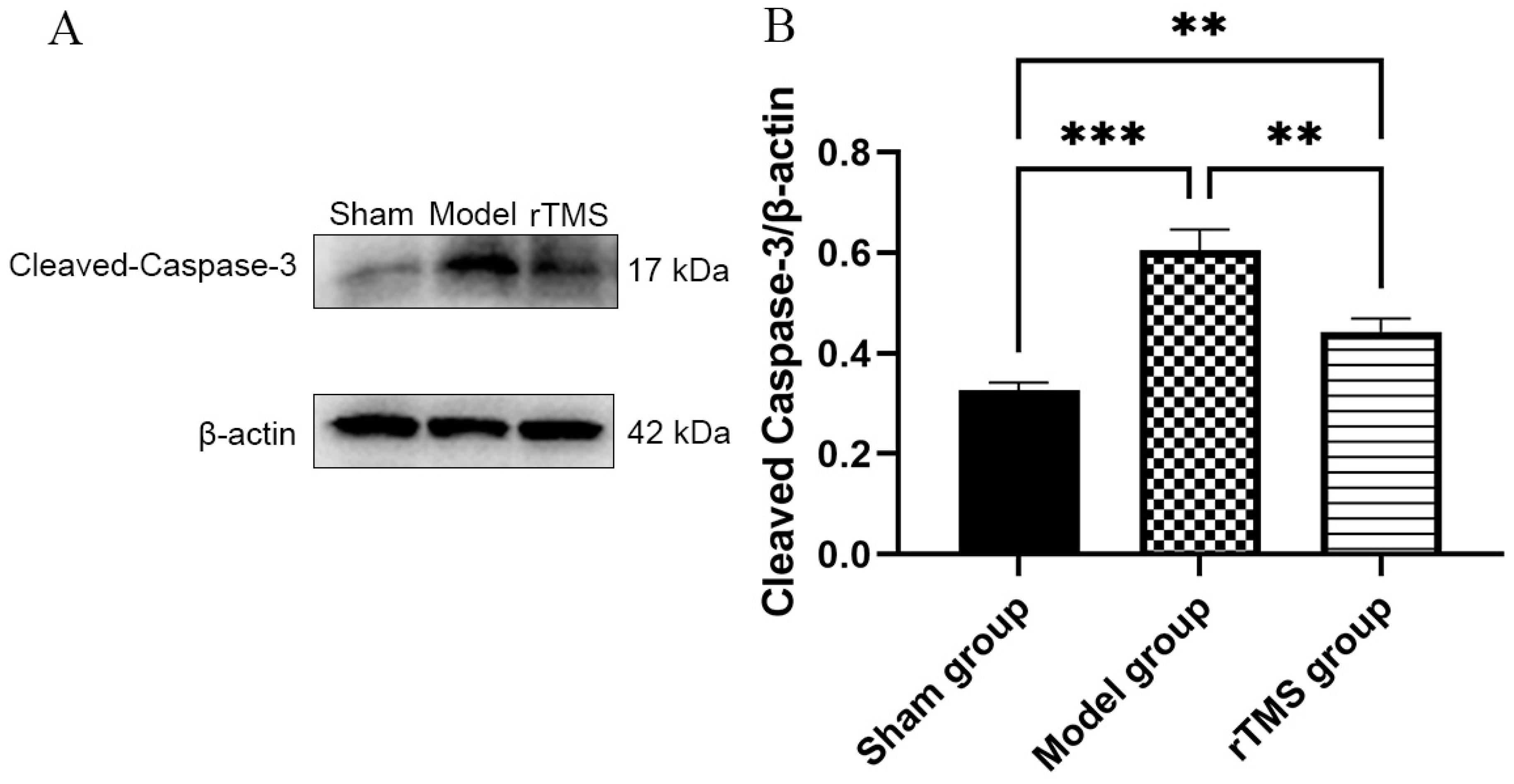
3.3. rTMS Inhibited the Activation of Caspase-3
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitra, S.; Rennie, J. Neonatal jaundice: Aetiology, diagnosis and treatment. Br. J. Hosp. Med. 2017, 78, 699–704. [Google Scholar] [CrossRef]
- Kuzniewicz, M.W.; Escobar, G.J.; Wi, S.; Liljestrand, P.; McCulloch, C.; Newman, T.B. Risk factors for severe hyperbilirubinemia among infants with borderline bilirubin levels: A nested case-control study. J. Pediatr. 2008, 153, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Watchko, J.F.; Tiribelli, C. Bilirubin-induced neurologic damage-mechanisms and management approaches. N. Engl. J. Med. 2013, 369, 2021–2030. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.; Hammerman, C. Understanding severe hyperbilirubinemia and preventing kernicterus: Adjuncts in the interpretation of neonatal serum bilirubin. Clin. Chim. Acta 2005, 356, 9–21. [Google Scholar] [CrossRef]
- Gottimukkala, S.B.; Lobo, L.; Gautham, K.S.; Bolisetty, S.; Fiander, M.; Schindler, T. Intermittent phototherapy versus continuous phototherapy for neonatal jaundice. Cochrane Database Syst. Rev. 2023, 3, Cd008168. [Google Scholar] [CrossRef]
- Olusanya, B.O.; Ogunlesi, T.A.; Slusher, T.M. Why is kernicterus still a major cause of death and disability in low-income and middle-income countries? Arch. Dis. Child. 2014, 99, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Kasirer, Y.; Kaplan, M.; Hammerman, C. Kernicterus on the Spectrum. NeoReviews 2023, 24, e329–e342. [Google Scholar] [CrossRef]
- Shapiro, S.M. Chronic bilirubin encephalopathy: Diagnosis and outcome. Semin. Fetal Neonatal Med. 2010, 15, 157–163. [Google Scholar] [CrossRef]
- Riordan, S.M.; Shapiro, S.M. Review of bilirubin neurotoxicity I: Molecular biology and neuropathology of disease. Pediatr. Res. 2020, 87, 327–331. [Google Scholar] [CrossRef]
- Hegeman, D.J.; Hong, E.S.; Hernandez, V.M.; Chan, C.S. The external globus pallidus: Progress and perspectives. Eur. J. Neurosci. 2016, 43, 1239–1265. [Google Scholar] [CrossRef]
- Mallet, N.; Micklem, B.R.; Henny, P.; Brown, M.T.; Williams, C.; Bolam, J.P.; Nakamura, K.C.; Magill, P.J. Dichotomous organization of the external globus pallidus. Neuron 2012, 74, 1075–1086. [Google Scholar] [CrossRef]
- Gazzin, S.; Zelenka, J.; Zdrahalova, L.; Konickova, R.; Zabetta, C.C.; Giraudi, P.J.; Berengeno, A.L.; Raseni, A.; Robert, M.C.; Vitek, L.; et al. Bilirubin accumulation and Cyp mRNA expression in selected brain regions of jaundiced Gunn rat pups. Pediatr. Res. 2012, 71, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.M.; Sola, S.; Brites, D. Bilirubin induces apoptosis via the mitochondrial pathway in developing rat brain neurons. Hepatology 2002, 35, 1186–1195. [Google Scholar] [CrossRef]
