
Long-Term Effects of Repetitive Transcranial Magnetic Stimulation on Tinnitus in a Guinea Pig Model

 , , and
, , and 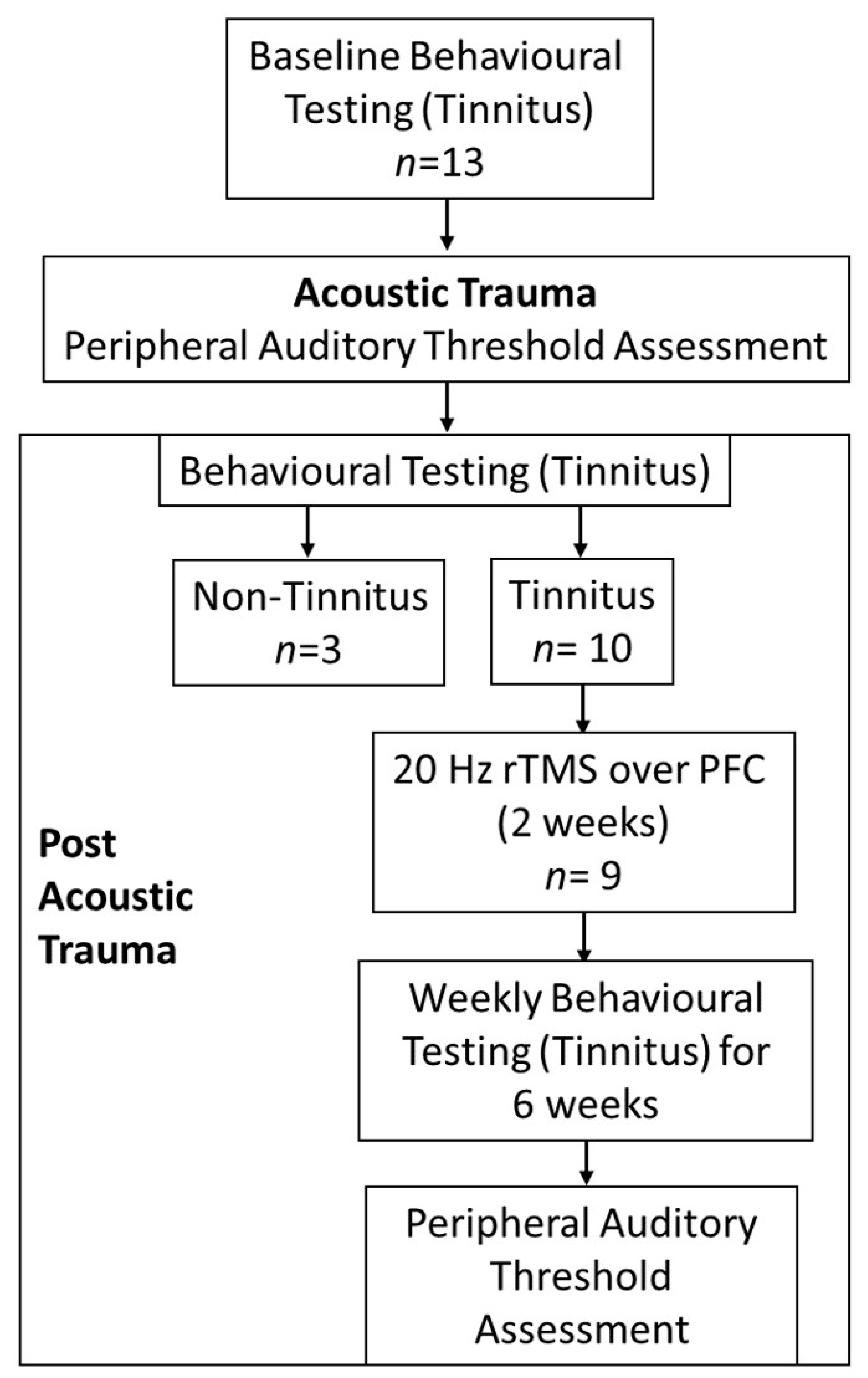{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Behavioral Assessments for Tinnitus
2.3. Acoustic Trauma Surgery
2.4. rTMS Treatment
2.5. End of Experiment
2.6. Immunohistochemistry and Analysis
2.7. Analyses
3. Results
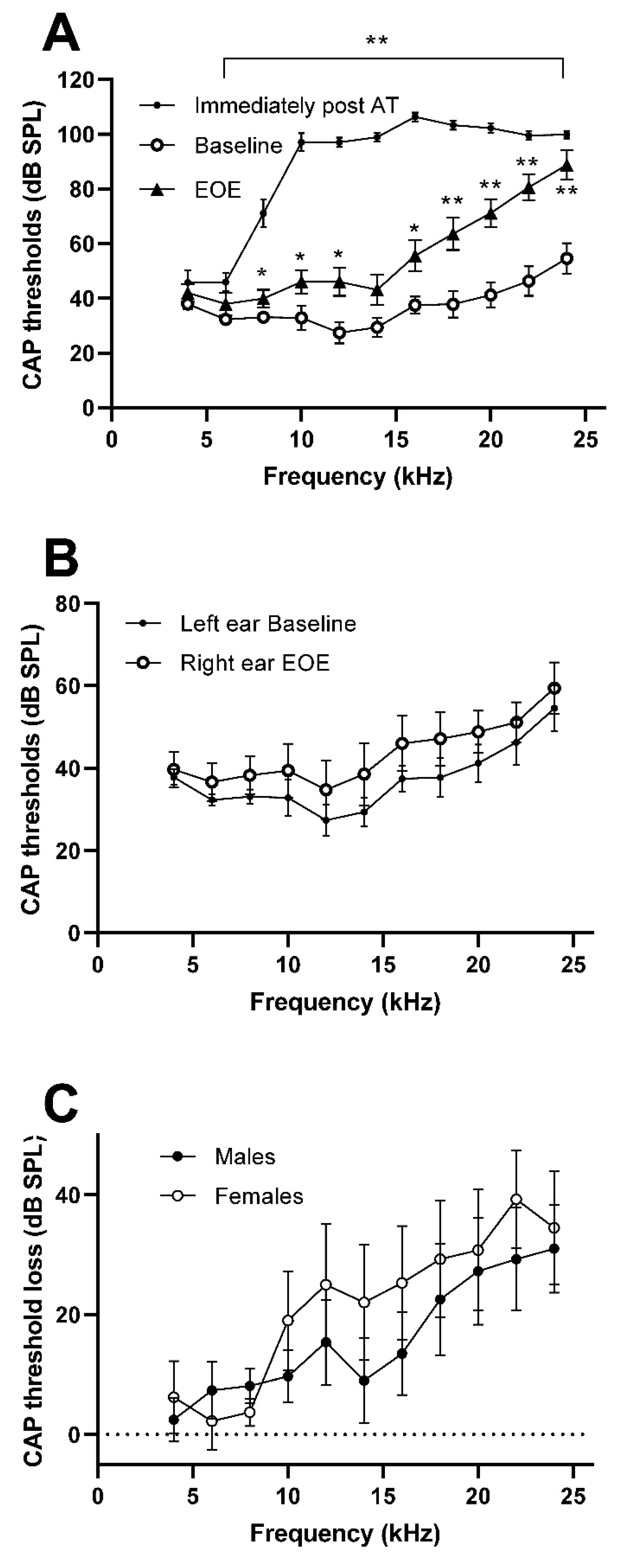
3.1. Hearing Loss
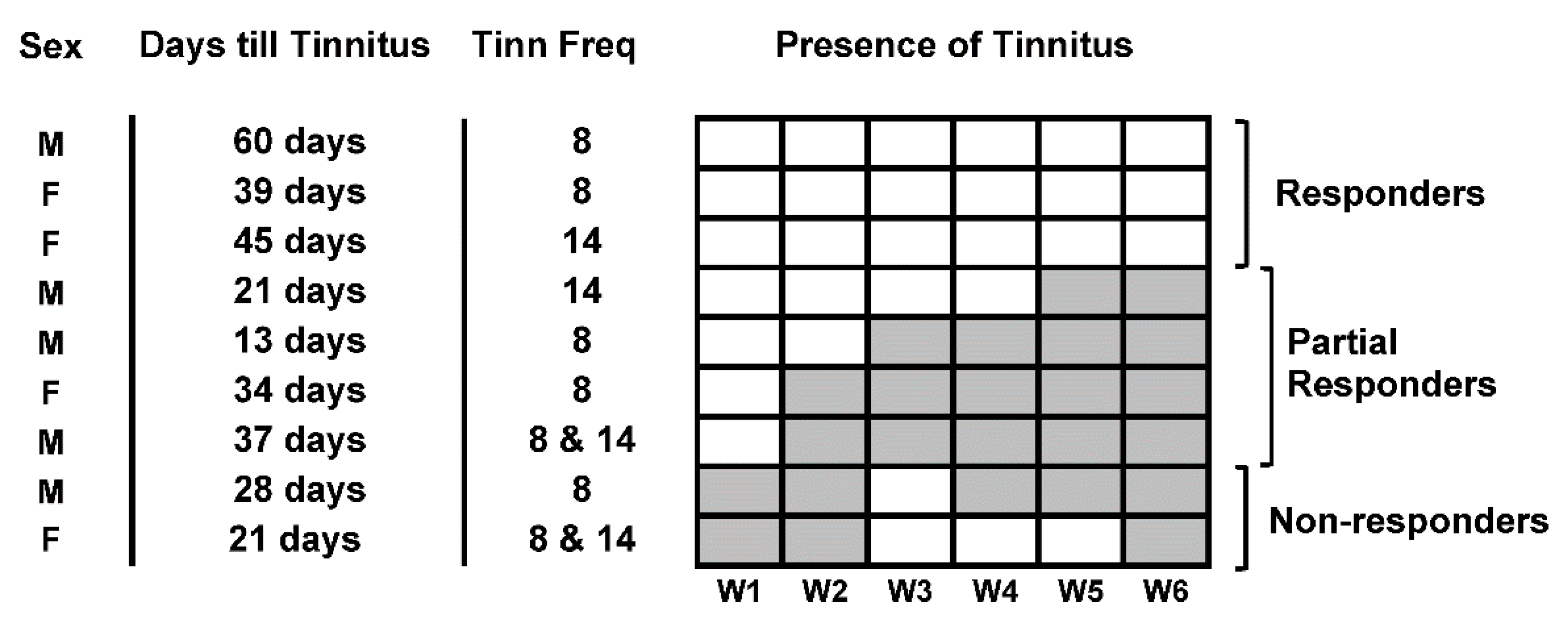
3.2. Tinnitus Development
3.3. Effect of Treatment
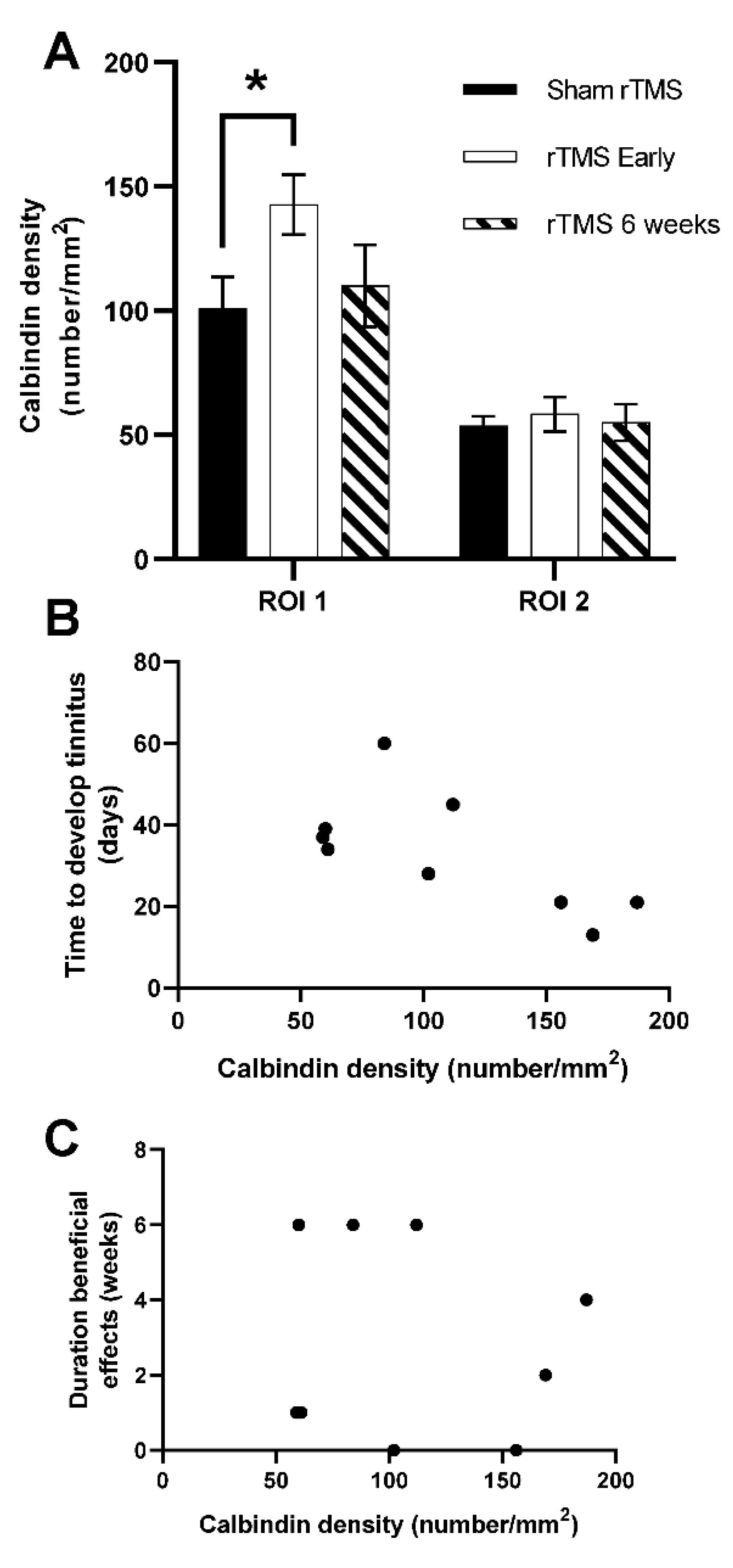
3.4. Neuronal Density of Calbindin Postivive Neurons in PFC
4. Discussion
5. Conclusion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baguley, D.; McFerran, D.; Hall, D. Tinnitus. Lancet 2013, 382, 1600–1607. [Google Scholar] [CrossRef]
- Henry, J.A.; Reavis, K.M.; Griest, S.E.; Thielman, E.J.; Theodoroff, S.M.; Grush, L.D.; Carlson, K.F. Tinnitus: An Epidemiologic Perspective. Otolaryngol. Clin. N. Am. 2020, 53, 481–499. [Google Scholar] [CrossRef] [PubMed]
- Elarbed, A.; Fackrell, K.; Bagsuley, D.M.; Hoare, D.J. Tinnitus and stress in adults: A scoping review. Int. J. Audiol. 2020, 60, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Langguth, B.; Kleinjung, T.; Landgrebe, M. Severe tinnitus and depressive symptoms: A complex interaction. Otolaryngol. Neck Surg. 2011, 145, 519. [Google Scholar] [CrossRef]
- Bhatt, J.M.; Bhattacharyya, N.; Lin, H.W. Relationships between tinnitus and the prevalence of anxiety and depression. Laryngoscope 2017, 127, 466–469. [Google Scholar] [CrossRef]
- Pattyn, T.; Van Den Eede, F.; Vanneste, S.; Cassiers, L.; Veltman, D.J.; Van De Heyning, P.; Sabbe, B.C.G. Tinnitus and anxiety disorders: A review. Hear. Res. 2016, 333, 255–265. [Google Scholar] [CrossRef]
- Tan, C.M.; Lecluyse, W.; McFerran, D.; Meddis, R. Tinnitus and patterns of hearing loss. J. Assoc. Res. Otolaryngol. 2013, 14, 275–282. [Google Scholar] [CrossRef]
- Koops, E.A.; Eggermont, J.J. The thalamus and tinnitus: Bridging the gap between animal data and findings in humans. Hear. Res. 2021, 407, 108280. [Google Scholar] [CrossRef]
- Norena, A.J.; Eggermont, J.J. Changes in spontaneous neural activity immediately after an acoustic trauma: Implications for neural correlates of tinnitus. Hear. Res. 2003, 183, 137–153. [Google Scholar] [CrossRef]
