

Altered Metabolic Networks in Mesial Temporal Lobe Epilepsy with Focal to Bilateral Seizures
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. 18F-FDG-PET Scanning
2.3. Image Preprocessing and Graph Theoretical Analyses
2.4. Statistical Analysis
3. Results
3.1. Demographic Data
3.2. Differences in SUVR Values
3.3. Brain Metabolic Network Abnormalities in MTLE Patients with FBTCS Relative to Healthy Controls
3.4. Brain Metabolic Network Abnormalities in FBTCS− Patients Relative to Healthy Controls
3.5. Differences in Brain Metabolic Networks between Patients with and without FBTCS
4. Discussion
4.1. Brain Metabolism in Patients with MTLE
4.2. Brain Metabolic Network Alterations
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blümcke, I.; Thom, M.; Aronica, E.; Armstrong, D.D.; Bartolomei, F.; Bernasconi, A.; Bernasconi, N.; Bien, C.G.; Cendes, F.; Coras, R.; et al. International consensus classification of hippocampal sclerosis in temporal lobe epilepsy: A Task Force report from the ILAE Commission on Diagnostic Methods. Epilepsia 2013, 54, 1315–1329. [Google Scholar] [CrossRef] [PubMed]
- Bone, B.; Fogarasi, A.; Schulz, R.; Gyimesi, C.; Kalmar, Z.; Kovacs, N.; Ebner, A.; Janszky, J. Secondarily generalized seizures in temporal lobe epilepsy. Epilepsia 2012, 53, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Harden, C.; Tomson, T.; Gloss, D.; Buchhalter, J.; Cross, J.H.; Donner, E.; French, J.A.; Gil-Nagel, A.; Hesdorffer, D.C.; Smithson, W.H.; et al. Practice guideline summary: Sudden unexpected death in epilepsy incidence rates and risk factors: Report of the Guideline Development, Dissemination, and Implementation Subcommittee of the American Academy of Neurology and the American Epilepsy Society. Neurology 2017, 88, 1674–1680. [Google Scholar] [CrossRef] [PubMed]
- Hemery, C.; Ryvlin, P.; Rheims, S. Prevention of generalized tonic-clonic seizures in refractory focal epilepsy: A meta-analysis. Epilepsia 2014, 55, 1789–1799. [Google Scholar] [CrossRef] [PubMed]
- Baud, M.O.; Vulliemoz, S.; Seeck, M. Recurrent secondary generalization in frontal lobe epilepsy: Predictors and a potential link to surgical outcome? Epilepsia 2015, 56, 1454–1462. [Google Scholar] [CrossRef]
- Perani, S.; Tierney, T.M.; Centeno, M.; Shamshiri, E.A.; Yaakub, S.N.; O’Muircheartaigh, J.; Carmichael, D.W.; Richardson, M.P. Thalamic volume reduction in drug-naive patients with new-onset genetic generalized epilepsy. Epilepsia 2018, 59, 226–234. [Google Scholar] [CrossRef]