- Vodret, S.; Bortolussi, G.; Iaconcig, A.; Martinelli, E.; Tiribelli, C.; Muro, A.F. Attenuation of neuro-inflammation improves survival and neurodegeneration in a mouse model of severe neonatal hyperbilirubinemia. Brain Behav. Immun. 2018, 70, 166–178. [Google Scholar] [CrossRef]
- Smith, K.; Leyden, J.J. Safety of doxycycline and minocycline: A systematic review. Clin. Ther. 2005, 27, 1329–1342. [Google Scholar] [CrossRef]
- Shapiro, S.M.; Riordan, S.M. Review of bilirubin neurotoxicity II: Preventing and treating acute bilirubin encephalopathy and kernicterus spectrum disorders. Pediatr. Res. 2020, 87, 332–337. [Google Scholar] [CrossRef]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A.; Safety of TMS Consensus Group. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [PubMed]
- Medina, F.J.; Tunez, I. Mechanisms and pathways underlying the therapeutic effect of transcranial magnetic stimulation. Rev. Neurosci. 2013, 24, 507–525. [Google Scholar] [CrossRef] [PubMed]
- Aguera, E.; Caballero-Villarraso, J.; Feijoo, M.; Escribano, B.M.; Conde, C.; Bahamonde, M.C.; Giraldo, A.I.; Paz-Rojas, E.; Tunez, I. Clinical and Neurochemical Effects of Transcranial Magnetic Stimulation (TMS) in Multiple Sclerosis: A Study Protocol for a Randomized Clinical Trial. Front. Neurol. 2020, 11, 750. [Google Scholar] [CrossRef]
- Gao, F.; Wang, S.; Guo, Y.; Wang, J.; Lou, M.; Wu, J.; Ding, M.; Tian, M.; Zhang, H. Protective effects of repetitive transcranial magnetic stimulation in a rat model of transient cerebral ischaemia: A microPET study. Eur. J. Nucl. Med. Mol. Imaging 2010, 37, 954–961. [Google Scholar] [CrossRef]
- Buetefisch, C.M.; Wei, L.; Gu, X.; Epstein, C.M.; Yu, S.P. Neuroprotection of Low-Frequency Repetitive Transcranial Magnetic Stimulation after Ischemic Stroke in Rats. Ann. Neurol. 2023, 93, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Wennberg, R.P.; Hance, A.J. Experimental bilirubin encephalopathy: Importance of total bilirubin, protein binding, and blood-brain barrier. Pediatr. Res. 1986, 20, 789–792. [Google Scholar] [CrossRef]
- Song, S.; Hu, Y.; Gu, X.; Si, F.; Hua, Z. A novel newborn rat kernicterus model created by injecting a bilirubin solution into the cisterna magna. PLoS ONE 2014, 9, e96171. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Watchko, J.F. Kernicterus and the molecular mechanisms of bilirubin-induced CNS injury in newborns. Neuromolecular Med. 2006, 8, 513–529. [Google Scholar] [CrossRef]
