
Processing of Degraded Speech in Brain Disorders

 , ,
, ,
Abstract
1. Introduction
1.1. Predictive Coding and Degraded Speech Perception
1.2. Neuroanatomy of Degraded Speech Processing
1.3. Scope of This Review
2. Factors Affecting Processing of Degraded Speech in the Healthy Brain
2.1. Healthy Ageing
2.2. Cognitive Factors
2.3. Experiential Factors
2.4. Perceptual Learning
2.5. Speech Production
3. Processing of Degraded Speech in Brain Disorders
3.1. Traumatic Brain Injury
3.2. Stroke Aphasia
3.3. Parkinson’s Disease
3.4. Alzheimer’s Disease
3.5. Primary Progressive Aphasia
4. A Predictive Coding Model of Degraded Speech Processing in Primary Progressive Aphasia
5. Therapeutic Approaches
6. A Critique of the Predictive Coding Paradigm of Degraded Speech Processing
7. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hervais-Adelman, A.; Davis, M.H.; Johnsrude, I.S.; Carlyon, R.P. Perceptual learning of noise vocoded words: Effects of feedback and lexicality. J. Exp. Psychol. Hum. Percept. Perform. 2008, 34, 460–474. [Google Scholar] [CrossRef] [PubMed]
- Mattys, S.L.; Davis, M.H.; Bradlow, A.R.; Scott, S.K. Speech recognition in adverse conditions: A review. Lang. Cogn. Process. 2012, 27, 953–978. [Google Scholar] [CrossRef]
- Adank, P.; Davis, M.H.; Hagoort, P. Neural dissociation in processing noise and accent in spoken language comprehension. Neuropsychologia 2012, 50, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.H.; Sohoglu, E. Three functions of prediction error for Bayesian inference in speech perception. In The Cognitive Neurosciences, 6th ed.; Gazzaniga, M., Mangun, R.D.P., Eds.; MIT Press: Camridge, MA, USA, 2020; p. 177. [Google Scholar]
- Hickok, G.; Poeppel, D. The cortical organization of speech processing. Nat. Rev. Neurosci. 2007, 8, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Rong, F.; Venezia, J.; Matchin, W.; Hsieh, I.H.; Saberi, K.; Serences, J.T.; Hickok, G. Hierarchical organization of human auditory cortex: Evidence from acoustic invariance in the response to intelligible speech. Cereb. Cortex 2010, 20, 2486–2495. [Google Scholar] [CrossRef]
- Peelle, J.E. Hierarchical processing for speech in human auditory cortex and beyond. Front. Hum. Neurosci. 2010. [Google Scholar] [CrossRef]
- Scott, S.K.; Blank, C.C.; Rosen, S.; Wise, R.J. Identification of a pathway for intelligible speech in the left temporal lobe. Brain 2000, 123, 2400–2406. [Google Scholar] [CrossRef]
- Bregman, A.S. Auditory Scene Analysis: The Perceptual Organization of Sound; MIT Press: Camridge, MA, USA, 1990. [Google Scholar]
- Davis, M.H.; Johnsrude, I.S. Hearing speech sounds: Top-down influences on the interface between audition and speech perception. Hear. Res. 2007, 229, 132–147. [Google Scholar] [CrossRef]
- Ainley, V.; Apps, M.A.J.; Fotopoulou, A.; Tsakiris, M. ‘Bodily precision’: A predictive coding account of individual differences in interoceptive accuracy. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20160003. [Google Scholar] [CrossRef]
- Friston, K. A theory of cortical responses. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 815–836. [Google Scholar] [CrossRef]
- Arnal, L.H.; Wyart, V.; Giraud, A.-L. Transitions in neural oscillations reflect prediction errors generated in audiovisual speech. Nat. Neurosci. 2011, 14, 797–801. [Google Scholar] [CrossRef]
- Heilbron, M.; Chait, M. Great Expectations: Is there Evidence for Predictive Coding in Auditory Cortex. Neuroscience 2018, 389, 54–73. [Google Scholar] [CrossRef] [PubMed]
- Kocagoncu, E.; Klimovich-Gray, A.; Hughes, L.; Rowe, J.B. Evidence and implications of abnormal predictive coding in dementia. arXiv 2020, arXiv:2006.06311. [Google Scholar]