- Mulders, W.H.; Robertson, D. Hyperactivity in the auditory midbrain after acoustic trauma: Dependence on cochlear activity. Neuroscience 2009, 164, 733–746. [Google Scholar] [CrossRef]
- Kalappa, B.I.; Brozoski, T.J.; Turner, J.G.; Caspary, D.M. Single unit hyperactivity and bursting in the auditory thalamus of awake rats directly correlates with behavioural evidence of tinnitus. J. Physiol. 2014, 592, 5065–5078. [Google Scholar] [CrossRef] [PubMed]
- Basura, G.J.; Koehler, S.D.; Shore, S.E. Bimodal stimulus timing-dependent plasticity in primary auditory cortex is altered after noise exposure with and without tinnitus. J. Neurophysiol. 2015, 114, 3064–3075. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Martel, D.T.; Shore, S.E. Increased Synchrony and Bursting of Dorsal Cochlear Nucleus Fusiform Cells Correlate with Tinnitus. J. Neurosci. 2016, 36, 2068–2073. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.A.; Barry, K.M.; Zimdahl, J.W.; Leggett, K.; Mulders, W. Spontaneous firing patterns in the medial geniculate nucleus in a guinea pig model of tinnitus. Hear. Res. 2021, 403, 108190. [Google Scholar] [CrossRef]
- Lan, L.; Li, J.; Chen, Y.; Chen, W.; Li, W.; Zhao, F.; Chen, G.; Liu, J.; Chen, Y.; Li, Y.; et al. Alterations of brain activity and functional connectivity in transition from acute to chronic tinnitus. Hum. Brain Mapp. 2021, 42, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Kok, T.E.; Domingo, D.; Hassan, J.; Vuong, A.; Hordacre, B.; Clark, C.; Katrakazas, P.; Shekhawat, G.S. Resting-state Networks in Tinnitus: A Scoping Review. Clin. Neuroradiol. 2022, 1–20. [Google Scholar] [CrossRef]
- Rauschecker, J.P.; Leaver, A.M.; Muhlau, M. Tuning out the noise: Limbic-auditory interactions in tinnitus. Neuron 2010, 66, 819–826. [Google Scholar] [CrossRef]
- Rauschecker, J.P.; May, E.S.; Maudoux, A.; Ploner, M. Frontostriatal Gating of Tinnitus and Chronic Pain. Trends Cogn. Sci. 2015, 19, 567–578. [Google Scholar] [CrossRef]
- Lan, L.; Chen, Y.C.; Shang, S.; Lu, L.; Xu, J.J.; Yin, X.; Wu, Y.; Cai, Y. Topological features of limbic dysfunction in chronicity of tinnitus with intact hearing: New hypothesis for ‘noise-cancellation’ mechanism. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 113, 110459. [Google Scholar] [CrossRef]
- Lefaucheur, J.P.; Andre-Obadia, N.; Antal, A.; Ayache, S.S.; Baeken, C.; Benninger, D.H.; Cantello, R.M.; Cincotta, M.; de Carvalho, M.; De Ridder, D.; et al. Evidence-based guidelines on the therapeutic use of repetitive transcranial magnetic stimulation (rTMS). Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2014, 125, 2150–2206. [Google Scholar] [CrossRef]
