
Nimodipine Exerts Beneficial Effects on the Rat Oligodendrocyte Cell Line OLN-93

 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture
2.2. Nimodipine
2.3. Reverse Transcription Polymerase Chain Reactions
2.4. Immunocytochemistry
2.5. Patch-Clamp Analysis
2.6. Real-Time Quantitative Polymerase Chain Reaction
2.7. Transmission Electron Microscopy
2.8. Whole Transcriptome Analysis/RNA Sequencing
2.9. Statistical Analysis
3. Results
3.1. OLN-93 Cells Resemble Oligodendrocytes and do Not Express Cav1.2 and Cav1.3
3.2. Nimodipine Increases the Expression of Plp1 in OLN-93 Cells
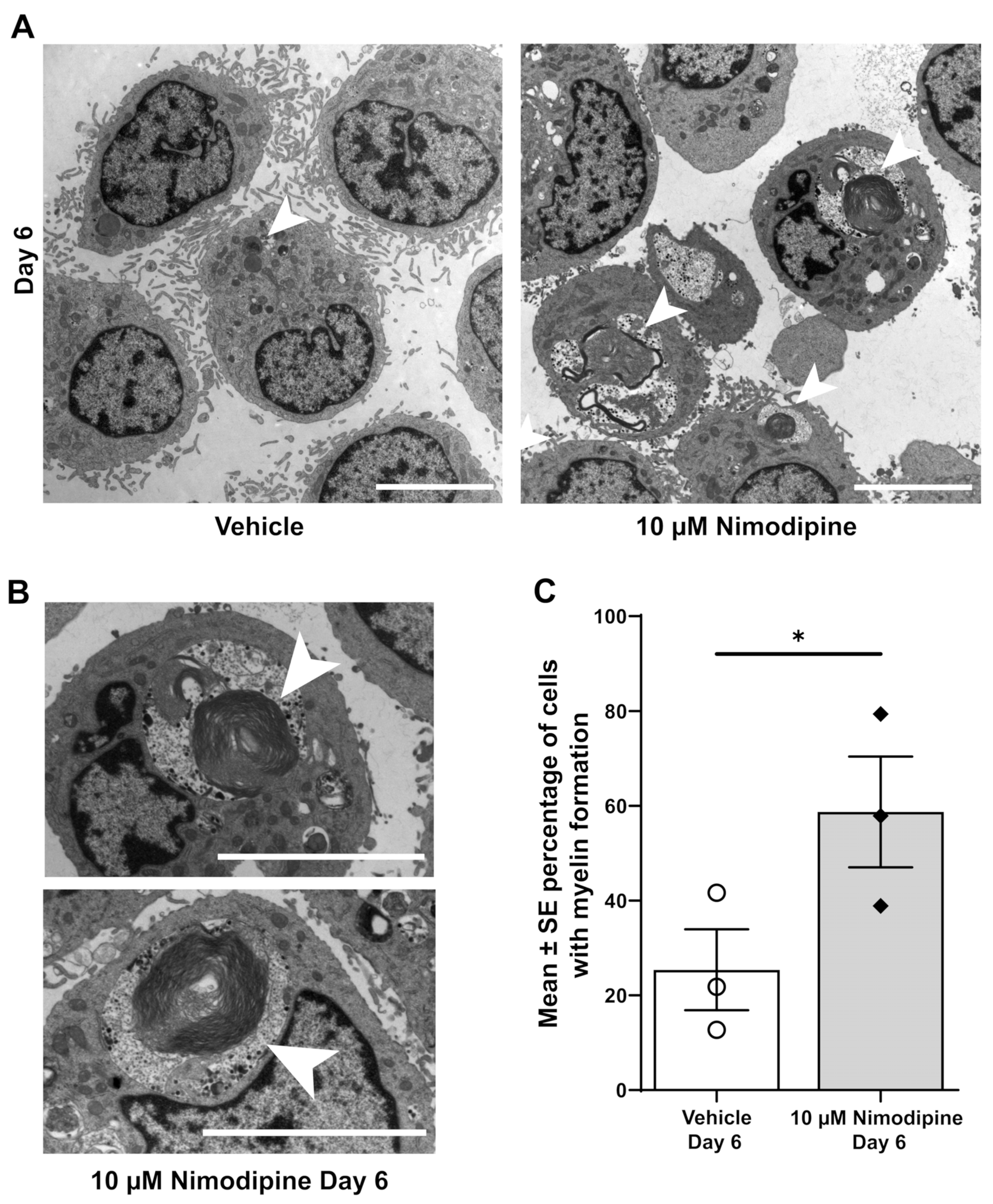
3.3. Nimodipine Increases the Formation of Myelin at the Ultrastructural Level
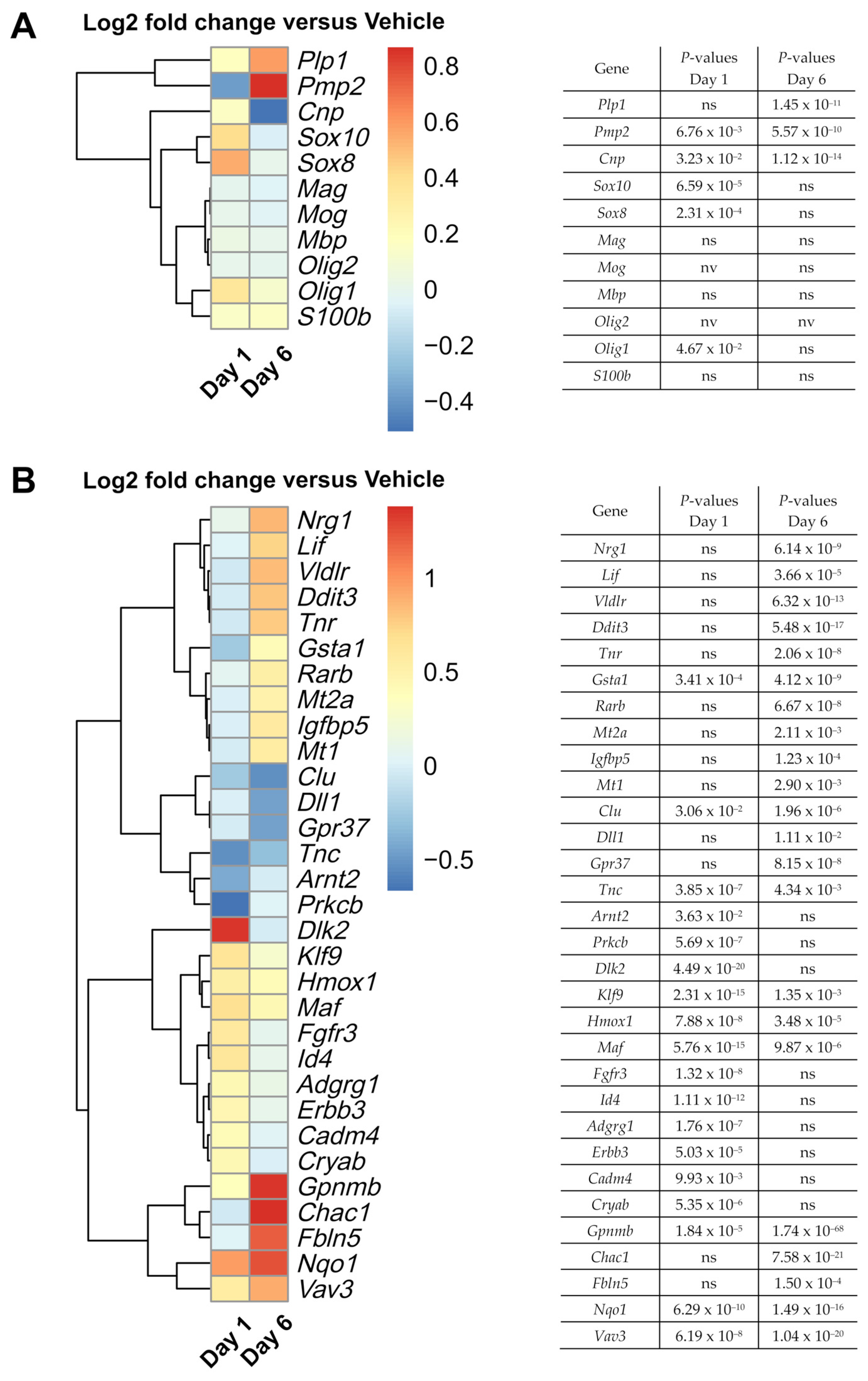
3.4. Nimodipine Shifts RNA Expression towards Myelination and Maturation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wallin, M.T.; Culpepper, W.J.; Nichols, E.; Bhutta, Z.A.; Gebrehiwot, T.T.; Hay, S.I.; Khalil, I.A.; Krohn, K.J.; Liang, X.; Naghavi, M.; et al. Global, regional, and national burden of multiple sclerosis 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 269–285. [Google Scholar] [CrossRef]
- Kobelt, G.; Thompson, A.; Berg, J.; Gannedahl, M.; Eriksson, J. New insights into the burden and costs of multiple sclerosis in Europe. Mult. Scler. 2017, 23, 1123–1136. [Google Scholar] [CrossRef] [PubMed]
- Mandia, D.; Ferraro, O.E.; Nosari, G.; Montomoli, C.; Zardini, E.; Bergamaschi, R. Environmental factors and multiple sclerosis severity: A descriptive study. Int. J. Environ. Res. Public Health 2014, 11, 6417–6432. [Google Scholar] [CrossRef]
- Hafler, D.A.; Compston, A.; Sawcer, S.; Lander, E.S.; Daly, M.J.; De Jager, P.L.; de Bakker, P.I.; Gabriel, S.B.; Mirel, D.B.; Ivinson, A.J.; et al. Risk alleles for multiple sclerosis identified by a genomewide study. N. Engl. J. Med. 2007, 357, 851–862. [Google Scholar] [PubMed]
- Sintzel, M.B.; Rametta, M.; Reder, A.T. Vitamin D and multiple sclerosis: A comprehensive review. Neurol. Ther. 2018, 7, 59–85. [Google Scholar] [CrossRef]
- Marrodan, M.; Alessandro, L.; Farez, M.F.; Correale, J. The role of infections in multiple sclerosis. Mult. Scler. 2019, 25, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Trapp, B.D.; Nave, K.-A. Multiple sclerosis: An immune or neurodegenerative disorder? Annu. Rev. Neurosci. 2008, 31, 247–269. [Google Scholar] [CrossRef] [PubMed]
- Kuerten, S.; Lanz, T.V.; Lingampalli, N.; Lahey, L.J.; Kleinschnitz, C.; Mäurer, M.; Schroeter, M.; Braune, S.; Ziemssen, T.; Ho, P.P.; et al. Autoantibodies against central nervous system antigens in a subset of B cell-dominant multiple sclerosis patients. Proc. Natl. Acad. Sci. USA 2020, 117, 21512–21518. [Google Scholar] [CrossRef] [PubMed]
- Comi, G.; Radaelli, M.; Soelberg Sørensen, P. Evolving concepts in the treatment of relapsing multiple sclerosis. Lancet 2017, 389, 1347–1356. [Google Scholar] [CrossRef]
- Lassmann, H. Multiple sclerosis pathology. Cold Spring Harb. Perspect. Med. 2018, 8, a028936. [Google Scholar] [CrossRef] [PubMed]
- Ellwardt, E.; Zipp, F. Molecular mechanisms linking neuroinflammation and neurodegeneration in MS. Exp. Neurol. 2014, 262 Pt A, 8–17. [Google Scholar] [CrossRef]
- Villoslada, P.; Steinman, L. New targets and therapeutics for neuroprotection, remyelination and repair in multiple sclerosis. Expert Opin. Investig. Drugs 2020, 29, 443–459. [Google Scholar] [CrossRef] [PubMed]
- Cree, B.A.C.; Cutter, G.; Wolinsky, J.S.; Freedman, M.S.; Comi, G.; Giovannoni, G.; Hartung, H.-P.; Arnold, D.; Kuhle, J.; Block, V.; et al. Safety and efficacy of MD1003 (high-dose biotin) in patients with progressive multiple sclerosis (SPI2): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Neurol. 2020, 19, 988–997. [Google Scholar] [CrossRef]
- Mi, S.; Miller, R.H.; Lee, X.; Scott, M.L.; Shulag-Morskaya, S.; Shao, Z.; Chang, J.; Thill, G.; Levesque, M.; Zhang, M.; et al. LINGO-1 negatively regulates myelination by oligodendrocytes. Nat. Neurosci. 2005, 8, 745–751. [Google Scholar] [CrossRef]
- Cadavid, D.; Mellion, M.; Hupperts, R.; Edwards, K.R.; Calabresi, P.A.; Drulović, J.; Giovannoni, G.; Hartung, H.P.; Arnold, D.L.; Fisher, E.; et al. Safety and efficacy of opicinumab in patients with relapsing multiple sclerosis (SYNERGY): A randomised, placebo-controlled, phase 2 trial. Lancet Neurol. 2019, 18, 845–856. [Google Scholar] [CrossRef]