- Das, S.; van Landeghem, F.K.H. Clinicopathological Spectrum of Bilirubin Encephalopathy/Kernicterus. Diagnostics 2019, 9, 24. [Google Scholar] [CrossRef]
- Yilmaz, Y.; Alper, G.; Kilicoglu, G.; Celik, L.; Karadeniz, L.; Yilmaz-Degirmenci, S. Magnetic resonance imaging findings in patients with severe neonatal indirect hyperbilirubinemia. J. Child Neurol. 2001, 16, 452–455. [Google Scholar] [CrossRef]
- Sugama, S.; Soeda, A.; Eto, Y. Magnetic resonance imaging in three children with kernicterus. Pediatr. Neurol. 2001, 25, 328–331. [Google Scholar] [CrossRef]
- Govaert, P.; Lequin, M.; Swarte, R.; Robben, S.; De Coo, R.; Weisglas-Kuperus, N.; De Rijke, Y.; Sinaasappel, M.; Barkovich, J. Changes in globus pallidus with (pre)term kernicterus. Pediatrics 2003, 112, 1256–1263. [Google Scholar] [CrossRef] [PubMed]
- Hachiya, Y.; Hayashi, M. Bilirubin encephalopathy: A study of neuronal subpopulations and neurodegenerative mechanisms in 12 autopsy cases. Brain Dev. 2008, 30, 269–278. [Google Scholar] [CrossRef]
- Johnston, M.V.; Hoon, A.H., Jr. Possible mechanisms in infants for selective basal ganglia damage from asphyxia, kernicterus, or mitochondrial encephalopathies. J. Child Neurol. 2000, 15, 588–591. [Google Scholar] [CrossRef]
- Rawat, V.; Bortolussi, G.; Gazzin, S.; Tiribelli, C.; Muro, A.F. Bilirubin-Induced Oxidative Stress Leads to DNA Damage in the Cerebellum of Hyperbilirubinemic Neonatal Mice and Activates DNA Double-Strand Break Repair Pathways in Human Cells. Oxidative Med. Cell. Longev. 2018, 2018, 1801243. [Google Scholar] [CrossRef]
- Brito, M.A.; Vaz, A.R.; Silva, S.L.; Falcao, A.S.; Fernandes, A.; Silva, R.F.; Brites, D. N-methyl-aspartate receptor and neuronal nitric oxide synthase activation mediate bilirubin-induced neurotoxicity. Mol. Med. 2010, 16, 372–380. [Google Scholar] [CrossRef]
- Ostrow, J.D.; Pascolo, L.; Brites, D.; Tiribelli, C. Molecular basis of bilirubin-induced neurotoxicity. Trends Mol. Med. 2004, 10, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Watchko, J.F. Bilirubin-Induced Neurotoxicity in the Preterm Neonate. Clin. Perinatol. 2016, 43, 297–311. [Google Scholar] [CrossRef]
- Qian, S.; Kumar, P.; Testai, F.D. Bilirubin Encephalopathy. Curr. Neurol. Neurosci. Rep. 2022, 22, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.M.P.; Solá, S.; Castro, R.E.; Laires, P.A.; Brites, D.; Moura, J.J.G. Perturbation of membrane dynamics in nerve cells as an early event during bilirubin-induced apoptosis. J. Lipid Res. 2002, 43, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Zorzo, C.; Higarza, S.G.; Mendez, M.; Martinez, J.A.; Pernia, A.M.; Arias, J.L. High frequency repetitive transcranial magnetic stimulation improves neuronal activity without affecting astrocytes and microglia density. Brain Res. Bull. 2019, 150, 13–20. [Google Scholar] [CrossRef]