- He, X.; Chaitanya, G.; Asma, B.; Caciagli, L.; Bassett, D.S.; Tracy, J.I.; Sperling, M.R. Disrupted basal ganglia–thalamocortical loops in focal to bilateral tonic-clonic seizures. Brain 2020, 143, 175–190. [Google Scholar] [CrossRef]
- Weng, Y.; Larivière, S.; Caciagli, L.; Vos de Wael, R.; Rodríguez-Cruces, R.; Royer, J.; Xu, Q.; Bernasconi, N.; Bernasconi, A.; Thomas Yeo, B.T.; et al. Macroscale and microcircuit dissociation of focal and generalized human epilepsies. Commun. Biol. 2020, 3, 244. [Google Scholar] [CrossRef]
- Chen, C.; Li, H.; Ding, F.; Yang, L.; Huang, P.; Wang, S.; Jin, B.; Xu, C.; Wang, Y.; Ding, M.; et al. Alterations in the hippocampal-thalamic pathway underlying secondarily generalized tonic-clonic seizures in mesial temporal lobe epilepsy: A diffusion tensor imaging study. Epilepsia 2019, 60, 121–130. [Google Scholar] [CrossRef]
- Blumenfeld, H.; Varghese, G.I.; Purcaro, M.J.; Motelow, J.E.; Enev, M.; McNally, K.A.; Levin, A.R.; Hirsch, L.J.; Tikofsky, R.; Zubal, I.G.; et al. Cortical and subcortical networks in human secondarily generalized tonic-clonic seizures. Brain 2009, 132 Pt 4, 999–1012. [Google Scholar] [CrossRef]
- Peng, S.-J.; Hsin, Y.-L. Altered structural and functional thalamocortical networks in secondarily generalized extratemporal lobe seizures. Neuroimage Clin. 2017, 13, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Dabrowska, N.; Joshi, S.; Williamson, J.; Lewczuk, E.; Lu, Y.; Oberoi, S.; Brodovskaya, A.; Kapur, J. Parallel pathways of seizure generalization. Brain 2019, 142, 2336–2351. [Google Scholar] [CrossRef] [PubMed]
- Bartolomei, F.; Lagarde, S.; Wendling, F.; McGonigal, A.; Jirsa, V.; Guye, M.; Bénar, C. Defining epileptogenic networks: Contribution of SEEG and signal analysis. Epilepsia 2017, 58, 1131–1147. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; An, D.; Xiao, F.; Lei, D.; Niu, R.; Li, W.; Ren, J.; Liu, W.; Tang, Y.; Zhang, L.; et al. Real-time effects of interictal spikes on hippocampus and amygdala functional connectivity in unilateral temporal lobe epilepsy: An EEG-fMRI study. Epilepsia 2019, 60, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Zhao, B.; Hu, W.; Zhang, C.; Wang, X.; Wang, Y.; Liu, C.; Mo, J.; Sang, L.; Ma, Y.; et al. Effective connectivity among the hippocampus, amygdala, and temporal neocortex in epilepsy patients: A cortico-cortical evoked potential study. Epilepsy Behav. 2021, 115, 107661. [Google Scholar] [CrossRef]
- Vaughan, D.N.; Rayner, G.; Tailby, C.; Jackson, G.D. MRI-negative temporal lobe epilepsy: A network disorder of neocortical connectivity. Neurology 2016, 87, 1934–1942. [Google Scholar] [CrossRef]