- Ahmed, Z.; Fulton, D.; Douglas, M.R. Opicinumab: Is it a potential treatment for multiple sclerosis? Ann. Transl. Med. 2020, 8, 892. [Google Scholar] [CrossRef] [PubMed]
- Carlson, A.P.; Hänggi, D.; Macdonald, R.L.; Shuttleworth, C.W. Nimodipine reappraised: An old drug with a future. Curr. Neuropharmacol. 2020, 18, 65–82. [Google Scholar] [CrossRef]
- Scriabine, A.; van den Kerckhoff, W. Pharmacology of nimodipine. A review. Ann. N. Y. Acad. Sci. 1988, 522, 698–706. [Google Scholar] [CrossRef]
- Daou, B.J.; Koduri, S.; Thompson, B.G.; Chaudhary, N.; Pandey, A.S. Clinical and experimental aspects of aneurysmal subarachnoid hemorrhage. CNS Neurosci. Ther. 2019, 25, 1096–1112. [Google Scholar] [CrossRef]
- Keyrouz, S.G.; Diringer, M.N. Clinical review: Prevention and therapy of vasospasm in subarachnoid hemorrhage. Crit. Care 2007, 11, 220. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Y.; Bao, Y.; Yin, C.; Xin, X.; Guo, Y.; Gao, F.; Huo, S.; Wang, X.; Wang, Q. Pretreatment with nimodipine reduces incidence of POCD by decreasing calcineurin mediated hippocampal neuroapoptosis in aged rats. BMC Anesthesiol. 2018, 18, 42. [Google Scholar] [CrossRef] [PubMed]
- Taya, K.; Watanabe, Y.; Kobayashi, H.; Fujiwara, M. Nimodipine improves the disruption of spatial cognition induced by cerebral ischemia. Physiol. Behav. 2000, 70, 19–25. [Google Scholar] [CrossRef]
- Schampel, A.; Volovitch, O.; Koeniger, T.; Scholz, C.J.; Jörg, S.; Linker, R.A.; Wischmeyer, E.; Wunsch, M.; Hell, J.W.; Ergün, S.; et al. Nimodipine fosters remyelination in a mouse model of multiple sclerosis and induces microglia-specific apoptosis. Proc. Natl. Acad. Sci. USA 2017, 114, E3295–E3304. [Google Scholar] [CrossRef] [PubMed]
- Ingwersen, J.; De Santi, L.; Wingerath, B.; Graf, J.; Koop, B.; Schneider, R.; Hecker, C.; Schröter, F.; Bayer, M.; Engelke, A.D.; et al. Nimodipine confers clinical improvement in two models of experimental autoimmune encephalomyelitis. J. Neurochem. 2018, 146, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Zamora, N.N.; Cheli, V.T.; Santiago González, D.A.; Wan, R.; Paez, P.M. Deletion of voltage-gated calcium channels in astrocytes during demyelination reduces brain inflammation and promotes myelin regeneration in mice. J. Neurosci. 2020, 40, 3332–3347. [Google Scholar] [CrossRef]
- Lin, R.J.; Klein-Fedyshin, M.; Rosen, C.A. Nimodipine improves vocal fold and facial motion recovery after injury: A systematic review and meta-analysis. Laryngoscope 2019, 129, 943–951. [Google Scholar] [CrossRef]
- Scheller, C.; Rampp, S.; Leisz, S.; Tatagiba, M.; Gharabaghi, A.; Ramina, K.F.; Ganslandt, O.; Matthies, C.; Westermaier, T.; Antoniadis, G.; et al. Prophylactic nimodipine treatment improves hearing outcome after vestibular schwannoma surgery in men: A subgroup analysis of a randomized multicenter phase III trial. Neurosurg. Rev. 2020, 44, 1729–1735. [Google Scholar] [CrossRef]
- Leisz, S.; Simmermacher, S.; Prell, J.; Strauss, C.; Scheller, C. Nimodipine-dependent protection of Schwann cells, astrocytes and neuronal cells from osmotic, oxidative and heat stress is associated with the activation of AKT and CREB. Int. J. Mol. Sci. 2019, 20, 4578. [Google Scholar] [CrossRef]
- Orefice, N.S.; Guillemot-Legris, O.; Capasso, R.; Bottemanne, P.; Hantraye, P.; Caraglia, M.; Orefice, G.; Alhouayek, M.; Muccioli, G.G. miRNA profile is altered in a modified EAE mouse model of multiple sclerosis featuring cortical lesions. eLife 2020, 9, e56916. [Google Scholar] [CrossRef]
- Kim, H.N.; Langley, M.R.; Simon, W.L.; Yoon, H.; Kleppe, L.; Lanza, I.R.; LeBrasseur, N.K.; Matveyenko, A.; Scarisbrick, I.A. A Western diet impairs CNS energy homeostasis and recovery after spinal cord injury: Link to astrocyte metabolism. Neurobiol. Dis. 2020, 141, 104934. [Google Scholar] [CrossRef]