- Hong, Y.; Lyu, J.; Zhu, L.; Wang, X.; Peng, M.; Chen, X.; Deng, Q.; Gao, J.; Yuan, Z.; Wang, D.; et al. High-frequency repetitive transcranial magnetic stimulation (rTMS) protects against ischemic stroke by inhibiting M1 microglia polarization through let-7b-5p/HMGA2/NF-kappaB signaling pathway. BMC Neurosci. 2022, 23, 49. [Google Scholar] [CrossRef]
- Lefaucheur, J.P. Principles of therapeutic use of transcranial and epidural cortical stimulation. Clin. Neurophysiol. 2008, 119, 2179–2184. [Google Scholar] [CrossRef]
- Cho, S.S.; Strafella, A.P. rTMS of the left dorsolateral prefrontal cortex modulates dopamine release in the ipsilateral anterior cingulate cortex and orbitofrontal cortex. PLoS ONE 2009, 4, e6725. [Google Scholar] [CrossRef]
- Yue, L.; Xiao-lin, H.; Tao, S. The effects of chronic repetitive transcranial magnetic stimulation on glutamate and gamma-aminobutyric acid in rat brain. Brain Res. 2009, 1260, 94–99. [Google Scholar] [CrossRef]
- Müller, M.B.; Toschi, N.; Kresse, A.E.; Post, A.; Keck, M.E. Long-term repetitive transcranial magnetic stimulation increases the expression of brain-derived neurotrophic factor and cholecystokinin mRNA, but not neuropeptide tyrosine mRNA in specific areas of rat brain. Neuropsychopharmacology 2000, 23, 205–215. [Google Scholar] [CrossRef]
- Lang, U.E.; Hellweg, R.; Gallinat, J.; Bajbouj, M. Acute prefrontal cortex transcranial magnetic stimulation in healthy volunteers: No effects on brain-derived neurotrophic factor (BDNF) concentrations in serum. J. Affect. Disord. 2008, 107, 255–258. [Google Scholar] [CrossRef]
- Tunez, I.; Montilla, P.; del Carmen Munoz, M.; Medina, F.J.; Drucker-Colin, R. Effect of transcranial magnetic stimulation on oxidative stress induced by 3-nitropropionic acid in cortical synaptosomes. Neurosci. Res. 2006, 56, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Aguera, E.; Caballero-Villarraso, J.; Feijoo, M.; Escribano, B.M.; Bahamonde, M.C.; Conde, C.; Galvan, A.; Tunez, I. Impact of Repetitive Transcranial Magnetic Stimulation on Neurocognition and Oxidative Stress in Relapsing-Remitting Multiple Sclerosis: A Case Report. Front. Neurol. 2020, 11, 817. [Google Scholar] [CrossRef] [PubMed]