- McDonald, C.R.; Ahmadi, M.E.; Hagler, D.J.; Tecoma, E.S.; Iragui, V.J.; Gharapetian, L.; Dale, A.M.; Halgren, E. Diffusion tensor imaging correlates of memory and language impairments in temporal lobe epilepsy. Neurology 2008, 71, 1869–1876. [Google Scholar] [CrossRef]
- Bullmore, E.; Sporns, O. The economy of brain network organization. Nat. Rev. Neurosci. 2012, 13, 336–349. [Google Scholar] [CrossRef]
- Riedl, V.; Utz, L.; Castrillón, G.; Grimmer, T.; Rauschecker, J.P.; Ploner, M.; Friston, K.J.; Drzezga, A.; Sorg, C. Metabolic connectivity mapping reveals effective connectivity in the resting human brain. Proc. Natl. Acad. Sci. USA 2016, 113, 428–433. [Google Scholar] [CrossRef]
- Yakushev, I.; Drzezga, A.; Habeck, C. Metabolic connectivity: Methods and applications. Curr. Opin. Neurol. 2017, 30, 677–685. [Google Scholar] [CrossRef]
- Wang, K.-L.; Hu, W.; Liu, T.-H.; Zhao, X.-B.; Han, C.-L.; Xia, X.-T.; Zhang, J.-G.; Wang, F.; Meng, F.-G. Metabolic covariance networks combining graph theory measuring aberrant topological patterns in mesial temporal lobe epilepsy. CNS Neurosci. Ther. 2019, 25, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Doyen, M.; Chawki, M.B.; Heyer, S.; Guedj, E.; Roch, V.; Marie, P.-Y.; Tyvaert, L.; Maillard, L.; Verger, A. Metabolic connectivity is associated with seizure outcome in surgically treated temporal lobe epilepsies: A 18F-FDG PET seed correlation analysis. NeuroImage Clin. 2022, 36, 103210. [Google Scholar] [CrossRef] [PubMed]
- Sinha, N.; Peternell, N.; Schroeder, G.M.; Tisi, J.; Vos, S.B.; Winston, G.P.; Duncan, J.S.; Wang, Y.; Taylor, P.N. Focal to bilateral tonic–clonic seizures are associated with widespread network abnormality in temporal lobe epilepsy. Epilepsia 2021, 62, 729–741. [Google Scholar] [CrossRef]
- Caciagli, L.; Allen, L.A.; He, X.; Trimmel, K.; Vos, S.B.; Centeno, M.; Galovic, M.; Sidhu, M.K.; Thompson, P.J.; Bassett, D.S.; et al. Thalamus and focal to bilateral seizures: A multiscale cognitive imaging study. Neurology 2020, 95, e2427–e2441. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, I.E.; Berkovic, S.; Capovilla, G.; Connolly, M.B.; French, J.; Guilhoto, L.; Hirsch, E.; Jain, S.; Mathern, G.W.; Moshé, S.L.; et al. ILAE classification of the epilepsies: Position paper of the ILAE Commission for Classification and Terminology. Epilepsia 2017, 58, 512–521. [Google Scholar] [CrossRef]