- Seewoo, B.J.; Etherington, S.J.; Rodger, J. Transcranial Magnetic Stimulation. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2019; pp. 1–8. [Google Scholar]
- Eldaief, M.C.; Press, D.Z.; Pascual-Leone, A. Transcranial magnetic stimulation in neurology: A review of established and prospective applications. Neurol. Clin. Pract. 2013, 3, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Somaa, F.A.; de Graaf, T.A.; Sack, A.T. Transcranial Magnetic Stimulation in the Treatment of Neurological Diseases. Front. Neurol. 2022, 13, 793253. [Google Scholar] [CrossRef] [PubMed]
- Hallett, M. Transcranial magnetic stimulation and the human brain. Nature 2000, 406, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Kleinjung, T.; Eichhammer, P.; Landgrebe, M.; Sand, P.; Hajak, G.; Steffens, T.; Strutz, J.; Langguth, B. Combined temporal and prefrontal transcranial magnetic stimulation for tinnitus treatment: A pilot study. Otolaryngol. Head Neck Surg. 2008, 138, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Ciminelli, P.; Machado, S.; Palmeira, M.; Coutinho, E.S.F.; Sender, D.; Nardi, A.E. Dorsomedial Prefrontal Cortex Repetitive Transcranial Magnetic Stimulation for Tinnitus: Promising Results of a Blinded, Randomized, Sham-Controlled Study. Ear Hear. 2021, 42, 12–19. [Google Scholar] [CrossRef]
- Langguth, B.; Landgrebe, M.; Frank, E.; Schecklmann, M.; Sand, P.G.; Vielsmeier, V.; Hajak, G.; Kleinjung, T. Efficacy of different protocols of transcranial magnetic stimulation for the treatment of tinnitus: Pooled analysis of two randomized controlled studies. World J. Biol. Psychiatry Off. J. World Fed. Soc. Biol. Psychiatry 2014, 15, 276–285. [Google Scholar] [CrossRef]
- Denton, A.J.; Finberg, A.; Ashman, P.E.; Bencie, N.B.; Scaglione, T.; Kuzbyt, B.; Telischi, F.F.; Mittal, R.; Eshraghi, A.A. Implications of Transcranial Magnetic Stimulation as a Treatment Modality for Tinnitus. J. Clin. Med. 2021, 10, 5422. [Google Scholar] [CrossRef]
- Watson, N.; Schaper, F.; Jabbour, S.; Sadler, S.; Bain, P.A.; Fox, M.D.; Naples, J.G. Is There an Optimal Repetitive Transcranial Magnetic Stimulation Target to Treat Chronic Tinnitus? Otolaryngol. Head Neck Surg. 2022. [Google Scholar] [CrossRef]