- Tunez, I.; Drucker-Colin, R.; Jimena, I.; Medina, F.J.; Munoz Mdel, C.; Pena, J.; Montilla, P. Transcranial magnetic stimulation attenuates cell loss and oxidative damage in the striatum induced in the 3-nitropropionic model of Huntington’s disease. J. Neurochem. 2006, 97, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Song, L.; Liu, Z. The effect of repetitive transcranial magnetic stimulation on a model rat of Parkinson’s disease. Neuroreport 2010, 21, 268–272. [Google Scholar] [CrossRef]
- Hong, Y.; Liu, Q.; Peng, M.; Bai, M.; Li, J.; Sun, R.; Guo, H.; Xu, P.; Xie, Y.; Li, Y.; et al. High-frequency repetitive transcranial magnetic stimulation improves functional recovery by inhibiting neurotoxic polarization of astrocytes in ischemic rats. J. Neuroinflammation 2020, 17, 150. [Google Scholar] [CrossRef]
- Hsieh, T.-H.; He, X.-K.; Liu, H.-H.; Chen, J.-J.J.; Peng, C.-W.; Liu, H.-L.; Rotenberg, A.; Chen, K.-T.; Chang, M.-Y.; Chiang, Y.-H.; et al. Early Repetitive Transcranial Magnetic Stimulation Exerts Neuroprotective Effects and Improves Motor Functions in Hemiparkinsonian Rats. Neural Plast. 2021, 2021, 1763533. [Google Scholar] [CrossRef]
- Dileone, M.; Profice, P.; Pilato, F.; Ranieri, F.; Capone, F.; Musumeci, G.; Florio, L.; Di Iorio, R.; Di Lazzaro, V. Repetitive transcranial magnetic stimulation for ALS. CNS Neurol. Disord. Drug Targets 2010, 9, 331–334. [Google Scholar] [CrossRef]
- Li, S.; Huang, H.; Wei, Q.; He, C.; Feng, J.; Wang, Y.; Li, M.; Zhang, Q.; Xia, X.; Hua, Z. Depression of Pyroptosis by Inhibiting Caspase-1 Activation Improves Neurological Outcomes of Kernicterus Model Rats. ACS Chem. Neurosci. 2021, 12, 2929–2939. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.H.; Fan, P.; Zhang, L.R.; Chen, C.Y.; Xu, J.; Huang, J.; Lu, W.T.; Zhu, S.J.; Qiu, G.P.; Xu, S.Y.; et al. Neuroglobin expression and function in the temporal cortex of bilirubin encephalopathy rats. Anat. Rec. 2022, 305, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Villarraso, J.; Medina, F.J.; Escribano, B.M.; Agüera, E.; Santamaría, A.; Pascual-Leone, A.; Túnez, I. Mechanisms Involved in Neuroprotective Effects of Transcranial Magnetic Stimulation. CNS Neurol. Disord. Drug Targets 2022, 21, 557–573. [Google Scholar] [CrossRef]
- Hernandez, V.M.; Hegeman, D.J.; Cui, Q.; Kelver, D.A.; Fiske, M.P.; Glajch, K.E.; Pitt, J.E.; Huang, T.Y.; Justice, N.J.; Chan, C.S. Parvalbumin+ Neurons and Npas1+ Neurons Are Distinct Neuron Classes in the Mouse External Globus Pallidus. J. Neurosci. 2015, 35, 11830–11847. [Google Scholar] [CrossRef] [PubMed]
- Abdi, A.; Mallet, N.; Mohamed, F.Y.; Sharott, A.; Dodson, P.D.; Nakamura, K.C.; Suri, S.; Avery, S.V.; Larvin, J.T.; Garas, F.N.; et al. Prototypic and arkypallidal neurons in the dopamine-intact external globus pallidus. J. Neurosci. 2015, 35, 6667–6688. [Google Scholar] [CrossRef]
