
Resting-State Functional Connectivity Analyses: Brain Functional Reorganization in a Rat Model of Postherpetic Neuralgia

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Nociceptive Behavioral Tests
2.3. fMRI Examination
2.4. Data Processing and Functional Connectivity Analysis
2.5. Statistical Analysis
3. Results
3.1. Nociceptive Behavioral Tests
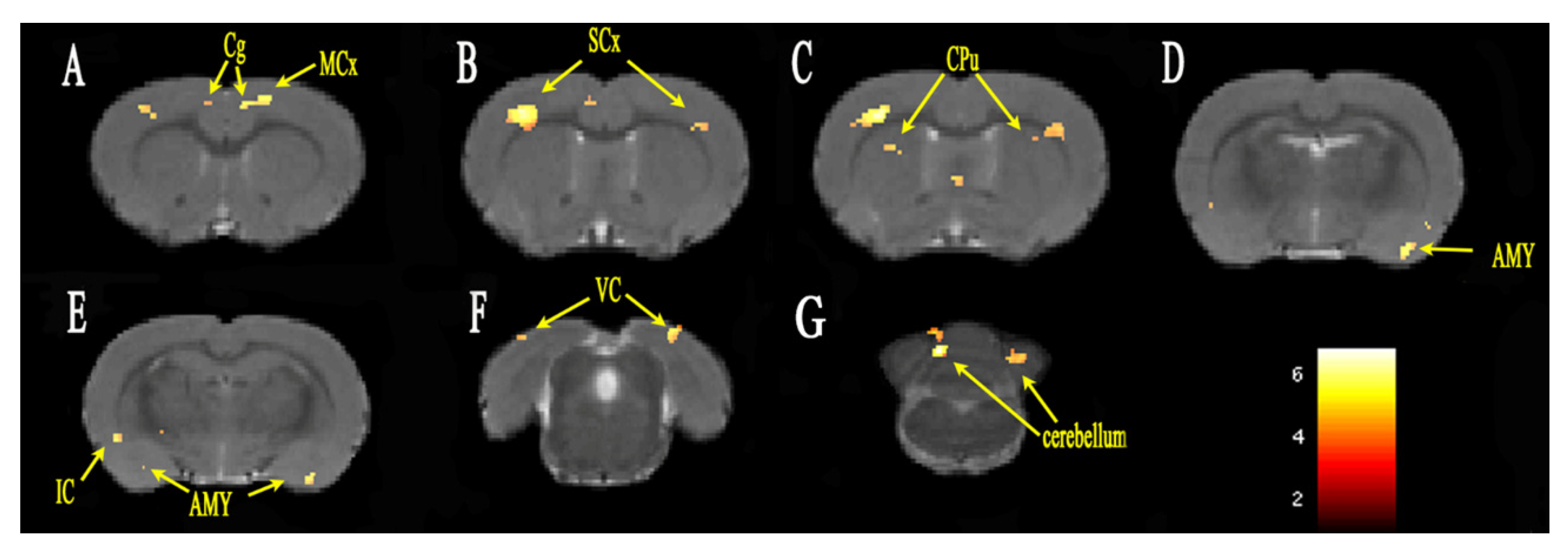
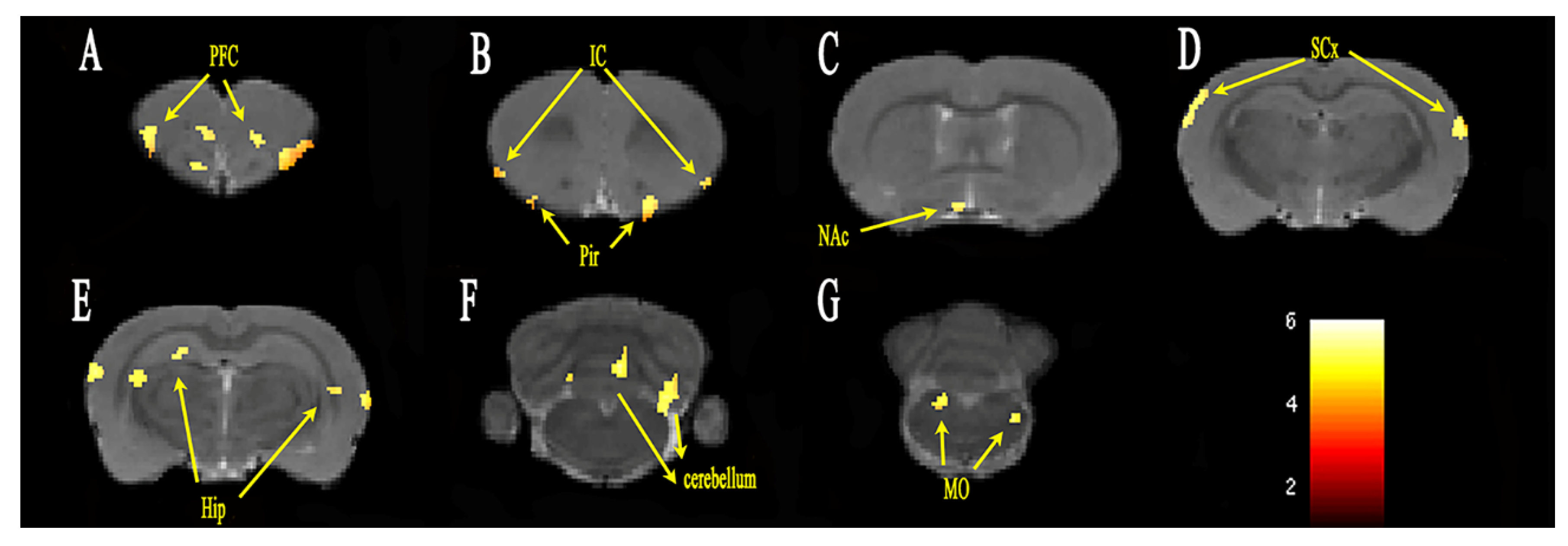
3.2. Functional Connectivity Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PHN | postherpetic neuralgia |
| RTX | resiniferatoxin |
| fMRI | functional magnetic resonance imaging |
| Day 7 | the seventh day after modeling |
| Day 14 | the 14th day after modeling |
| FC | functional connection |
| ACC | anterior cingulate |
| HZ | herpes zoster |
| PFC | prefrontal cortices |
| Thal | thalamus |
| CON | control |
| RAGE | turbo-rapid acquisition relaxation enhancement |
| ROI | region of interest |
| NAc | nucleus accumbens |
| Mcx | motor cortex |
| Scx | sensory cortex |
| Cpu | Caudate Putamen |
| IC | insular cortex |
| AMY | amygdala |
| VC | visual cortex |
| Pir | piriform cortex |
| Hip | hippocampus |
| MO | medulla oblongata |
References
- Thompson, R.R.; Kong, C.L.; Porco, T.C.; Kim, E.; Ebert, C.D.; Acharya, N.R. Herpes Zoster and Postherpetic Neuralgia: Changing Incidence Rates From 1994 to 2018 in the United States. Clin. Infect. Dis. 2021, 73, e3210–e3217. [Google Scholar] [CrossRef] [PubMed]
- Varghese, L.; Standaert, B.; Olivieri, A.; Curran, D. The temporal impact of aging on the burden of herpes zoster. BMC Geriatr. 2017, 17, 30. [Google Scholar] [CrossRef] [Green Version]
- Curran, D.; Schmidt-Ott, R.; Schutter, U.; Simon, J.; Anastassopoulou, A.; Matthews, S. Impact of herpes zoster and postherpetic neuralgia on the quality of life of Germans aged 50 or above. BMC Infect. Dis. 2018, 18, 496. [Google Scholar] [CrossRef] [PubMed]
- Matthews, S.; De Maria, A.; Passamonti, M.; Ristori, G.; Loiacono, I.; Puggina, A.; Curran, D. The Economic Burden and Impact on Quality of Life of Herpes Zoster and Postherpetic Neuralgia in Individuals Aged 50 Years or Older in Italy. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2019; Volume 6. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, M.; Zheng, T.; Xiao, Y.; Wang, S.; Han, F.; Chen, G. Structural and functional brain abnormalities in postherpetic neuralgia: A systematic review of neuroimaging studies. Brain Res. 2021, 1752, 147219. [Google Scholar] [CrossRef] [PubMed]