- Wilson, S.; Croarkin, P.E.; Aaronson, S.T.; Carpenter, L.L.; Cochran, M.; Stultz, D.J.; Kozel, F.A. Systematic review of preservation TMS that includes continuation, maintenance, relapse-prevention, and rescue TMS. J. Affect. Disord. 2022, 296, 79–88. [Google Scholar] [CrossRef]
- Muller, M.; Kamping, S.; Benrath, J.; Skowronek, H.; Schmitz, J.; Klinger, R.; Flor, H. Treatment history and placebo responses to experimental and clinical pain in chronic pain patients. Eur. J. Pain 2016, 20, 1530–1541. [Google Scholar] [CrossRef]
- Duckert, L.G.; Rees, T.S. Placebo effect in tinnitus management. Otolaryngol. Head Neck Surg. 1984, 92, 697–699. [Google Scholar] [CrossRef] [PubMed]
- Zimdahl, J.W.; Thomas, H.; Bolland, S.J.; Leggett, K.; Barry, K.M.; Rodger, J.; Mulders, W. Excitatory Repetitive Transcranial Magnetic Stimulation Over Prefrontal Cortex in a Guinea Pig Model Ameliorates Tinnitus. Front. Neurosci. 2021, 15, 693935. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.; Bester, C.; Vogler, D.; Mulders, W.H. Spontaneous hyperactivity in the auditory midbrain: Relationship to afferent input. Hear. Res. 2013, 295, 124–129. [Google Scholar] [CrossRef]
- Mulders, W.H.; Barry, K.M.; Robertson, D. Effects of furosemide on cochlear neural activity, central hyperactivity and behavioural tinnitus after cochlear trauma in guinea pig. PLoS ONE 2014, 9, e97948. [Google Scholar] [CrossRef]
- Mulders, W.H.; McMahen, C.; Robertson, D. Effects of chronic furosemide on central neural hyperactivity and cochlear thresholds after cochlear trauma in Guinea pig. Front. Neurol. 2014, 5, 146. [Google Scholar] [CrossRef] [PubMed]
- Leggett, K.; Mendis, V.; Mulders, W. Divergent Responses in the Gap Prepulse Inhibition of the Acoustic Startle Reflex in Two Different Guinea Pig Colonies. Int. Tinnitus J. 2018, 22, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mulders, W.H.; Ding, D.; Salvi, R.; Robertson, D. Relationship between auditory thresholds, central spontaneous activity, and hair cell loss after acoustic trauma. J. Comp. Neurol. 2011, 519, 2637–2647. [Google Scholar] [CrossRef]
- Mulders, W.H.; Robertson, D. Progressive centralization of midbrain hyperactivity after acoustic trauma. Neuroscience 2011, 192, 753–760. [Google Scholar] [CrossRef]
- Vogler, D.P.; Robertson, D.; Mulders, W.H. Hyperactivity in the ventral cochlear nucleus after cochlear trauma. J. Neurosci. 2011, 31, 6639–6645. [Google Scholar] [CrossRef]