- Ren, S.; Huang, Q.; Bao, W.; Jiang, D.; Xiao, J.; Li, J.; Xie, F.; Guan, Y.; Feng, R.; Hua, F. Metabolic Brain Network and Surgical Outcome in Temporal Lobe Epilepsy: A Graph Theoretical Study Based on 18F-fluorodeoxyglucose PET. Neuroscience 2021, 478, 39–48. [Google Scholar] [CrossRef]
- Makris, N.; Meyer, J.W.; Bates, J.F.; Yeterian, E.H.; Kennedy, D.N.; Caviness, V.S. MRI-Based topographic parcellation of human cerebral white matter and nuclei II. Rationale and applications with systematics of cerebral connectivity. Neuroimage 1999, 9, 18–45. [Google Scholar] [CrossRef]
- Mijalkov, M.; Kakaei, E.; Pereira, J.B.; Westman, E.; Volpe, G. Alzheimer’s Disease Neuroimaging Initiative BRAPH: A graph theory software for the analysis of brain connectivity. PLoS ONE 2017, 12, e0178798. [Google Scholar] [CrossRef]
- Rissman, J.; Gazzaley, A.; D’Esposito, M. Measuring functional connectivity during distinct stages of a cognitive task. Neuroimage 2004, 23, 752–763. [Google Scholar] [CrossRef]
- Zhao, B.; Seguin, C.; Ai, L.; Sun, T.; Hu, W.; Zhang, C.; Wang, X.; Liu, C.; Wang, Y.; Mo, J.; et al. Aberrant Metabolic Patterns Networks in Insular Epilepsy. Front. Neurol. 2020, 11, 605256. [Google Scholar] [CrossRef]
- Farahani, F.V.; Karwowski, W.; Lighthall, N.R. Application of Graph Theory for Identifying Connectivity Patterns in Human Brain Networks: A Systematic Review. Front. Neurosci. 2019, 13, 585. [Google Scholar] [CrossRef] [PubMed]
- Wilke, C.; Worrell, G.; He, B. Graph analysis of epileptogenic networks in human partial epilepsy. Epilepsia 2011, 52, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Blumer, A.; Lee, K. An algorithm for modularity analysis of directed and weighted biological networks based on edge-betweenness centrality. Bioinformatics 2006, 22, 3106–3108. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Lee, D.S.; Lee, S.K.; Kim, S.-K.; Chung, C.K.; Chang, K.H.; Choi, K.Y.; Chung, J.-K.; Lee, M.C. Differential features of metabolic abnormalities between medial and lateral temporal lobe epilepsy: Quantitative analysis of (18)F-FDG PET using SPM. J. Nucl. Med. 2003, 44, 1006–1012. [Google Scholar]
- Shim, H.-K.; Lee, H.-J.; Kim, S.E.; Lee, B.I.; Park, S.; Park, K.M. Alterations in the metabolic networks of temporal lobe epilepsy patients: A graph theoretical analysis using FDG-PET. NeuroImage Clin. 2020, 27, 102349. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.H.; Bleasel, A.; Wen, L.; Eberl, S.; Byth, K.; Fulham, M.; Somerville, E.; Mohamed, A. The topography and significance of extratemporal hypometabolism in refractory mesial temporal lobe epilepsy examined by FDG-PET. Epilepsia 2010, 51, 1365–1373. [Google Scholar] [CrossRef]