- Bargagna-Mohan, P.; Schultz, G.; Rheaume, B.; Trakhtenberg, E.F.; Robson, P.; Pal-Ghosh, S.; Stepp, M.A.; Given, K.S.; Macklin, W.B.; Mohan, R. Corneal nonmyelinating Schwann cells illuminated by single-cell transcriptomics and visualized by protein biomarkers. J. Neurosci. Res. 2021, 99, 731–749. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, O.; Aliuos, P.; Wissel, K.; Lenarz, T.; Werner, D.; Reuter, G.; Kral, A.; Warnecke, A. Dissociated Neurons and Glial Cells Derived from Rat Inferior Colliculi after Digestion with Papain. PLoS ONE 2013, 8, e80490. [Google Scholar] [CrossRef] [PubMed]
- Keen, A.N.; Payne, L.A.; Mehta, V.; Rice, A.; Simpson, L.J.; Pang, K.L.; del Rio Hernandez, A.; Reader, J.S.; Tzima, E. Eukaryotic initiation factor 6 regulates mechanical responses in endothelial cells. J. Cell Biol. 2022, 221, e202005213. [Google Scholar] [CrossRef] [PubMed]
- McCloy, R.A.; Rogers, S.; Caldon, C.E.; Lorca, T.; Castro, A.; Burgess, A. Partial inhibition of Cdk1 in G 2 phase overrides the SAC and decouples mitotic events. Cell Cycle 2014, 13, 1400–1412. [Google Scholar] [CrossRef]
- Babai, N.; Gierke, K.; Müller, T.; Regus-Leidig, H.; Brandstätter, J.H.; Feigenspan, A. Signal transmission at invaginating cone photoreceptor synaptic contacts following deletion of the presynaptic cytomatrix protein Bassoon in mouse retina. Acta Physiol. 2019, 226, e13241. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: http://www.R-project.org (accessed on 8 February 2021).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Richter-Landsberg, C.; Heinrich, M. OLN-93: A new permanent oligodendroglia cell line derived from primary rat brain glial cultures. J. Neurosci. Res. 1996, 45, 161–173. [Google Scholar] [CrossRef]
- Stolt, C.C.; Rehberg, S.; Ader, M.; Lommes, P.; Riethmacher, D.; Schachner, M.; Bartsch, U.; Wegner, M. Terminal differentiation of myelin-forming oligodendrocytes depends on the transcription factor Sox10. Genes. Dev. 2002, 16, 165–170. [Google Scholar] [CrossRef]
- Kuhlbrodt, K.; Herbarth, B.; Sock, E.; Hermans-Borgmeyer, I.; Wegner, M. Sox10, a novel transcriptional modulator in glial cells. J. Neurosci. 1998, 18, 237–250. [Google Scholar] [CrossRef]
- Schlick, B.; Flucher, B.E.; Obermair, G.J. Voltage-activated calcium channel expression profiles in mouse brain and cultured hippocampal neurons. Neuroscience 2010, 167, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Enders, M.; Heider, T.; Ludwig, A.; Kuerten, S. Strategies for neuroprotection in multiple sclerosis and the role of calcium. Int. J. Mol. Sci. 2020, 21, 1663. [Google Scholar] [CrossRef] [PubMed]
- Koschak, A.; Fernandez-Quintero, M.L.; Heigl, T.; Ruzza, M.; Seitter, H.; Zanetti, L. Cav1.4 dysfunction and congenital stationary night blindness type 2. Pflügers Arch.-Eur. J. Physiol. 2021, 473, 1437–1454. [Google Scholar] [CrossRef] [PubMed]
- Stadelmann, C.; Timmler, S.; Barrantes-Freer, A.; Simons, M. Myelin in the central nervous system: Structure, function, and pathology. Physiol. Rev. 2019, 99, 1381–1431. [Google Scholar] [CrossRef] [PubMed]
- Hayasaka, K.; Himoro, M.; Takada, G.; Takahashi, E.; Minoshima, S.; Shimizu, N. Structure and localization of the gene encoding human peripheral myelin protein 2 (PMP2). Genomics 1993, 18, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Raasakka, A.; Kursula, P. The myelin membrane-associated enzyme 2’,3’-cyclic nucleotide 3’-phosphodiesterase: On a highway to structure and function. Neurosci. Bull. 2014, 30, 956–966. [Google Scholar] [CrossRef]
- Dai, J.; Bercury, K.K.; Ahrendsen, J.T.; Macklin, W.B. Olig1 function is required for oligodendrocyte differentiation in the mouse brain. J. Neurosci. 2015, 35, 4386–4402. [Google Scholar] [CrossRef]
- Turnescu, T.; Arter, J.; Reiprich, S.; Tamm, E.R.; Waisman, A.; Wegner, M. Sox8 and Sox10 jointly maintain myelin gene expression in oligodendrocytes. Glia 2018, 66, 279–294. [Google Scholar] [CrossRef]
- Pesheva, P.; Gloor, S.; Schachner, M.; Probstmeier, R. Tenascin-R is an intrinsic autocrine factor for oligodendrocyte differentiation and promotes cell adhesion by a sulfatide-mediated mechanism. J. Neurosci. 1997, 17, 4642–4651. [Google Scholar] [CrossRef]
- Czopka, T.; Von Holst, A.; Schmidt, G.; Ffrench-Constant, C.; Faissner, A. Tenascin C and tenascin R similarly prevent the formation of myelin membranes in a RhoA-dependent manner, but antagonistically regulate the expression of myelin basic protein via a separate pathway. Glia 2009, 57, 1790–1801. [Google Scholar] [CrossRef]