- Kleppner, S.R.; Tobin, A.J. GABA signalling: Therapeutic targets for epilepsy, Parkinson’s disease and Huntington’s disease. Expert Opin. Ther. Targets 2001, 5, 219–239. [Google Scholar] [CrossRef]
- Gajcy, K.; Lochyński, S.; Librowski, T. A role of GABA analogues in the treatment of neurological diseases. Curr. Med. Chem. 2010, 17, 2338–2347. [Google Scholar] [CrossRef]
- Galvan, A.; Wichmann, T. GABAergic circuits in the basal ganglia and movement disorders. Prog. Brain Res. 2007, 160, 287–312. [Google Scholar] [CrossRef]
- Haridas, A.; Tagliati, M.; Osborn, I.; Isaias, I.; Gologorsky, Y.; Bressman, S.B.; Weisz, D.; Alterman, R.L. Pallidal deep brain stimulation for primary dystonia in children. Neurosurgery 2011, 68, 738–743. [Google Scholar] [CrossRef]
- Sanger, T.D.; Liker, M.; Arguelles, E.; Deshpande, R.; Maskooki, A.; Ferman, D.; Tongol, A.; Robison, A. Pediatric Deep Brain Stimulation Using Awake Recording and Stimulation for Target Selection in an Inpatient Neuromodulation Monitoring Unit. Brain Sci. 2018, 8, 135. [Google Scholar] [CrossRef]
- Baysal, B.; Tüzün, F.; Yücesoy, E.; Özbal, S.; Uğur, B.E.; Sonmez, A.; Olgun, Y.; Kirkim, G.; Evin, H.; Duman, N.; et al. Topiramate and darbepoetin alpha ameliorate bilirubin-induced neuronal cell damage. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 3585–3593. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, N.; Jia, Y.; Zhou, X.; Wang, X.; Zhou, H.; Xiao, N. Effects of Repetitive Transcranial Magnetic Stimulation on Pallidum GABAergic Neurons and Motor Function in Rat Models of Kernicterus. Brain Sci. 2023, 13, 1252. https://doi.org/10.3390/brainsci13091252
Wang N, Jia Y, Zhou X, Wang X, Zhou H, Xiao N. Effects of Repetitive Transcranial Magnetic Stimulation on Pallidum GABAergic Neurons and Motor Function in Rat Models of Kernicterus. Brain Sciences. 2023; 13(9):1252. https://doi.org/10.3390/brainsci13091252
Chicago/Turabian StyleWang, Nanqin, Yongzhu Jia, Xuanzi Zhou, Xia Wang, Huyao Zhou, and Nong Xiao. 2023. "Effects of Repetitive Transcranial Magnetic Stimulation on Pallidum GABAergic Neurons and Motor Function in Rat Models of Kernicterus" Brain Sciences 13, no. 9: 1252. https://doi.org/10.3390/brainsci13091252
APA StyleWang, N., Jia, Y., Zhou, X., Wang, X., Zhou, H., & Xiao, N. (2023). Effects of Repetitive Transcranial Magnetic Stimulation on Pallidum GABAergic Neurons and Motor Function in Rat Models of Kernicterus. Brain Sciences, 13(9), 1252. https://doi.org/10.3390/brainsci13091252