- Kashino, M. Phonemic restoration: The brain creates missing speech sounds. Acoust. Sci. Technol. 2006, 6, 4. [Google Scholar] [CrossRef]
- Sohoglu, E.; Peelle, J.E.; Carlyon, R.P.; Davis, M.H. Top-down influences of written text on perceived clarity of degraded speech. J. Exp. Psychol. Hum. Percept. Perform. 2014, 40, 186–199. [Google Scholar] [CrossRef]
- Alain, C.; Du, Y.; Bernstein, L.J.; Barten, T.; Banai, K. Listening under difficult conditions: An activation likelihood estimation meta-analysis. Hum. Brain Mapp. 2018, 39, 2695–2709. [Google Scholar] [CrossRef] [PubMed]
- Di Liberto, G.M.; Lalor, E.C.; Millman, R.E. Causal cortical dynamics of a predictive enhancement of speech intelligibility. NeuroImage 2018, 166, 247–258. [Google Scholar] [CrossRef]
- Johnsrude, I.S. The neuropsychological consequences of temporal lobe lesions. In Cognitive Deficits in Brain Disorders; Harrison, J.E., Owen, A.M., Eds.; Martin Dunitz Ltd.: London, UK, 2002; pp. 37–57. [Google Scholar]
- Strange, B.A.; Otten, L.J.; Josephs, O.; Rugg, M.D.; Dolan, R.J. Dissociable Human Perirhinal, Hippocampal, and Parahippocampal Roles during Verbal Encoding. J. Neurosci. 2002, 22, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Binney, R.J.; Embleton, K.V.; Jefferies, E.; Parker, G.J.; Ralph, M.A. The ventral and inferolateral aspects of the anterior temporal lobe are crucial in semantic memory: Evidence from a novel direct comparison of distortion-corrected fMRI, rTMS, and semantic dementia. Cereb. Cortex 2010, 20, 2728–2738. [Google Scholar] [CrossRef]
- Lambon Ralph, M.A.; Patterson, K. Generalization and Differentiation in Semantic Memory. Ann. N. Y. Acad. Sci. 2008, 1124, 61–76. [Google Scholar] [CrossRef]
- Pobric, G.; Jefferies, E.; Ralph, M.A.L. Anterior temporal lobes mediate semantic representation: Mimicking semantic dementia by using rTMS in normal participants. Proc. Natl. Acad. Sci. USA 2007, 104, 20137–20141. [Google Scholar] [CrossRef] [PubMed]
- Awad, M.; Warren, J.E.; Scott, S.K.; Turkheimer, F.E.; Wise, R.J.S. A Common System for the Comprehension and Production of Narrative Speech. J. Neurosci. 2007, 27, 11455–11464. [Google Scholar] [CrossRef]
- Rodd, J.M.; Davis, M.H.; Johnsrude, I.S. The neural mechanisms of speech comprehension: fMRI studies of semantic ambiguity. Cereb. Cortex 2005, 15, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Erb, J.; Henry, M.J.; Eisner, F.; Obleser, J. The Brain Dynamics of Rapid Perceptual Adaptation to Adverse Listening Conditions. J. Neurosci. 2013, 33, 10688–10697. [Google Scholar] [CrossRef]
- Hagoort, P. On Broca, brain, and binding: A new framework. Trends Cogn. Sci. 2005, 9, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Wild, C.J.; Yusuf, A.; Wilson, D.E.; Peelle, J.E.; Davis, M.H.; Johnsrude, I.S. Effortful Listening: The Processing of Degraded Speech Depends Critically on Attention. J. Neurosci. 2012, 32, 14010–14021. [Google Scholar] [CrossRef]
- Adank, P.; Nuttall, H.E.; Banks, B.; Kennedy-Higgins, D. Neural bases of accented speech perception. Front. Hum. Neurosci. 2015, 9. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Sakai, K.L. Brain activations during conscious self-monitoring of speech production with delayed auditory feedback: An fMRI study. Hum. Brain Mapp. 2003, 20, 22–28. [Google Scholar] [CrossRef]
- Hirnstein, M.; Westerhausen, R.; Korsnes, M.S.; Hugdahl, K. Sex differences in language asymmetry are age-dependent and small: A large-scale, consonant-vowel dichotic listening study with behavioral and fMRI data. Cortex 2013, 49, 1910–1921. [Google Scholar] [CrossRef]