- Parthoens, J.; Verhaeghe, J.; Servaes, S.; Miranda, A.; Stroobants, S.; Staelens, S. Performance Characterization of an Actively Cooled Repetitive Transcranial Magnetic Stimulation Coil for the Rat. Neuromodul. J. Int. Neuromodul. Soc. 2016, 19, 459–468. [Google Scholar] [CrossRef]
- Mulders, W.; Leggett, K.; Mendis, V.; Tarawneh, H.; Wong, J.K.; Rodger, J. Low-intensity repetitive transcranial magnetic stimulation over prefrontal cortex in an animal model alters activity in the auditory thalamus but does not affect behavioural measures of tinnitus. Exp. Brain Res. 2019, 237, 883–896. [Google Scholar] [CrossRef] [PubMed]
- Mulders, W.H.; Vooys, V.; Makowiecki, K.; Tang, A.D.; Rodger, J. The effects of repetitive transcranial magnetic stimulation in an animal model of tinnitus. Sci. Rep. 2016, 6, 38234. [Google Scholar] [CrossRef] [PubMed]
- Eggermont, J.J.; Roberts, L.E. Tinnitus: Animal models and findings in humans. Cell Tissue Res. 2015, 361, 311–336. [Google Scholar] [CrossRef] [PubMed]
- Cody, A.R.; Johnstone, B.M. Acoustic trauma: Single neuron basis for the “half-octave shift”. J. Acoust. Soc. Am. 1981, 70, 707–711. [Google Scholar] [CrossRef]
- Gao, W.Y.; Ding, D.L.; Zheng, X.Y.; Ruan, F.M.; Liu, Y.J. A comparison of changes in the stereocilia between temporary and permanent hearing losses in acoustic trauma. Hear. Res. 1992, 62, 27–41. [Google Scholar] [CrossRef]
- Mulders, W.H.; Robertson, D. Development of hyperactivity after acoustic trauma in the guinea pig inferior colliculus. Hear. Res. 2013, 298, 104–108. [Google Scholar] [CrossRef]
- Cody, A.R.; Johnstone, B.M. Reduced temporary and permanent hearing losses with multiple tone exposures. Hear. Res. 1982, 6, 291–301. [Google Scholar] [CrossRef]
- Attias, J.; Pratt, H. Auditory evoked potentials and audiological follow-up of subjects developing noise-induced permanent threshold shift. Audiol. Off. Organ Int. Soc. Audiol. 1984, 23, 498–508. [Google Scholar] [CrossRef]
- Melnick, W. Human temporary threshold shift (TTS) and damage risk. J. Acoust. Soc. Am. 1991, 90, 147–154. [Google Scholar] [CrossRef]
- Hertzano, R.; Lipford, E.L.; Depireux, D. Noise: Acoustic Trauma to the Inner Ear. Otolaryngol. Clin. N. Am. 2020, 53, 531–542. [Google Scholar] [CrossRef]