- Rittchen, S.; Boyd, A.; Burns, A.; Park, J.; Fahmy, T.M.; Metcalfe, S.; Williams, A. Myelin repair in vivo is increased by targeting oligodendrocyte precursor cells with nanoparticles encapsulating leukaemia inhibitory factor (LIF). Biomaterials 2015, 56, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-J.; Vainshtein, A.; Maik-Rachline, G.; Peles, E. G protein-coupled receptor 37 is a negative regulator of oligodendrocyte differentiation and myelination. Nat. Commun. 2016, 7, 10884. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, T.; Lee, P.R.; Baba, H.; Fields, R.D. Leukemia inhibitory factor regulates the timing of oligodendrocyte development and myelination in the postnatal optic nerve. J. Neurosci. Res. 2009, 87, 3343–3355. [Google Scholar] [CrossRef] [PubMed]
- Dugas, J.C.; Ibrahim, A.; Barres, B.A. The T3-induced gene KLF9 regulates oligodendrocyte differentiation and myelin regeneration. Mol. Cell. Neurosci. 2012, 50, 45–57. [Google Scholar] [CrossRef]
- Chora, Â.A.; Fontoura, P.; Cunha, A.; Pais, T.F.; Cardoso, S.; Ho, P.P.; Lee, L.Y.; Sobel, R.A.; Steinman, L.; Soares, M.P. Heme oxygenase–1 and carbon monoxide suppress autoimmune neuroinflammation. J. Clin. Investig. 2007, 117, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Ulc, A.; Zeug, A.; Bauch, J.; van Leeuwen, S.; Kuhlmann, T.; Ffrench-Constant, C.; Ponimaskin, E.; Faissner, A. The guanine nucleotide exchange factor Vav3 modulates oligodendrocyte precursor differentiation and supports remyelination in white matter lesions. Glia 2019, 67, 376–392. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.-M.F.; Nakanishi, S.T.; Hoghooghi, V.; Eaton, S.E.A.; Palmer, A.L.; Frederick, A.; Stratton, J.A.; Stykel, M.G.; Whelan, P.J.; Zochodne, D.W.; et al. AlphaB-crystallin regulates remyelination after peripheral nerve injury. Proc. Natl. Acad. Sci. USA 2017, 114, E1707–E1716. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Hu, X.; Park, J.; Zhu, Y.; Zhu, Q.; Li, H.; Luo, C.; Han, R.; Cooper, N.; Qiu, M. Selective expression of LDLR and VLDLR in myelinating oligodendrocytes. Dev. Dyn. 2007, 236, 2708–2712. [Google Scholar] [CrossRef]
- Kim, M.; Wende, H.; Walcher, J.; Kühnemund, J.; Cheret, C.; Kempa, S.; McShane, E.; Selbach, M.; Lewin, G.R.; Birchmeier, C. Maf links Neuregulin1 signaling to cholesterol synthesis in myelinating Schwann cells. Genes Dev. 2018, 32, 645–657. [Google Scholar] [CrossRef]
- Won, S.Y.; Kwon, S.; Jeong, H.S.; Chung, K.W.; Choi, B.O.; Chang, J.W.; Lee, J.E. Fibulin 5, a human Wharton’s jelly-derived mesenchymal stem cells-secreted paracrine factor, attenuates peripheral nervous system myelination defects through the Integrin-RAC1 signaling axis. Stem Cells 2020, 38, 1578–1593. [Google Scholar] [CrossRef]
- Stringer, J.L.; Gaikwad, A.; Gonzales, B.N.; Long Jr., D.J.; Marks, L.M.; Jaiswal, A.K. Presence and induction of the enzyme NAD(P)H:quinone oxidoreductase 1 in the central nervous system. J. Comp. Neurol. 2004, 471, 289–297. [Google Scholar] [CrossRef]
- Allocati, N.; Masulli, M.; Di Ilio, C.; Federici, L. Glutathione transferases: Substrates, inihibitors and pro-drugs in cancer and neurodegenerative diseases. Oncogenesis 2018, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Suzuki, Y.; Takagi, T.; Kitashoji, A.; Ono, Y.; Tsuruma, K.; Yoshimura, S.; Shimazawa, M.; Iwama, T.; Hara, H. Glycoprotein nonmetastatic melanoma protein B (GPNMB) as a novel neuroprotective factor in cerebral ischemia-reperfusion injury. Neuroscience 2014, 277, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, Y.; Shimazawa, M.; Tanaka, H.; Ono, Y.; Noda, Y.; Ohuchi, K.; Tsuruma, K.; Katsuno, M.; Sobue, G.; Hara, H. Glycoprotein nonmetastatic melanoma protein B ameliorates skeletal muscle lesions in a SOD1G93A mouse model of amyotrophic lateral sclerosis. J. Neurosci. Res. 2015, 93, 1552–1566. [Google Scholar] [CrossRef] [PubMed]