- Davis, M.H.; Johnsrude, I.S. Hierarchical Processing in Spoken Language Comprehension. J. Neurosci. 2003, 23, 3423–3431. [Google Scholar] [CrossRef] [PubMed]
- Hervais-Adelman, A.G.; Carlyon, R.P.; Johnsrude, I.S.; Davis, M.H. Brain regions recruited for the effortful comprehension of noise-vocoded words. Lang. Cogn. Process. 2012, 27, 1145–1166. [Google Scholar] [CrossRef]
- Scott, S.K.; Rosen, S.; Lang, H.; Wise, R.J.S. Neural correlates of intelligibility in speech investigated with noise vocoded speech—A positron emission tomography study. J. Acoust. Soc. Am. 2006, 120, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Sunami, K.; Ishii, A.; Takano, S.; Yamamoto, H.; Sakashita, T.; Tanaka, M.; Watanabe, Y.; Yamane, H. Neural mechanisms of phonemic restoration for speech comprehension revealed by magnetoencephalography. Brain Res. 2013, 1537, 164–173. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Möttönen, R.; Calvert, G.A.; Jääskeläinen, I.P.; Matthews, P.M.; Thesen, T.; Tuomainen, J.; Sams, M. Perceiving identical sounds as speech or non-speech modulates activity in the left posterior superior temporal sulcus. Neuroimage 2006, 30, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-H.; Li, C.-W.; Wu, C.-W.; Chen, J.-H.; Liu, T.-C. Aging Effects on the Activation of the Auditory Cortex during Binaural Speech Listening in White Noise: An fMRI Study. Audiol. Neurotol. 2007, 12, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Adank, P.; Devlin, J.T. On-line plasticity in spoken sentence comprehension: Adapting to time-compressed speech. Neuroimage 2010, 49, 1124–1132. [Google Scholar] [CrossRef] [PubMed]
- Hartwigsen, G.; Golombek, T.; Obleser, J. Repetitive transcranial magnetic stimulation over left angular gyrus modulates the predictability gain in degraded speech comprehension. Cortex 2015, 68, 100–110. [Google Scholar] [CrossRef]
- Shahin, A.J.; Bishop, C.W.; Miller, L.M. Neural mechanisms for illusory filling-in of degraded speech. NeuroImage 2009, 44, 1133–1143. [Google Scholar] [CrossRef] [PubMed]
- Guediche, S.; Reilly, M.; Santiago, C.; Laurent, P.; Blumstein, S.E. An fMRI study investigating effects of conceptually related sentences on the perception of degraded speech. Cortex 2016, 79, 57–74. [Google Scholar] [CrossRef]
- Leonard, M.K.; Baud, M.O.; Sjerps, M.J.; Chang, E.F. Perceptual restoration of masked speech in human cortex. Nat. Commun. 2016, 7, 13619. [Google Scholar] [CrossRef]
- Becker, R.; Pefkou, M.; Michel, C.M.; Hervais-Adelman, A.G. Left temporal alpha-band activity reflects single word intelligibility. Front. Syst. Neurosci. 2013, 7, 121. [Google Scholar] [CrossRef]
- Arnal, L.H.; Doelling, K.B.; Poeppel, D. Delta-Beta Coupled Oscillations Underlie Temporal Prediction Accuracy. Cereb. Cortex 2015, 25, 3077–3085. [Google Scholar] [CrossRef] [PubMed]
- Hovsepyan, S.; Olasagasti, I.; Giraud, A.-L. Combining predictive coding and neural oscillations enables online syllable recognition in natural speech. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- Sedley, W.; Gander, P.E.; Kumar, S.; Kovach, C.K.; Oya, H.; Kawasaki, H.; Howard, M.A.; Griffiths, T.D. Neural signatures of perceptual inference. eLife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Hakonen, M.; May, P.J.C.; Alho, J.; Alku, P.; Jokinen, E.; Jääskeläinen, I.P.; Tiitinen, H. Previous exposure to intact speech increases intelligibility of its digitally degraded counterpart as a function of stimulus complexity. Neuroimage 2016, 125, 131–143. [Google Scholar] [CrossRef]