- Rektor, I.; Zákopcan, J.; Tyrlíková, I.; Kuba, R.; Brázdil, M.; Chrastina, J.; Novák, Z. Secondary generalization in seizures of temporal lobe origin: Ictal EEG pattern in a stereo-EEG study. Epilepsy Behav. 2009, 15, 235–239. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Farooque, P.; Chen, W.C.; Youngblood, M.W.; Zaveri, H.P.; Gerrard, J.L.; Spencer, D.D.; Hirsch, L.J.; Blumenfeld, H. Ictal spread of medial temporal lobe seizures with and without secondary generalization: An intracranial electroencephalography analysis. Epilepsia 2014, 55, 289–295. [Google Scholar] [CrossRef]
- Foldvary-Schaefer, N.; Unnwongse, K. Localizing and lateralizing features of auras and seizures. Epilepsy Behav. 2011, 20, 160–166. [Google Scholar] [CrossRef]
- Choi, J.Y.; Kim, S.J.; Hong, S.B.; Seo, D.W.; Hong, S.C.; Kim, B.-T.; Kim, S.E. Extratemporal hypometabolism on FDG PET in temporal lobe epilepsy as a predictor of seizure outcome after temporal lobectomy. Eur. J. Nucl. Med. Mol. Imaging 2003, 30, 581–587. [Google Scholar] [CrossRef]
- Van Bogaert, P.; Massager, N.; Tugendhaft, P.; Wikler, D.; Damhaut, P.; Levivier, M.; Brotchi, J.; Goldman, S. Statistical Parametric Mapping of Regional Glucose Metabolism in Mesial Temporal Lobe Epilepsy. NeuroImage 2000, 12, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Savio, A.; Fünger, S.; Tahmasian, M.; Rachakonda, S.; Manoliu, A.; Sorg, C.; Grimmer, T.; Calhoun, V.; Drzezga, A.; Riedl, V.; et al. Resting-State Networks as Simultaneously Measured with Functional MRI and PET. J. Nucl. Med. 2017, 58, 1314–1317. [Google Scholar] [CrossRef] [PubMed]
- Albert, R.; Jeong, H.; Barabasi, A.L. Error and attack tolerance of complex networks. Nature 2000, 406, 378–382. [Google Scholar] [CrossRef]
- Newman, M.E.J. Assortative mixing in networks. Phys. Rev. Lett. 2002, 89, 208701. [Google Scholar] [CrossRef] [PubMed]
- Bialonski, S.; Lehnertz, K. Assortative mixing in functional brain networks during epileptic seizures. Chaos 2013, 23, 033139. [Google Scholar] [CrossRef]
- Miller, J.W. Are Generalized Tonic–Clonic Seizures Really “Generalized”? Epilepsy Curr. 2010, 10, 80–81. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| FBTCS+ Patients | Healthy Controls | Statistic | p-Value | |
|---|---|---|---|---|
| Age, years | 25.1 ± 5.8 | 27.5 ± 8.8 | Independence Student’s t = −1.211 | 0.23 |
| Sex, male/female | 12/15 | 15/15 | χ2 = 0.176 | 0.67 |