- Ryan, A.F.; Kujawa, S.G.; Hammill, T.; Le Prell, C.; Kil, J. Temporary and Permanent Noise-induced Threshold Shifts: A Review of Basic and Clinical Observations. Otol. Neurotol. 2016, 37, e271–e275. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.A.; Mackey, C.A.; MacDonald, K.S.; Hackett, T.A.; Ramachandran, R. Changes in audiometric threshold and frequency selectivity correlate with cochlear histopathology in macaque monkeys with permanent noise-induced hearing loss. Hear. Res. 2020, 398, 108082. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.Z.; O’Malley, J.T.; de Gruttola, V.; Liberman, M.C. Primary Neural Degeneration in Noise-Exposed Human Cochleas: Correlations with Outer Hair Cell Loss and Word-Discrimination Scores. J. Neurosci. 2021, 41, 4439–4447. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Larsen, D.; Hughes, L.; Moechars, D.; Shore, S. Time course of tinnitus development following noise exposure in mice. J. Neurosci. Res. 2012, 90, 1480–1488. [Google Scholar] [CrossRef]
- Longenecker, R.J.; Galazyuk, A.V. Variable Effects of Acoustic Trauma on Behavioral and Neural Correlates of Tinnitus In Individual Animals. Front. Behav. Neurosci. 2016, 10, 207. [Google Scholar] [CrossRef]
- Middleton, J.W.; Kiritani, T.; Pedersen, C.; Turner, J.G.; Shepherd, G.M.; Tzounopoulos, T. Mice with behavioral evidence of tinnitus exhibit dorsal cochlear nucleus hyperactivity because of decreased GABAergic inhibition. Proc. Natl. Acad. Sci. USA 2011, 108, 7601–7606. [Google Scholar] [CrossRef]
- Ruttiger, L.; Singer, W.; Panford-Walsh, R.; Matsumoto, M.; Lee, S.C.; Zuccotti, A.; Zimmermann, U.; Jaumann, M.; Rohbock, K.; Xiong, H.; et al. The reduced cochlear output and the failure to adapt the central auditory response causes tinnitus in noise exposed rats. PLoS ONE 2013, 8, e57247. [Google Scholar] [CrossRef]
- Yenigun, A.; Dogan, R.; Aksoy, F.; Akyuz, S.; Dabak, H. Assessment of tinnitus with tinnitus severity index, tinnitus handicap inventory and distortion product otoacoustic emissions in patients with normal hearing and hearing loss. Turk. J. Ear Nose Throat 2014, 24, 11–16. [Google Scholar] [CrossRef][Green Version]
- Muhlmeier, G.; Baguley, D.; Cox, T.; Suckfull, M.; Meyer, T. Characteristics and Spontaneous Recovery of Tinnitus Related to Idiopathic Sudden Sensorineural Hearing Loss. Otol. Neurotol. 2016, 37, 634–641. [Google Scholar] [CrossRef]
- Penner, M.J. Variability in matches to subjective tinnitus. J. Speech Hear. Res. 1983, 26, 263–267. [Google Scholar] [CrossRef]