- Melzack, R.; Wall, P.D.; Steptoe, A.; Wardle, J. Pain mechanisms: A new theory. Psychosoc. Process. Health Read. 1965, 150, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Larrea, L.; Bastuji, H. Pain and consciousness. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 87, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.L.; Khan, G.M.; Alloway, K.D.; Chen, S.R. Resiniferatoxin induces paradoxical changes in thermal and mechanical sensitivities in rats: Mechanism of action. J. Neurosci. 2003, 23, 2911–2919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Wang, L.; Hua, J.; Jin, X.H.; Ji, F.; Peng, K.; Zhou, B.; Yang, J.; Meng, X.W. Inhibiting BDNF/TrkB.T1 receptor improves resiniferatoxin-induced postherpetic neuralgia through decreasing ASIC3 signaling in dorsal root ganglia. J. Neuroinflamm. 2021, 18, 96. [Google Scholar] [CrossRef] [PubMed]
- Dalziel, R.G.; Bingham, S.; Sutton, D.; Grant, D.; Champion, J.M.; Dennis, S.A.; Quinn, J.P.; Bountra, C.; Mark, M.A. Allodynia in rats infected with varicella zoster virus--a small animal model for post-herpetic neuralgia. Brain Res. Rev. 2004, 46, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Apkarian, A.V.; Hashmi, J.A.; Baliki, M.N. Pain and the brain: Specificity and plasticity of the brain in clinical chronic pain. Pain 2011, 152 (Suppl. 3), S49–S64. [Google Scholar] [CrossRef] [PubMed]
- Baliki, M.N.; Chang, P.C.; Baria, A.T.; Centeno, M.V.; Apkarian, A.V. Resting-sate functional reorganization of the rat limbic system following neuropathic injury. Sci. Rep. 2014, 4, 6186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbard, C.S.; Khan, S.A.; Xu, S.; Cha, M.; Masri, R.; Seminowicz, D.A. Behavioral, metabolic and functional brain changes in a rat model of chronic neuropathic pain: A longitudinal MRI study. NeuroImage 2015, 107, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Bilbao, A.; Falfán-Melgoza, C.; Leixner, S.; Becker, R.; Singaravelu, S.K.; Sack, M.; Sartorius, A.; Spanagel, R.; Weber-Fahr, W. Longitudinal Structural and Functional Brain Network Alterations in a Mouse Model of Neuropathic Pain. Neuroscience 2018, 387, 104–115. [Google Scholar] [CrossRef]
- Muthulingam, J.; Olesen, S.S.; Hansen, T.M.; Seminowicz, D.A.; Burrowes, S.; Drewes, A.M.; Frøkjær, J.B. Progression of Structural Brain Changes in Patients with Chronic Pancreatitis and Its Association to Chronic Pain: A 7-Year Longitudinal Follow-up Study. Pancreas 2018, 47, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Lv, Z.T.; Zhao, Y.; Gao, Y.; Li, J.Q.; Gao, F.; Meng, X.F.; Tian, B.; Shi, J.; Pan, H.L.; et al. Electroacupuncture improves thermal and mechanical sensitivities in a rat model of postherpetic neuralgia. Mol. Pain 2013, 9, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.L.; Kuner, R. Neocortical circuits in pain and pain relief. Nat. Rev. Neurosci. 2021, 22, 458–471. [Google Scholar] [CrossRef]