- Ousman, S.S.; Tomooka, B.H.; van Noort, J.M.; Wawrousek, E.F.; O’Conner, K.; Hafler, D.A.; Sobel, R.A.; Robinson, W.H.; Steinman, L. Protective and therapeutic role for αB-crystallin in autoimmune demyelination. Nature 2007, 448, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Monin, A.; Baumann, P.S.; Griffa, A.; Xin, L.; Mekle, R.; Fournier, M.; Butticaz, C.; Klaey, M.; Cabungcal, J.H.; Steullet, P.; et al. Glutathione deficit impairs myelin maturation: Relevance for white matter integrity in schizophrenia patients. Mol. Psychiatry 2015, 20, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Gow, A.; Wrabetz, L. CHOP and the endoplasmic reticulum stress response in myelinating glia. Curr. Opin. Neurobiol. 2009, 19, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Fischbach, F.; Nedelcu, J.; Leopold, P.; Zhan, J.; Clarner, T.; Nellessen, L.; Beißel, C.; van Heuvel, Y.; Goswami, A.; Weis, J.; et al. Cuprizone-induced graded oligodendrocyte vulnerability is regulated by the transcription factor DNA damage-inducible transcript 3. Glia 2019, 67, 263–276. [Google Scholar] [CrossRef]
- Southwood, C.M.; Fykkolodziej, B.; Maheras, K.J.; Garshott, D.M.; Estill, M.; Fribley, A.M.; Gow, A. Overexpression of CHOP in myelinating cells does not confer a significant phenotype under normal or metabolic stress conditions. J. Neurosci. 2016, 36, 6803–6819. [Google Scholar] [CrossRef]
- Southwood, C.M.; Garbern, J.; Jiang, W.; Gow, A. The unfolded protein response modulates disease severity in Pelizaeus-Merzbacher disease. Neuron 2002, 36, 585–596. [Google Scholar] [CrossRef]
- Latasa, M.J.; Ituero, M.; Moran-Gonzalez, A.; Aranda, A.; Cosgaya, J.M. Retinoic acid regulates myelin formation in the peripheral nervous system. Glia 2010, 58, 1451–1464. [Google Scholar] [CrossRef] [PubMed]
- Morrison, V.E.; Smith, V.N.; Huang, J.K. Retinoic acid is required for oligodendrocyte precursor cell production and differentiation in the postnatal mouse corpus callosum. eNeuro 2020, 7, ENEURO.0270-19.2019. [Google Scholar] [CrossRef] [PubMed]
- Siddiq, M.M.; Hannila, S.S.; Carmel, J.B.; Bryson, J.B.; Hou, J.; Nikulina, E.; Willis, M.R.; Mellado, W.; Richman, E.L.; Hilaire, M.; et al. Metallothionein-I/II promotes axonal regeneration in the central nervous system. J. Biol. Chem. 2015, 290, 16343–16356. [Google Scholar] [CrossRef]
- Penkowa, M.; Espejo, C.; Ortega-Aznar, A.; Hidalgo, J.; Montalban, X.; Martínez Cáceres, E.M. Metallothionein expression in the central nervous system of multiple sclerosis patients. Cell. Mol. Life Sci. 2003, 60, 1258–1266. [Google Scholar] [CrossRef]
- Comes, G.; Fernandez-Gayol, O.; Molinero, A.; Giralt, M.; Capdevila, M.; Atrian, S.; Hidalgo, J. Mouse metallothionein-1 and metallothionein-2 are not biologically interchangeable in an animal model of multiple sclerosis, EAE. Metallomics 2019, 11, 327–337. [Google Scholar] [CrossRef]
- Beilharz, E.J.; Klempt, N.D.; Klempt, M.; Sirimanne, E.; Dragunow, M.; Gluckman, P.D. Differential expression of insulin-like growth factor binding proteins (IGFBP) 4 and 5 mRNA in the rat brain after transient hypoxic-ischemic injury. Brain Res. Mol. Brain Res. 1993, 18, 209–215. [Google Scholar] [CrossRef]
- van Luijn, M.M.; van Meurs, M.; Stoop, M.P.; Verbraak, E.; Wierenga-Wolf, A.F.; Melief, M.-J.; Kreft, K.L.; Verdijk, R.M.; ’t Hart, B.A.; Luider, T.M.; et al. Elevated expression of the cerebrospinal fluid disease markers chromogranin A and clusterin in astrocytes of multiple sclerosis white matter lesions. J. Neuropathol. Exp. Neurol. 2016, 75, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Polihronis, M.; Paizis, K.; Carter, G.; Sedal, L.; Murphy, B. Elevation of human cerebrospinal fluid clusterin concentration is associated with acute neuropathology. J. Neurol. Sci. 1993, 115, 230–233. [Google Scholar] [CrossRef]
- Li, H.; Ma, J.; Fang, Q.; Li, H.; Shen, H.; Li, X.; Xue, Q.; Zhu, J.; Chen, G. Botch protects neurons from ischemic insult by antagonizing Notch-mediated neuroinflammation. Exp. Neurol. 2019, 321, 113028. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Solana, B.; Nueda, M.L.; Ruvira, M.D.; Ruiz-Hidalgo, M.J.; Monsalve, E.M.; Rivero, S.; García-Ramírez, J.J.; Díaz-Guerra, M.J.; Baladrón, V.; Laborda, J. The EGF-like proteins DLK1 and DLK2 function as inhibitory non-canonical ligands of NOTCH1 receptor that modulate each other’s activities. Biochim. Biophys. Acta 2011, 1813, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, R.; Celià-Terrassa, T.; Kumar, S.; Hang, X.; Wei, Y.; Choudhury, A.; Hwang, J.; Peng, J.; Nixon, B.; Grady, J.J.; et al. Notch ligand Dll1 mediates cross-talk between mammary stem cells and the macrophageal niche. Science 2018, 360, eaan4153. [Google Scholar] [CrossRef]