| Age at seizure onset, years | 10.8 ± 5.9 | N/A | N/A | N/A |
| Epilepsy duration, years | 14.3 ± 6.8 | N/A | N/A | N/A |
| Handedness, left/right | 4/23 | 4/26 | χ2 = 0.026 | 0.83 |
| Side, left/right | 15/12 | N/A | N/A | N/A |
| FBTCS−Patients | Healthy Controls | Statistic | p-Value | |
| Age, years | 27.8 ± 5.7 | 27.5 ± 8.8 | Independence Student’s t = 0.139 | 0.89 |
| Sex, male/female | 13/14 | 15/15 | χ2 = 0.020 | 0.89 |
| Age at seizure onset, years | 13.8 ± 5.0 | N/A | N/A | N/A |
| Epilepsy duration, years | 14.0 ± 7.4 | N/A | N/A | N/A |
| Handedness, left/right | 3/24 | 4/26 | χ2 = 0.065 | 0.88 |
| Side, left/right | 12/15 | N/A | N/A | N/A |
| FBTCS+ Patients | FBTCS−Patients | Statistic | p-Value | |
| Age, years | 25.1 ± 5.8 | 27.8 ± 5.7 | Independence Student’s t = −1.721 | 0.09 |
| Sex, male/female | 12/15 | 13/14 | χ2 = 0.075 | 0.78 |
| Age at seizure onset, years | 10.8 ± 5.9 | 13.8 ± 5.0 | Independence Student’s t = −1.987 | 0.05 |
| Epilepsy duration, years | 14.3 ± 6.8 | 14.0 ± 7.4 | Independence Student’s t = 0.135 | 0.89 |
| Handedness, left/right | 4/23 | 3/24 | χ2 = 0.164 | >0.99 |
| Side, left/right | 15/12 | 12/15 | χ2 = 0.667 | 0.41 |
| Follow-up, years | 4.8 ± 1.5 | 4.7 ± 1.3 | Independence Student’s t = 0.070 | 0.95 |
| Engel class, I/II-IV | 24/3 | 23/4 | χ2 = 0.164 | >0.99 |
| FBTCS+ | Healthy Controls | Difference | CI Lower | CI Upper | p-Value | |
|---|---|---|---|---|---|---|
| Average degree | 110.054 | 106.875 | −3.178 | −5.106 | 6.778 | 0.276 |
| Average strength | 62.530 | 48.726 | −13.803 | −18.803 | 18.363 | 0.205 |
| Characteristic path length | 1.908 | 2.408 | 0.500 | −0.648 | 0.676 | 0.220 |
| Global efficiency | 0.578 | 0.469 | −0.109 | −0.130 | 0.133 | 0.179 |
| Local efficiency | 3.602 | 2.376 | −1.223 | −1.463 | 1.484 | 0.184 |
| Mean clustering coefficient | 0.544 | 0.422 | −0.122 | −0.173 | 0.172 | 0.291 |
| Modularity | 0.053 | 0.058 | 0.004 | −0.046 | 0.040 | 0.828 |
| Assortative coefficient | −0.045 | −0.013 | 0.032 | −0.031 | 0.027 | 0.031 * |
| Small-worldness index | 0.954 | 0.930 | −0.024 | −0.071 | 0.079 | 0.583 |
| FBTCS− | Healthy Controls | Difference | CI Lower | CI Upper | p-Value | |
| Average degree | 110.929 | 106.875 | −4.054 | −3.145 | 4.030 | 0.063 |
| Average strength | 68.707 | 48.726 | −19.981 | −18.275 | 19.237 | 0.081 |
| Characteristic path length | 1.733 | 2.408 | 0.675 | −0.599 | 0.569 | 0.057 |
| Global efficiency | 0.625 | 0.469 | −0.156 | −0.140 | 0.129 | 0.058 |
| Local efficiency | 4.191 | 2.376 | −1.815 | −1.693 | 1.729 | 0.073 |