- Khan, G.M.; Chen, S.R.; Pan, H.L. Role of primary afferent nerves in allodynia caused by diabetic neuropathy in rats. Neuroscience 2002, 114, 291–299. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Lehto, L.J.; Idiyatullin, D.; Zhang, J.; Utecht, L.; Adriany, G.; Garwood, M.; Gröhn, O.; Michaeli, S.; Mangia, S. MB-SWIFT functional MRI during deep brain stimulation in rats. Neuroimage 2017, 159, 443–448. [Google Scholar] [CrossRef]
- Hennig, J.; Nauerth, A.; Friedburg, H. RARE imaging: A fast imaging method for clinical MR. Magn. Reson. Med. 1986, 3, 823–833. [Google Scholar] [CrossRef]
- Chen, G.; Yang, B.; Chen, J.; Zhu, L.; Jiang, H.; Yu, W.; Zang, F.; Chen, Y.; Dai, Y. Changes in Male Rat Sexual Behavior and Brain Activity Revealed by Functional Magnetic Resonance Imaging in Response to Chronic Mild Stress. J. Sex. Med. 2018, 15, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Khazipov, R.; Zaynutdinova, D.; Ogievetsky, E.; Valeeva, G.; Mitrukhina, O.; Manent, J.B.; Represa, A. Atlas of the Postnatal Rat Brain in Stereotaxic Coordinates. Front. Neuroanat. 2015, 9, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowbotham, M.C.; Fields, H.L. The relationship of pain, allodynia and thermal sensation in post-herpetic neuralgia. Brain A J. Neurol. 1996, 119 Pt 2, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vierck, C.J.; Whitsel, B.L.; Favorov, O.V.; Brown, A.W.; Tommerdahl, M. Role of primary somatosensory cortex in the coding of pain. Pain 2013, 154, 334–344. [Google Scholar] [CrossRef] [Green Version]
- Taoka, M.I.K.I.; Toda, T.; Hihara, S.; Tanaka, M.; Iriki, A.; Iwamura, Y. A systematic analysis of neurons with large somatosensory receptive fields covering multiple body regions in the secondary somatosensory area of macaque monkeys. J. Neurophysiol. 2016, 116, 2152–2162. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Larrea, L.; Peyron, R. Pain matrices and neuropathic pain matrices: A review. Pain 2013, 154 (Suppl. 1), S29–S43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geha, P.Y.; Baliki, M.N.; Wang, X.; Harden, R.N.; Paice, J.A.; Apkarian, A.V. Brain dynamics for perception of tactile allodynia (touch-induced pain) in postherpetic neuralgia. Pain 2008, 138, 641–656. [Google Scholar] [CrossRef] [Green Version]
- Barcelo, A.C.; Filippini, B.; Pazo, J.H. The striatum and pain modulation. Cell Mol. Neurobiol. 2012, 32, 1–12. [Google Scholar] [CrossRef]
- Makin, T.R.; Flor, H. Brain (re)organisation following amputation: Implications for phantom limb pain. NeuroImage 2020, 218, 116943. [Google Scholar] [CrossRef]
- Swart, C.M.; Stins, J.F.; Beek, P.J. Cortical changes in complex regional pain syndrome (CRPS). Eur. J. Pain 2009, 13, 902–907. [Google Scholar] [CrossRef]