- Wang, S.; Sdrulla, A.D.; diSibio, G.; Bush, G.; Nofziger, D.; Hicks, C.; Weinmaster, G.; Barres, B.A. Notch receptor activation inhibits oligodendrocyte differentiation. Neuron 1998, 21, 63–75. [Google Scholar] [CrossRef]
- Jurynczyk, M.; Jurewicz, A.; Bielecki, B.; Raine, C.S.; Selmaj, K. Inhibition of Notch signaling enhances tissue repair in an animal model of multiple sclerosis. J. Neuroimmunol. 2005, 170, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Becquart, P.; Johnston, J.; Vilariño-Güell, C.; Quandt, J.A. Oligodendrocyte ARNT2 expression is altered in models of MS. Neurol. Neuroimmunol. Neuroinflamm. 2020, 7, e745. [Google Scholar] [CrossRef]
- Stariha, R.L.; Kim, S.U. Protein kinase C and mitogen-activated protein kinase signalling in oligodendrocytes. Microsc. Res. Tech. 2001, 52, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Oh, L.Y.S.; Denninger, A.; Colvin, J.S.; Vyas, A.; Tole, S.; Ornitz, D.M.; Bansal, R. Fibroblast growth factor receptor 3 signaling regulates the onset of oligodendrocyte terminal differentiation. J. Neurosci. 2003, 23, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Marin-Husstege, M.; He, Y.; Li, J.; Kondo, T.; Sablitzky, F.; Casaccia-Bonnefil, P. Multiple roles of Id4 in developmental myelination: Predicted outcomes and unexpected findings. Glia 2006, 54, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Giera, S.; Luo, R.; Ying, Y.; Ackerman, S.D.; Jeong, S.-J.; Stoveken, H.M.; Folts, C.J.; Welsh, C.A.; Tall, G.G.; Stevens, B.; et al. Microglial transglutaminase-2 drives myelination and myelin repair via GPR56/ADGRG1 in oligodendrocyte precursor cells. eLife 2018, 7, e33385. [Google Scholar] [CrossRef]
- Elazar, N.; Vainshtein, A.; Golan, N.; Vijayaragavan, B.; Schaeren-Wiemers, N.; Eshed-Eisenbach, Y.; Peles, E. Axoglial adhesion by Cadm4 regulates CNS myelination. Neuron 2019, 101, 224–231.e225. [Google Scholar] [CrossRef]
- Elazar, N.; Vainshtein, A.; Rechav, K.; Tsoory, M.; Eshed-Eisenbach, Y.; Peles, E. Coordinated internodal and paranodal adhesion controls accurate myelination by oligodendrocytes. J. Cell Biol. 2019, 218, 2887–2895. [Google Scholar] [CrossRef]
- Cheng, S.; Lv, H.; Zhang, W.; Wang, Z.; Shi, X.; Liang, W.; Yuan, Y. Adult-onset demyelinating neuropathy associated with FBLN5 gene mutation. Clin. Neuropathol. 2017, 36, 171–177. [Google Scholar] [CrossRef][Green Version]
- Alexoudi, A.; Zachaki, S.; Stavropoulou, C.; Chatzi, I.; Koumbi, D.; Stavropoulou, K.; Kollia, P.; Karageorgiou, C.E.; Sambani, C. Combined GSTP1 and NQO1 germline polymorphisms in the susceptibility to multiple sclerosis. Int. J. Neurosci. 2015, 125, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.L.; van der Pol, S.M.A.; Baron, W.; McCord, J.M.; de Vries, H.E.; van Horssen, J. Protandim Protects Oligodendrocytes against an Oxidative Insult. Antioxidants 2016, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Medrano, J.; Segura-Ulate, I.; Yang, B.; Chinnasamy, R.; Arterburn, J.B.; Perez, R.G. FTY720-Mitoxy reduces toxicity associated with MSA-like α-synuclein and oxidative stress by increasing trophic factor expression and myelin protein in OLN-93 oligodendroglia cell cultures. Neuropharmacology 2019, 158, 107701. [Google Scholar] [CrossRef] [PubMed]
- Askari, V.R.; Shafiee-Nick, R. Promising neuroprotective effects of β-caryophyllene against LPS-induced oligodendrocyte toxicity: A mechanistic study. Biochem Pharmacol 2019, 159, 154–171. [Google Scholar] [CrossRef]
- Fan, B.Y.; Pang, Y.L.; Li, W.X.; Zhao, C.X.; Zhang, Y.; Wang, X.; Ning, G.Z.; Kong, X.H.; Liu, C.; Yao, X.; et al. Liproxstatin-1 is an effective inhibitor of oligodendrocyte ferroptosis induced by inhibition of glutathione peroxidase 4. Neural Regen. Res. 2021, 16, 561–566. [Google Scholar] [CrossRef]
- Segura-Ulate, I.; Yang, B.; Vargas-Medrano, J.; Perez, R.G. FTY720 (Fingolimod) reverses α-synuclein-induced downregulation of brain-derived neurotrophic factor mRNA in OLN-93 oligodendroglial cells. Neuropharmacology 2017, 117, 149–157. [Google Scholar] [CrossRef]