| Mean clustering coefficient | 0.605 | 0.422 | −0.183 | −0.168 | 0.175 | 0.079 |
| Modularity | 0.035 | 0.058 | 0.023 | −0.054 | 0.053 | 0.462 |
| Assortative coefficient | −0.029 | −0.013 | 0.016 | −0.027 | 0.028 | 0.313 |
| Small-worldness index | 0.974 | 0.930 | −0.044 | −0.054 | 0.059 | 0.170 |
| FBTCS+ | FBTCS− | Difference | CI Lower | CI Upper | p-Value | |
| Average degree | 110.054 | 110.929 | 0.875 | −3.776 | 3.562 | 0.601 |
| Average strength | 62.530 | 68.707 | 6.177 | −19.774 | 20.686 | 0.614 |
| Characteristic path length | 1.908 | 1.733 | −0.175 | −0.613 | 0.592 | 0.632 |
| Global efficiency | 0.578 | 0.625 | 0.047 | −0.153 | 0.148 | 0.640 |
| Local efficiency | 3.602 | 4.191 | 0.589 | −1.741 | 1.807 | 0.637 |
| Mean clustering coefficient | 0.578 | 0.625 | 0.047 | −0.153 | 0.148 | 0.640 |
| Modularity | 0.054 | 0.035 | −0.019 | −0.043 | 0.040 | 0.521 |
| Assortative coefficient | −0.045 | −0.029 | 0.016 | −0.041 | 0.030 | 0.437 |
| Small-worldness index | 0.954 | 0.974 | 0.021 | −0.062 | 0.066 | 0.630 |
| FBTCS+ | Healthy Controls | Difference | CI Lower | CI Upper | p-Value | |
|---|---|---|---|---|---|---|
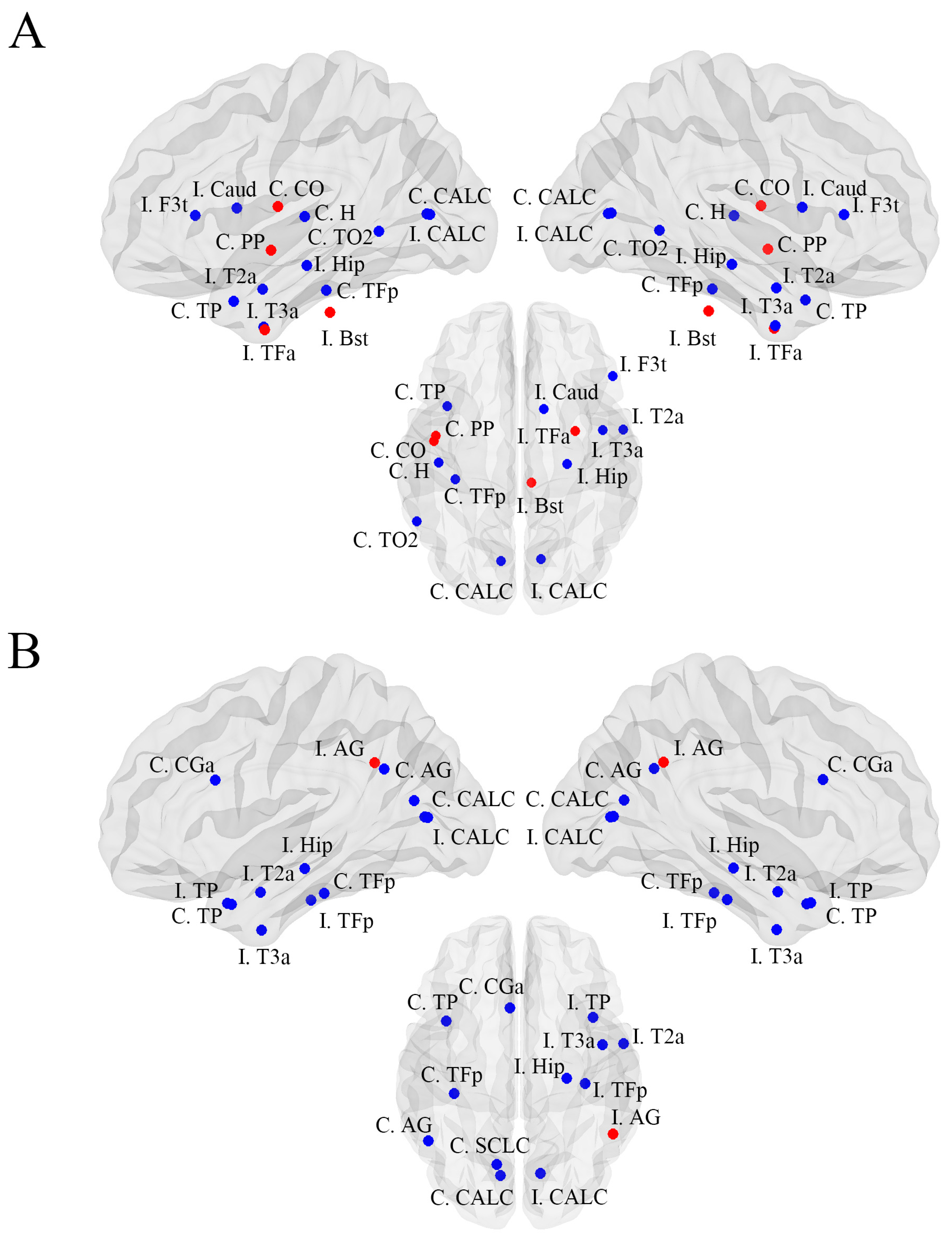
| I. Inferior Frontal Gyrus, pars triangularis | 0.000 | 0.012 | 0.012 | −0.008 | 0.007 | 0.027 |
| C. Temporal Pole | 0.000 | 0.019 | 0.019 | −0.008 | 0.007 | 0.003 |
| I. Middle Temporal Gyrus, anterior division | 0.000 | 0.002 | 0.001 | 0.000 | 0.000 | 0.005 |
| C. Middle Temporal Gyrus, temporooccipital part | 0.000 | 0.008 | 0.008 | −0.006 | 0.005 | 0.038 |
| I. Inferior Temporal Gyrus, anterior division | 0.000 | 0.002 | 0.002 | −0.001 | 0.001 | 0.021 |
| C. Intracalcarine Cortex | 0.000 | 0.011 | 0.011 | −0.001 | 0.001 | 0.001 |
| I. Intracalcarine Cortex | 0.000 | 0.003 | 0.003 | −0.002 | 0.002 | 0.032 |
| I. Temporal Fusiform Cortex, anterior division | 0.002 | 0.000 | −0.002 | −0.002 | 0.002 | 0.028 |
| C. Temporal Fusiform Cortex, posterior division | 0.000 | 0.021 | 0.021 | −0.012 | 0.010 | 0.013 |
| C. Central Opercular Cortex | 0.005 | 0.000 | −0.005 | −0.003 | 0.004 | 0.036 |
| C. Planum Polare | 0.007 | 0.000 | −0.007 | −0.003 | 0.002 | 0.013 |
| C. Heschls Gyrus | 0.000 | 0.011 | 0.011 | −0.008 | 0.007 | 0.011 |
| I. Brain-Stem | 0.003 | 0.000 | −0.003 | −0.002 | 0.001 | 0.032 |
| I. Caudate | 0.000 | 0.002 | 0.002 | −0.001 | 0.000 | 0.035 |
| I. Hippocampus | 0.000 | 0.026 | 0.026 | −0.013 | 0.012 | 0.001 |
| FBTCS− | Healthy Controls | Difference | CI Lower | CI Upper | p-Value | |