- Tsao, H.; Galea, M.P.; Hodges, P.W. Reorganization of the motor cortex is associated with postural control deficits in recurrent low back pain. Brain 2008, 131, 2161–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moulton, E.A.; Schmahmann, J.D.; Becerra, L.; Borsook, D. The cerebellum and pain: Passive integrator or active participator? Brain Res. Rev. 2010, 65, 14–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmchen, C.; Mohr, C.; Erdmann, C.; Petersen, D.; Nitschke, M.F. Differential cerebellar activation related to perceived pain intensity during noxious thermal stimulation in humans: A functional magnetic resonance imaging study. Neurosci. Lett. 2003, 335, 202–206. [Google Scholar] [CrossRef]
- Neugebauer, V.; Li, W.; Bird, G.C.; Han, J.S. The amygdala and persistent pain. Neuroscientist 2004, 10, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, J.; Chen, L.; Zhang, M.; Wan, Y. Basolateral amygdala lesion inhibits the development of pain chronicity in neuropathic pain rats. PLoS ONE 2013, 8, e70921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkus, C.; McHugh, S.B.; Sprengel, R.; Seeburg, P.H.; Rawlins, J.N.P.; Bannerman, D.M. Hippocampal NMDA receptors and anxiety: At the interface between cognition and emotion. Eur. J. Pharmacol. 2010, 626, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutso, A.A.; Radzicki, D.; Baliki, M.N.; Huang, L.; Banisadr, G.; Centeno, M.V.; Radulovic, J.; Martina, M.; Miller, R.J.; Apkarian, A.V. Abnormalities in hippocampal functioning with persistent pain. J. Neurosci. 2012, 32, 5747–5756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, W.J.; Liu, Y.; Zhou, L.J.; Li, W.; Zhong, Y.; Pang, R.P.; Xin, W.J.; Wei, X.H.; Wang, J.; Zhu, H.Q.; et al. Peripheral nerve injury leads to working memory deficits and dysfunction of the hippocampus by upregulation of TNF-alpha in rodents. Neuropsychopharmacology 2011, 36, 979–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apkarian, A.V. Pain perception in relation to emotional learning. Curr. Opin. Neurobiol. 2008, 18, 464–468. [Google Scholar] [CrossRef] [Green Version]
- Gusnard, D.A.; Akbudak, E.; Shulman, G.L.; Raichle, M.E. Medial prefrontal cortex and self-referential mental activity: Relation to a default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 4259–4264. [Google Scholar] [CrossRef] [Green Version]