- Sohoglu, E.; Davis, M.H. Perceptual learning of degraded speech by minimizing prediction error. Proc. Natl. Acad. Sci. USA 2016, 113, E1747–E1756. [Google Scholar] [CrossRef]
- Samuel, A.G. Speech Perception. Annu. Rev. Psychol. 2011, 62, 49–72. [Google Scholar] [CrossRef]
- Warren, J.D.; Rohrer, J.D.; Schott, J.M.; Fox, N.C.; Hardy, J.; Rossor, M.N. Molecular nexopathies: A new paradigm of neurodegenerative disease. Trends Neurosci. 2013, 36, 561–569. [Google Scholar] [CrossRef]
- Hardy, C.J.; Marshall, C.R.; Golden, H.L.; Clark, C.N.; Mummery, C.J.; Griffiths, T.D.; Bamiou, D.E.; Warren, J.D. Hearing and dementia. J. Neurol. 2016, 263, 2339–2354. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, P.D.; Downey, L.E.; Agustus, J.L.; Hailstone, J.C.; Tyndall, M.H.; Cifelli, A.; Schott, J.M.; Warrington, E.K.; Warren, J.D. Agnosia for accents in primary progressive aphasia. Neuropsychologia 2013, 51, 1709–1715. [Google Scholar] [CrossRef]
- Floccia, C.; Butler, J.; Goslin, J.; Ellis, L. Regional and Foreign Accent Processing in English: Can Listeners Adapt? J. Psycholinguist. Res. 2009, 38, 379–412. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B. Speech-in-noise tests: How and why to include them in your basic test battery. Hear. J. 2003, 56, 42–46. [Google Scholar] [CrossRef]
- Lidestam, B.; Holgersson, J.; Moradi, S. Comparison of informational vs. energetic masking effects on speechreading performance. Front. Psychol. 2014, 5, 639. [Google Scholar] [CrossRef] [PubMed]
- Warren, R.M. Perceptual restoration of missing speech sounds. Science 1970, 167, 392–393. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.H.; Johnsrude, I.S.; Hervais-Adelman, A.; Taylor, K.; McGettigan, C. Lexical information drives perceptual learning of distorted speech: Evidence from the comprehension of noise-vocoded sentences. J. Exp. Psychol. Gen. 2005, 134, 222–241. [Google Scholar] [CrossRef]
- Shannon, R.V.; Zeng, F.G.; Kamath, V.; Wygonski, J.; Ekelid, M. Speech Recognition with Primarily Temporal Cues. Science 1995, 270, 303–304. [Google Scholar] [CrossRef]
- Dupoux, E.; Green, K. Perceptual adjustment to highly compressed speech: Effects of talker and rate changes. J. Exp. Psychol Hum. Percept. Perform. 1997, 23, 914–927. [Google Scholar] [CrossRef]
- Fausto, B.A.; Badana, A.N.S.; Arnold, M.L.; Lister, J.J.; Edwards, J.D. Comparison of Subjective and Objective Measures of Hearing, Auditory Processing, and Cognition Among Older Adults With and Without Mild Cognitive Impairment. J. Speech Lang. Hear. Res. 2018, 61, 945–956. [Google Scholar] [CrossRef]
- Foulke, E.; Sticht, T.G. Review of research on the intelligibility and comprehension of accelerated speech. Psychol. Bull. 1969, 72, 50–62. [Google Scholar] [CrossRef]
- Remez, R.; Rubin, P.; Pisoni, D.; Carrell, T. Speech perception without traditional speech cues. Science 1981, 212, 947–949. [Google Scholar] [CrossRef]
- Barker, J.; Cooke, M. Is the sine-wave speech cocktail party worth attending? Speech Commun. 1999, 27, 159–174. [Google Scholar] [CrossRef]
- Bent, T.; Bradlow, A.R. The interlanguage speech intelligibility benefit. J. Acoust. Soc. Am. 2003, 114, 1600–1610. [Google Scholar] [CrossRef]
- Clarke, C.M.; Garrett, M.F. Rapid adaptation to foreign-accented English. J. Acoust. Soc. Am. 2004, 116, 3647–3658. [Google Scholar] [CrossRef] [PubMed]
- Siegel, G.M.; Pick, H.L., Jr. Auditory feedback in the regulation of voice. J. Acoust. Soc. Am. 1974, 56, 1618–1624. [Google Scholar] [CrossRef]
- Jones, J.A.; Munhall, K.G. Perceptual calibration of F0 production: Evidence from feedback perturbation. J. Acoust. Soc. Am. 2000, 108, 1246–1251. [Google Scholar] [CrossRef]