- Man, A.; Naggan, L. Characteristics of tinnitus in acoustic trauma. Audiol. Off. Organ Int. Soc. Audiol. 1981, 20, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Zhang, X.; Huang, Z.; Feng, X. The Characteristic and Short-Term Prognosis of Tinnitus Associated with Sudden Sensorineural Hearing Loss. Neural Plast. 2018, 2018, 6059697. [Google Scholar] [CrossRef] [PubMed]
- Mondelli, M.F.C.G.; Rocha, A.B. Between the Audiologic Findings and Tinnitus Disorder. Int. Arch. Otorhinolaryngol. 2011, 15, 172–180. [Google Scholar]
- Lehner, A.; Schecklmann, M.; Greenlee, M.W.; Rupprecht, R.; Langguth, B. Triple-site rTMS for the treatment of chronic tinnitus: A randomized controlled trial. Sci. Rep. 2016, 6, 22302. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer, P.M.; Poeppl, T.B.; Vielsmeier, V.; Schecklmann, M.; Langguth, B.; Lehner, A. The more the merrier? Preliminary results regarding treatment duration and stimulation frequency of multisite repetitive transcranial magnetic stimulation in chronic tinnitus. Prog. Brain Res. 2021, 262, 287–307. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer, P.M.; Landgrebe, M.; Schecklmann, M.; Poeppl, T.B.; Vielsmeier, V.; Hajak, G.; Kleinjung, T.; Langguth, B. Can Temporal Repetitive Transcranial Magnetic Stimulation be Enhanced by Targeting Affective Components of Tinnitus with Frontal rTMS? A Randomized Controlled Pilot Trial. Front. Syst. Neurosci. 2011, 5, 88. [Google Scholar] [CrossRef]
- Noh, T.S.; Kyong, J.S.; Chang, M.Y.; Park, M.K.; Lee, J.H.; Oh, S.H.; Kim, J.S.; Chung, C.K.; Suh, M.W. Comparison of Treatment Outcomes Following Either Prefrontal Cortical-only or Dual-site Repetitive Transcranial Magnetic Stimulation in Chronic Tinnitus Patients: A Double-blind Randomized Study. Otol. Neurotol. 2017, 38, 296–303. [Google Scholar] [CrossRef]
- Poeppl, T.B.; Langguth, B.; Lehner, A.; Frodl, T.; Rupprecht, R.; Kreuzer, P.M.; Landgrebe, M.; Schecklmann, M. Brain stimulation-induced neuroplasticity underlying therapeutic response in phantom sounds. Hum. Brain Mapp. 2018, 39, 554–562. [Google Scholar] [CrossRef]
- Yang, H.; Cheng, G.; Liang, Z.; Deng, W.; Huang, X.; Gao, M.; Zheng, Y. Efficacy of Repetitive Transcranial Magnetic Stimulation (rTMS) for Tinnitus: A Retrospective Study. Ear Nose Throat J. 2021, 01455613211016896. [Google Scholar] [CrossRef]
- Hoogendam, J.M.; Ramakers, G.M.; Di Lazzaro, V. Physiology of repetitive transcranial magnetic stimulation of the human brain. Brain Stimul. 2010, 3, 95–118. [Google Scholar] [CrossRef]