- Metz, A.E.; Yau, H.J.; Centeno, M.V.; Apkarian, A.V.; Martina, M. Morphological and functional reorganization of rat medial prefrontal cortex in neuropathic pain. Proc. Natl. Acad. Sci. USA 2009, 106, 2423–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.C.; Pollema-Mays, S.L.; Centeno, M.V.; Procissi, D.; Contini, M.; Baria, A.T.; Martina, M.; Apkarian, A.V. Role of nucleus accumbens in neuropathic pain: Linked multi-scale evidence in the rat transitioning to neuropathic pain. Pain 2014, 155, 1128–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baliki, M.N.; Petre, B.; Torbey, S.; Herrmann, K.M.; Huang, L.; Schnitzer, T.J.; Fields, H.L.; Apkarian, A.V. Corticostriatal functional connectivity predicts transition to chronic back pain. Nat. Neurosci. 2012, 15, 1117–1119. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Li, Y.; Deng, W.; Qin, B.; Zhang, Y.; Xie, P.; Yuan, J.; Yu, B.; Tian, Y. Local Brain Activity Differences Between Herpes Zoster and Postherpetic Neuralgia Patients: A Resting-State Functional MRI Study. Pain Physician 2017, 20, E687–E699. [Google Scholar]
- Liu, J.; Hao, Y.; Du, M.; Wang, X.; Zhang, J.; Manor, B.; Jiang, X.; Fang, W.; Wang, D. Quantitative cerebral blood flow mapping and functional connectivity of postherpetic neuralgia pain: A perfusion fMRI study. Pain 2013, 154, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, M.C.; Ceko, M.; Low, L.A. Cognitive and emotional control of pain and its disruption in chronic pain. Nat. Rev. Neurosci. 2013, 14, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Ventura-Antunes, L.; Mota, B.; Herculano-Houzel, S. Different scaling of white matter volume, cortical connectivity, and gyrification across rodent and primate brains. Front. Neuroanat. 2013, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Halley, A.C.; Krubitzer, L. Not all cortical expansions are the same: The coevolution of the neocortex and the dorsal thalamus in mammals. Curr. Opin. Neurobiol. 2019, 56, 78–86. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coordinates of Peak Voxel | |||||
|---|---|---|---|---|---|
| Brain Region | Cluster Size | t-Value | X | Y | Z |
| L cingulate gyrus | 87 | 5.037 | 0.4984 | 1.4022 | 1.8021 |
| R cingulate gyrus | 93 | 4.029 | −0.5641 | 1.3830 | 1.3221 |
| L motor cortex | 128 | 5.120 | 1.4339 | 1.1642 | 1.8021 |
| R motor cortex | 31 | 4.065 | −3.6445 | 1.1627 | 1.8021 |
| L sensory cortex | 333 | 5.021 | 4.6644 | 3.8351 | 0.1221 |
| R sensory cortex | 964 | 6.204 | −3.8953 | 1.8379 | 0.6021 |
| L amygdaloid body | 176 | 5.107 | 4.1497 | 8.8969 | −1.3179 |
| R amygdaloid body | 86 | 4.628 | −5.7135 | 9.3718 | −3.2379 |
| L striatum | 97 | 3.972 | 2.6994 | 7.3334 | −2.7579 |
| R striatum | 174 | 4.562 | −3.0901 | 3.6578 | 0.3621 |
| L poaterior lobe of cerebellum | 251 | 5.621 | 3.2290 | 1.9082 | −12.1179 |
| R poaterior lobe of cerebellum | 217 | 6.825 | −1.1515 | 3.3053 | −14.2279 |
| L corpus callosum | 39 | 4.485 | 3.4518 | 2.8256 | 0.8421 |
| R corpus callosum | 52 | 4.287 | −4.8110 | 3.2258 | −0.8379 |
| L visual cortex | 547 | 4.854 | 3.8444 | 1.1680 | −8.2779 |