| C. Temporal Pole | 0.000 | 0.019 | 0.019 | −0.010 | 0.009 | 0.003 |
| I. Temporal Pole | 0.000 | 0.007 | 0.007 | 0.000 | 0.000 | 0.001 |
| I. Middle Temporal Gyrus, anterior division | 0.000 | 0.002 | 0.001 | 0.000 | 0.000 | 0.021 |
| I. Inferior Temporal Gyrus, anterior division | 0.000 | 0.002 | 0.002 | 0.000 | 0.000 | 0.010 |
| C. Angular Gyrus | 0.000 | 0.004 | 0.004 | −0.003 | 0.002 | 0.030 |
| I. Angular Gyrus | 0.011 | 0.000 | −0.010 | −0.006 | 0.005 | 0.024 |
| C. Intracalcarine Cortex | 0.000 | 0.011 | 0.011 | −0.002 | 0.000 | 0.001 |
| I. Intracalcarine Cortex | 0.000 | 0.003 | 0.003 | 0.000 | 0.000 | 0.003 |
| C. Cingulate Gyrus, anterior division | 0.000 | 0.018 | 0.018 | −0.010 | 0.008 | 0.016 |
| C. Temporal Fusiform Cortex, posterior division | 0.000 | 0.021 | 0.021 | −0.013 | 0.014 | 0.022 |
| I. Temporal Fusiform Cortex, posterior division | 0.000 | 0.004 | 0.004 | −0.003 | 0.003 | 0.042 |
| C. Supracalcarine Cortex | 0.000 | 0.001 | 0.001 | 0.000 | 0.000 | 0.017 |
| Rt. Hippocampus | 0.000 | 0.026 | 0.026 | −0.009 | 0.007 | 0.001 |
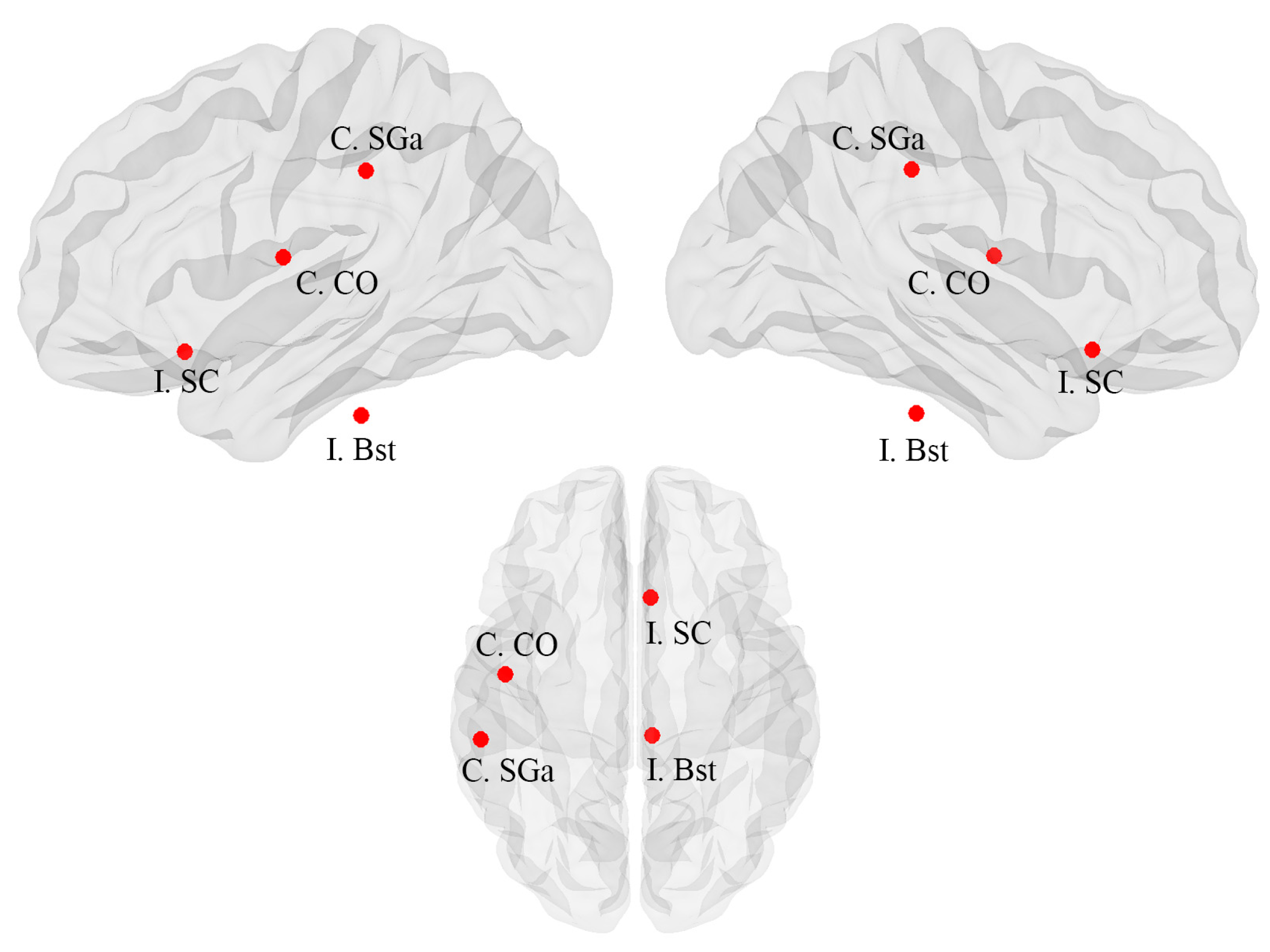
| FBTCS+ | FBTCS− | Difference | CI lower | CI upper | p-value | |
| C. Supramarginal Gyrus, anterior division | 0.020 | 0.000 | −0.020 | −0.011 | 0.013 | 0.031 |
| I. Subcallosal Cortex | 0.002 | 0.000 | −0.002 | −0.002 | 0.002 | 0.043 |
| C. Central Opercular Cortex | 0.005 | 0.000 | −0.005 | −0.003 | 0.003 | 0.039 |
| I. Brain-Stem | 0.003 | 0.000 | −0.003 | −0.004 | 0.003 | 0.029 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.; Mo, J.; Zhang, J.; Hu, W.; Zhang, C.; Wang, X.; Zhao, B.; Zhang, K. Altered Metabolic Networks in Mesial Temporal Lobe Epilepsy with Focal to Bilateral Seizures. Brain Sci. 2023, 13, 1239. https://doi.org/10.3390/brainsci13091239
Guo Z, Mo J, Zhang J, Hu W, Zhang C, Wang X, Zhao B, Zhang K. Altered Metabolic Networks in Mesial Temporal Lobe Epilepsy with Focal to Bilateral Seizures. Brain Sciences. 2023; 13(9):1239. https://doi.org/10.3390/brainsci13091239
Chicago/Turabian StyleGuo, Zhihao, Jiajie Mo, Jianguo Zhang, Wenhan Hu, Chao Zhang, Xiu Wang, Baotian Zhao, and Kai Zhang. 2023. "Altered Metabolic Networks in Mesial Temporal Lobe Epilepsy with Focal to Bilateral Seizures" Brain Sciences 13, no. 9: 1239. https://doi.org/10.3390/brainsci13091239
APA StyleGuo, Z., Mo, J., Zhang, J., Hu, W., Zhang, C., Wang, X., Zhao, B., & Zhang, K. (2023). Altered Metabolic Networks in Mesial Temporal Lobe Epilepsy with Focal to Bilateral Seizures. Brain Sciences, 13(9), 1239. https://doi.org/10.3390/brainsci13091239