- Donath, T.M.; Natke, U.; Kalveram, K.T. Effects of frequency-shifted auditory feedback on voice F0 contours in syllables. J. Acoust. Soc. Am. 2002, 111, 357–366. [Google Scholar] [CrossRef]
- Stuart, A.; Kalinowski, J.; Rastatter, M.P.; Lynch, K. Effect of delayed auditory feedback on normal speakers at two speech rates. J. Acoust. Soc. Am. 2002, 111, 2237–2241. [Google Scholar] [CrossRef] [PubMed]
- Moray, N. Attention in Dichotic Listening: Affective Cues and the Influence of Instructions. Q. J. Exp. Psychol. 1959, 11, 56–60. [Google Scholar] [CrossRef]
- Lewis, J.L. Semantic processing of unattended messages using dichotic listening. J. Exp. Psychol. 1970, 85, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.; Simon, J.Z. Neural coding of continuous speech in auditory cortex during monaural and dichotic listening. J. Neurophysiol. 2012, 107, 78–89. [Google Scholar] [CrossRef]
- Samuel, A.G. Phonemic restoration: Insights from a new methodology. J. Exp. Psychol. Gen. 1981, 110, 474–494. [Google Scholar] [CrossRef]
- Pichora-Fuller, M.K.; Schneider, B.A.; Daneman, M. How young and old adults listen to and remember speech in noise. J. Acoust. Soc. Am. 1995, 97, 593–608. [Google Scholar] [CrossRef]
- Parbery-Clark, A.; Skoe, E.; Lam, C.; Kraus, N. Musician enhancement for speech-in-noise. Ear Hear. 2009, 30, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.; White-Schwoch, T.; Parbery-Clark, A.; Kraus, N. A dynamic auditory-cognitive system supports speech-in-noise perception in older adults. Hear. Res. 2013, 300, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Poldrack, R.A.; Temple, E.; Protopapas, A.; Nagarajan, S.; Tallal, P.; Merzenich, M.; Gabrieli, J.D. Relations between the neural bases of dynamic auditory processing and phonological processing: Evidence from fMRI. J. Cogn. Neurosci. 2001, 13, 687–697. [Google Scholar] [CrossRef]
- Peelle, J.E.; McMillan, C.; Moore, P.; Grossman, M.; Wingfield, A. Dissociable patterns of brain activity during comprehension of rapid and syntactically complex speech: Evidence from fMRI. Brain Lang. 2004, 91, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Gallun, F.J.; Diedesch, A.C.; Kubli, L.R.; Walden, T.C.; Folmer, R.L.; Lewis, M.S.; McDermott, D.J.; Fausti, S.A.; Leek, M.R. Performance on tests of central auditory processing by individuals exposed to high-intensity blasts. J. Rehabil. Res. Dev. 2012, 49, 1005. [Google Scholar] [CrossRef] [PubMed]
- Saunders, G.H.; Frederick, M.T.; Arnold, M.; Silverman, S.; Chisolm, T.H.; Myers, P. Auditory difficulties in blast-exposed Veterans with clinically normal hearing. J. Rehabil. Res. Dev. 2015, 52, 343–360. [Google Scholar] [CrossRef]
- Gallun, F.J.; Lewis, M.S.; Folmer, R.L.; Hutter, M.; Papesh, M.A.; Belding, H.; Leek, M.R. Chronic effects of exposure to high-intensity blasts: Results of tests of central auditory processing. J. Rehabil. Res. Dev. 2016, 53, 15. [Google Scholar] [CrossRef]
- Papesh, M.A.; Elliott, J.E.; Callahan, M.L.; Storzbach, D.; Lim, M.M.; Gallun, F.J. Blast Exposure Impairs Sensory Gating: Evidence from Measures of Acoustic Startle and Auditory Event-Related Potentials. J. Neurotrauma 2019, 36, 702–712. [Google Scholar] [CrossRef]
- Bamiou, D.E.; Musiek, F.E.; Stow, I.; Stevens, J.; Cipolotti, L.; Brown, M.M.; Luxon, L.M. Auditory temporal processing deficits in patients with insular stroke. Neurology 2006, 67, 614–619. [Google Scholar] [CrossRef]
- Dunton, J.; Bruce, C.; Newton, C. Investigating the impact of unfamiliar speaker accent on auditory comprehension in adults with aphasia. Int. J. Lang. Commun. Disord. 2011, 46, 63–73. [Google Scholar] [CrossRef]