- Miraee, S.; Farhadi, M.; Jameie, S.B.; Najafie, S. The protective effects of apigenin on OLN-93 cellular model of cerebral stroke. Physiol. Pharmacol. 2021, 25, 270–278. [Google Scholar] [CrossRef]
- Steiner, J.; Sarnyai, Z.; Westphal, S.; Gos, T.; Bernstein, H.-G.; Bogerts, B.; Keilhoff, G. Protective effects of haloperidol and clozapine on energy-deprived OLN-93 oligodendrocytes. Eur. Arch. Psychiatry Clin. Neurosci. 2011, 261, 477–482. [Google Scholar] [CrossRef]
- Santiago González, D.A.; Cheli, V.T.; Zamora, N.N.; Lama, T.N.; Spreuer, V.; Murphy, G.G.; Paez, P.M. Conditional deletion of the L-type calcium channel Cav1.2 in NG2-positive cells impairs remyelination in mice. J. Neurosci. 2017, 37, 10038–10051. [Google Scholar] [CrossRef]
- Cheli, V.T.; Santiago González, D.A.; Namgyal Lama, T.; Spreuer, V.; Handley, V.; Murphy, G.G.; Paez, P.M. Conditional deletion of the L-type calcium channel Cav1.2 in oligodendrocyte progenitor cells affects postnatal myelination in mice. J. Neurosci. 2016, 36, 10853–10869. [Google Scholar] [CrossRef] [PubMed]
- Cheli, V.T.; Santiago González, D.A.; Spreuer, V.; Paez, P.M. Voltage-gated Ca2+ entry promotes oligodendrocyte progenitor cell maturation and myelination in vitro. Exp. Neurol. 2015, 265, 69–83. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| Actb | 5′-AGCCTTCCTTCCTGGGTATGG-3′ | 5′-GCAGCTCAGTAACAGTCCGC-3′ |
| Cav1.2 | 5′-CGGCATCACCAACTTCGACA-3′ | 5′-GCATGCTCATGTTTCGGGGT-3′ |
| Cav1.3 | 5′-TACGTGGTGAACTCCTCGCC-3′ | 5′-CTTCGCTGAGTGCCACGTCT-3′ |
| Vehicle Day 6 | 10 µM Nimodipine Day 6 | ||
|---|---|---|---|
| Mbp | Plp1 | Mbp | Plp1 |
| 4.026 (0.6479) | 18.95 (7.631) | 4.818 (0.7941) | 33.51 (12.91) ** |
| Gene | Key Functions and Summary of Key Studies |
|---|---|
| Maf; Nrg1; Erbb3 | The upregulation of Maf supports the idea of a promyelinating effect of nimodipine, as it was shown to be involved in cholesterol synthesis in myelinating Schwann cells linking Nrg1 to cholesterol synthesis via the tyrosine kinase receptor gene Erbb3 [60]. |
| Vldlr | Vldlr was reported to be expressed in mature myelinating oligodendrocytes in the postnatal CNS of mice [59]. |
| Cryab | Cryab was reported to have anti-apoptotic, anti-inflammatory and neuroprotective effects. Cryab knockout mice developed worse experimental autoimmune encephalomyelitis (EAE), and the administration of CRYAB ameliorated clinical symptoms in EAE mice [66]. In the peripheral nervous system (PNS), Cryab is a regulator of remyelination after peripheral nerve injury [58]. |
| Gsta1 | Glutathione transferases are involved in cell detoxification and amelioration of oxidative stress [63]. Glutathione and redox regulation are important for myelination. A deficit in glutathione was reported to impair myelin formation in patients with schizophrenia [67]. |
| Lif | Lif is required for postnatal mouse optic nerve myelination and promotes oligodendrocyte survival [54]. In vitro and in vivo administration of LIF induced oligodendrocyte precursor cell (OPC) differentiation and myelination [52]. |
| Ddit 3/Chop | Controversial findings indicate a pro- and anti-apoptotic role of Ddit3/Chop [68,69,70,71]. |
| Tnc; Tnr | Tnr has regulatory properties in oligodendrocyte differentiation [50]. Tnr induces oligodendrocyte differentiation, while Tnc was reported to inhibit the process [51]. |
| Rarb | Retinoic acid is a regulator of myelination in the PNS [72]. It is required for OPC differentiation in the postnatal mouse corpus callosum [73]. |
| Mt1; Mt2a | Mt2a is the major isoform of Mt1 and Mt2 in the CNS. It promotes axonal regeneration in a variety of neurons [74]. In inactive multiple sclerosis (MS) lesions, mt1 and mt2 expression was slightly increased compared with active lesions, suggesting a role in disease remission [75]. MT2 administration ameliorated clinical symptoms in EAE mice [76]. |
| Igfbp5 | Igbp5 was reported to be involved in rat brain recovery after transient hypoxic-ischaemic injury and could play an important role in oligodendrocyte regeneration [77]. |
| Clu | CLU is elevated in the cerebrospinal fluid of patients with MS and may play a role as a neuro-inflammatory mediator [78,79]. |