| R visual cortex | 298 | 5.059 | −4.8357 | 1.2926 | −8.7579 |
| L capsule | 41 | 4.527 | 2.7060 | 7.6681 | −3.2379 |
| R capsule | 81 | 4.329 | −3.7187 | 6.2112 | −2.5179 |
| R insular cortex | 59 | 4.444 | −5.8636 | 6.3370 | −2.0379 |
| L anterior lobe of cerebellum | 25 | 5.569 | 3.0954 | 1.9005 | −12.1179 |
| R bed nucleus of stria terminalis | 23 | 4.122 | −1.0756 | 5.8819 | −0.3579 |
| L cerebellar nucleus | 94 | 4.542 | 2.3166 | 4.1936 | −13.7979 |
| L claustral layer | 24 | 4.725 | 4.9416 | 7.5651 | −0.5979 |
| R olfactory cortex | 99 | 4.971 | −5.7646 | 2.4493 | −9.2379 |
| R pfi flocculonodular lobe | 20 | 5.495 | −1.2951 | 3.1515 | −14.2779 |
| L piriform cortex | 183 | 4.693 | 4.9449 | 7.5864 | −0.8379 |
| R septal area | 86 | 4.543 | −0.6812 | 5.2784 | 0.1221 |
| L temporal association cortex | 20 | 3.961 | 6.1131 | 2.8843 | −8.0379 |
| L cerebral peduncle | 27 | 4.506 | 2.7093 | 7.6894 | −3.4779 |
| L subthalamic nucleus | 20 | 4.497 | 2.7093 | 7.5434 | −3.4779 |
| L tegmentum of pons | 60 | 4.750 | 1.221 | 8.0601 | −11.8779 |
| Coordinates of Peak Voxel | |||||
|---|---|---|---|---|---|
| Brain Region | Cluster Size | t-Value | X | Y | Z |
| L sensory cortex | 234 | 4.400 | 7.0997 | 4.0198 | −2.0379 |
| R sensory cortex | 365 | 4.786 | −6.7826 | 1.8627 | −3.2379 |
| L insular cortex | 317 | 4.574 | 4.8987 | 6.2660 | 2.5221 |
| R insular cortex | 63 | 3.896 | −5.532 | 4.7447 | 3.0021 |
| L hippocampus | 34 | 5.225 | 5.6824 | 8.3644 | −5.8779 |
| R hippocampus | 428 | 4.892 | −5.4264 | 4.1121 | −4.6779 |
| L orbital cortex | 473 | 4.702 | 0.57926 | 2.6754 | 5.6421 |
| R orbital cortex | 353 | 4.400 | −3.9580 | 2.8934 | 5.1621 |
| L frontal association cortex | 64 | 4.603 | 0.5859 | 2.5082 | 5.8821 |
| R frontal association cortex | 92 | 4.431 | −3.5604 | 2.6033 | 5.4021 |
| L piriform cortex | 292 | 5.483 | 3.1680 | 8.1065 | 2.4021 |
| R piriform cortex | 199 | 4.852 | −1.8131 | 4.5199 | 4.6821 |
| L anterior lobe of cerebellum | 208 | 5.486 | 0.6898 | 4.6820 | −12.1179 |
| R anterior lobe of cerebellum | 84 | 4.831 | −1.4749 | 0.8833 | −10.1979 |
| L cerebellar nucleus | 64 | 5.131 | 3.0954 | 6.1352 | −12.1179 |
| R cerebellar nucleus | 30 | 4.306 | −2.1101 | 4.8544 | −12.5979 |
| L corpus callosum | 52 | 4.086 | 5.5554 | 8.3993 | −6.3579 |
| R corpus callosum | 31 | 4.381 | −5.8273 | 4.2350 | −4.6779 |
| L dentate gyrus | 22 | 4.479 | 3.1465 | 8.0929 | −6.1179 |
| R dentate gyrus | 93 | 4.221 | −2.4962 | 2.0278 | −3.9579 |
| L medulla oblongata | 163 | 5.314 | 2.3166 | 7.8442 | −13.7979 |
| R medulla oblongata | 257 | 5.089 | −2.5011 | 6.6474 | −13.3179 |
| L poaterior lobe of cerebellum | 306 | 5.700 | 2.8050 | 1.7360 | −10.4379 |
| R poaterior lobe of cerebellum | 301 | 5.638 | −2.808 | 1.5575 | −10.4379 |