- Klomjai, W.; Katz, R.; Lackmy-Vallee, A. Basic principles of transcranial magnetic stimulation (TMS) and repetitive TMS (rTMS). Ann. Phys. Rehabil. Med. 2015, 58, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Cornwall, J.; Cooper, J.D.; Phillipson, O.T. Projections to the rostral reticular thalamic nucleus in the rat. Exp. Brain Res. 1990, 80, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Zikopoulos, B.; Barbas, H. Prefrontal projections to the thalamic reticular nucleus form a unique circuit for attentional mechanisms. J. Neurosci. 2006, 26, 7348–7361. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.J.; Xu, X.X.; He, S.; He, J. Change detection by thalamic reticular neurons. Nat. Neurosci. 2009, 12, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Pandya, D.N.; Kuypers, H.G. Cortico-cortical connections in the rhesus monkey. Brain Res. 1969, 13, 13–36. [Google Scholar] [CrossRef]
- Petrides, M.; Pandya, D.N. Association fiber pathways to the frontal cortex from the superior temporal region in the rhesus monkey. J. Comp. Neurol. 1988, 273, 52–66. [Google Scholar] [CrossRef]
- Winkowski, D.E.; Nagode, D.A.; Donaldson, K.J.; Yin, P.; Shamma, S.A.; Fritz, J.B.; Kanold, P.O. Orbitofrontal Cortex Neurons Respond to Sound and Activate Primary Auditory Cortex Neurons. Cereb. Cortex 2018, 28, 868–879. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amat, F.; Zimdahl, J.W.; Barry, K.M.; Rodger, J.; Mulders, W.H.A.M. Long-Term Effects of Repetitive Transcranial Magnetic Stimulation on Tinnitus in a Guinea Pig Model. Brain Sci. 2022, 12, 1096. https://doi.org/10.3390/brainsci12081096
Amat F, Zimdahl JW, Barry KM, Rodger J, Mulders WHAM. Long-Term Effects of Repetitive Transcranial Magnetic Stimulation on Tinnitus in a Guinea Pig Model. Brain Sciences. 2022; 12(8):1096. https://doi.org/10.3390/brainsci12081096
Chicago/Turabian StyleAmat, Farah, Jack W. Zimdahl, Kristin M. Barry, Jennifer Rodger, and Wilhelmina H. A. M. Mulders. 2022. "Long-Term Effects of Repetitive Transcranial Magnetic Stimulation on Tinnitus in a Guinea Pig Model" Brain Sciences 12, no. 8: 1096. https://doi.org/10.3390/brainsci12081096
APA StyleAmat, F., Zimdahl, J. W., Barry, K. M., Rodger, J., & Mulders, W. H. A. M. (2022). Long-Term Effects of Repetitive Transcranial Magnetic Stimulation on Tinnitus in a Guinea Pig Model. Brain Sciences, 12(8), 1096. https://doi.org/10.3390/brainsci12081096