| Chac1; Dlk2; Dll1 | Chac1 and Dlk2 play a role in antagonising the common signalling pathway Notch [80,81], while Dll1 is a Notch-ligand [82]. Notch1 inhibition accelerated remyelination in cuprizone-treated mice [82], whereas Notch receptor activation inhibited oligodendrocyte differentiation [83]. In EAE mice, Notch signalling inhibition ameliorated clinical symptoms and enhanced remyelination [84]. In a model of ischaemic injury, Chac1 had neuroprotective properties by antagonising Notch1 maturation [80]. |
| Gpr37 | Gpr37 is a negative regulator of myelination and oligodendrocyte differentiation [53]. |
| Arnt2 | Arnt2 expression decreased with the maturation of the OPC cell line Oli-neu. Arnt2 knockdown increased the number of myelinating oligodendrocytes [85]. |
| Prkcb | Protein kinase C (Prkc) was reported to have a proliferative effect on immature oligodendrocytes while at the same time having an inhibitory effect on oligodendrocyte differentiation. In mature oligodendrocytes, Prkc increased process extension and myelin formation [86]. |
| Hmox1 | Hmox1 knockout led to increased demyelination, paralysis, and mortality in EAE mice. Increased Hmox1 expression suppressed autoimmune neuroinflammation [56]. |
| Klf9 | Klf9 is required for normal myelin regeneration in cuprizone-treated mice [55]. |
| Fgfr3 | Regulator of oligodendrocyte development. Expression of Fgfr3 was increased when late oligodendrocyte progenitors entered terminal differentiation. Fgfr3-deficient mice showed a reduced number of differentiated oligodendrocytes and delayed myelination [87]. |
| Id4 | Overexpression of Id4 may inhibit oligodendrocyte differentiation, yet it seems to differentially regulate the expression of myelin genes as decreased levels of Plp1 were found in Id4-null mice [88]. |
| Adgrg1 (Gpr56) | Regulator of oligodendrocyte development. Loss of Gpr56 leads to hypomyelination. Transient overexpression leads to OPC proliferation. Gpr56 was relevant for remyelination in cuprizone-treated mice [89]. |
| Cadm4 | Important for axoglial adhesion and correct placement of the myelin sheath [90,91]. |
| Gpnmb | Gpnmb may play a role in neuroprotection as it led to prolonged survival in a mouse model of amyotrophic lateral sclerosis and reduced infarct volume in an ischaemic injury mouse model [64,65]. |
| Fbln5 | It plays an important role in Schwann cell development and could restore myelination in a zebrafish model of Charcot-Marie-Tooth type 1 disease (CMT1), a demyelinating disease of the PNS [61]. A subtype of autosomal CMT1 is associated with a mutation in the Fbln5 gene [92]. |
| Nqo1 | An antioxidant protein that may be an indicator of oxidative stress, but it may also be important for myelination [62]. Genetic polymorphisms of the detoxification-associated genes Nqo1 and Gstp1 (glutathione S-transferase P1) might be relevant for susceptibility and clinical presentation in patients with MS [93]. |
| Vav3 | A regulator of myelination and oligodendrocyte maturation. In the Vav3 knockout of cerebellar slice cultures, remyelination was impaired as strongly as in cuprizone-treated Vav3 knockout mice [57]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boltz, F.; Enders, M.; Feigenspan, A.; Kirchner, P.; Ekici, A.; Kuerten, S. Nimodipine Exerts Beneficial Effects on the Rat Oligodendrocyte Cell Line OLN-93. Brain Sci. 2022, 12, 476. https://doi.org/10.3390/brainsci12040476
Boltz F, Enders M, Feigenspan A, Kirchner P, Ekici A, Kuerten S. Nimodipine Exerts Beneficial Effects on the Rat Oligodendrocyte Cell Line OLN-93. Brain Sciences. 2022; 12(4):476. https://doi.org/10.3390/brainsci12040476
Chicago/Turabian StyleBoltz, Felix, Michael Enders, Andreas Feigenspan, Philipp Kirchner, Arif Ekici, and Stefanie Kuerten. 2022. "Nimodipine Exerts Beneficial Effects on the Rat Oligodendrocyte Cell Line OLN-93" Brain Sciences 12, no. 4: 476. https://doi.org/10.3390/brainsci12040476
APA StyleBoltz, F., Enders, M., Feigenspan, A., Kirchner, P., Ekici, A., & Kuerten, S. (2022). Nimodipine Exerts Beneficial Effects on the Rat Oligodendrocyte Cell Line OLN-93. Brain Sciences, 12(4), 476. https://doi.org/10.3390/brainsci12040476