| L pfi flocculonodular lobe | 46 | 5.168 | 3.0954 | 6.2813 | −12.1179 |
| R pfi flocculonodular lobe | 47 | 4.281 | −2.1134 | 4.9791 | −12.3579 |
| L tegmentum of pons | 63 | 5.097 | 2.3133 | 7.6769 | −13.5579 |
| R tegmentum of pons | 69 | 5.040 | −2.3675 | 6.6551 | −13.3179 |
| L auditory cortex | 359 | 4.379 | 6.8489 | 4.1109 | −3.2379 |
| R auditory cortex | 256 | 4.909 | −7.1736 | 2.9256 | −3.9579 |
| L olfactory bulb | 327 | 4.653 | 0.5759 | 2.6541 | 5.8821 |
| R olfactory bulb | 246 | 4.707 | −1.9467 | 4.5122 | 4.6821 |
| L olfactory tubercle | 207 | 5.126 | 3.1713 | 8.2739 | 1.8021 |
| R olfactory tubercle | 52 | 3.979 | −0.5707 | 7.4734 | 1.8021 |
| R nucleus accumbens | 67 | 4.236 | −1.7702 | 7.4252 | 1.5621 |
| L anterior commissure | 51 | 4.198 | 1.6880 | 6.4974 | 2.7621 |
| R frontal cortex area 3 | 24 | 4.025 | −3.8177 | 2.9438 | 4.6821 |
| R motor cortex | 21 | 4.080 | −3.5571 | 2.4785 | 5.1621 |
| R preoptic region | 69 | 4.048 | −0.9485 | 7.7454 | 0.1221 |
| R septal area | 116 | 4.512 | −5.0354 | 3.4873 | −3.9579 |
| R striatum | 46 | 4.026 | −1.2224 | 7.6874 | 0.6021 |
| L tenia tecta | 33 | 3.823 | 1.5412 | 5.6744 | 3.7221 |
| R third ventricle | 73 | 4.430 | −5.2994 | 3.6391 | −4.1979 |
| L fourth ventricle | 42 | 4.777 | 2.8281 | 6.7039 | −12.1179 |
| L inferior colliculus | 26 | 4.029 | 3.0492 | 3.6467 | −8.7579 |
| L olfactory cortex | 105 | 5.359 | 5.8161 | 8.5181 | −5.8779 |
| L visual cortex | 24 | 4.629 | 2.3876 | 1.6063 | −9.2379 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, S.; Wu, G.; Wei, X.; Meng, X.; Zang, F.; Shen, L.; Dai, H.; Wang, L.; Li, Y. Resting-State Functional Connectivity Analyses: Brain Functional Reorganization in a Rat Model of Postherpetic Neuralgia. Brain Sci. 2022, 12, 1029. https://doi.org/10.3390/brainsci12081029
Han S, Wu G, Wei X, Meng X, Zang F, Shen L, Dai H, Wang L, Li Y. Resting-State Functional Connectivity Analyses: Brain Functional Reorganization in a Rat Model of Postherpetic Neuralgia. Brain Sciences. 2022; 12(8):1029. https://doi.org/10.3390/brainsci12081029
Chicago/Turabian StyleHan, Shuting, Guanzuan Wu, Xiang Wei, Xiaowen Meng, Fengchao Zang, Lan Shen, Hui Dai, Lina Wang, and Yonggang Li. 2022. "Resting-State Functional Connectivity Analyses: Brain Functional Reorganization in a Rat Model of Postherpetic Neuralgia" Brain Sciences 12, no. 8: 1029. https://doi.org/10.3390/brainsci12081029
APA StyleHan, S., Wu, G., Wei, X., Meng, X., Zang, F., Shen, L., Dai, H., Wang, L., & Li, Y. (2022). Resting-State Functional Connectivity Analyses: Brain Functional Reorganization in a Rat Model of Postherpetic Neuralgia. Brain Sciences, 12(8), 1029. https://doi.org/10.3390/brainsci12081029