- Jacks, A.; Haley, K.L. Auditory Masking Effects on Speech Fluency in Apraxia of Speech and Aphasia: Comparison to Altered Auditory Feedback. J. Speech Lang. Hear. Res. 2015, 58, 1670–1686. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, E.Q.; Metman, L.V.; Larson, C.R. Vocal responses to perturbations in voice auditory feedback in individuals with Parkinson’s disease. PLoS ONE 2012, 7, e33629. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, X.; Wang, E.Q.; Chen, L.; Li, W.; Chen, Z.; Liu, H. Sensorimotor control of vocal pitch production in Parkinson’s disease. Brain Res. 2013, 1527, 99–107. [Google Scholar] [CrossRef]
- Gates, G.A.; Anderson, M.L.; Feeney, M.P.; McCurry, S.M.; Larson, E.B. Central Auditory Dysfunction in Older Persons With Memory Impairment or Alzheimer Dementia. Arch. Otolaryngol. Head Neck Surg. 2008, 134, 771. [Google Scholar] [CrossRef]
- Golden, H.L.; Agustus, J.L.; Goll, J.C.; Downey, L.E.; Mummery, C.J.; Schott, J.M.; Crutch, S.J.; Warren, J.D. Functional neuroanatomy of auditory scene analysis in Alzheimer’s disease. Neuroimage Clin. 2015, 7, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, K.G.; Gill, J.S.; Kothare, H.; Beagle, A.J.; Mizuiri, D.; Honma, S.M.; Gorno-Tempini, M.L.; Miller, B.L.; Vossel, K.A.; Nagarajan, S.S.; et al. Abnormal vocal behavior predicts executive and memory deficits in Alzheimer’s disease. Neurobiol. Aging 2017, 52, 71–80. [Google Scholar] [CrossRef]
- Hailstone, J.C.; Ridgway, G.R.; Bartlett, J.W.; Goll, J.C.; Crutch, S.J.; Warren, J.D. Accent processing in dementia. Neuropsychologia 2012, 50, 2233–2244. [Google Scholar] [CrossRef]
- Cope, T.E.; Sohoglu, E.; Sedley, W.; Patterson, K.; Jones, P.S.; Wiggins, J.; Dawson, C.; Grube, M.; Carlyon, R.P.; Griffiths, T.D.; et al. Evidence for causal top-down frontal contributions to predictive processes in speech perception. Nat. Commun. 2017, 8, 2154. [Google Scholar] [CrossRef] [PubMed]
- Hardy, C.J.D.; Marshall, C.R.; Bond, R.L.; Russell, L.L.; Dick, K.; Ariti, C.; Thomas, D.L.; Ross, S.J.; Agustus, J.L.; Crutch, S.J.; et al. Retained capacity for perceptual learning of degraded speech in primary progressive aphasia and Alzheimer’s disease. Alzheimers Res. Ther. 2018, 10, 70. [Google Scholar] [CrossRef] [PubMed]
- Gordon-Salant, S.; Fitzgibbons, P.J. Temporal factors and speech recognition performance in young and elderly listeners. J. Speech Hear. Res. 1993, 36, 1276–1285. [Google Scholar] [CrossRef]
- Frisina, D.R.; Frisina, R.D. Speech recognition in noise and presbycusis: Relations to possible neural mechanisms. Hear. Res. 1997, 106, 95–104. [Google Scholar] [CrossRef]
- Ross, B.; Dobri, S.; Schumann, A. Speech-in-noise understanding in older age: The role of inhibitory cortical responses. Eur. J. Neurosci. 2020, 51, 891–908. [Google Scholar] [CrossRef]
- Roth, T.N. Aging of the auditory system. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 129, pp. 357–373. [Google Scholar]
- Bidelman, G.M.; Howell, M. Functional changes in inter- and intra-hemispheric cortical processing underlying degraded speech perception. Neuroimage 2016, 124, 581–590. [Google Scholar] [CrossRef]
- Henry, M.J.; Herrmann, B.; Kunke, D.; Obleser, J. Aging affects the balance of neural entrainment and top-down neural modulation in the listening brain. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Gates, G.A.; Mills, J.H. Presbycusis. Lancet 2005, 366, 1111–1120. [Google Scholar] [CrossRef]
- Kane, M.J.; Hasher, L.; Stoltzfus, E.R.; Zacks, R.T.; Connelly, S.L. Inhibitory attentional mechanisms and aging. Psychol. Aging 1994, 9, 103–112. [Google Scholar] [CrossRef]
- Salthouse, T.A. The processing-speed theory of adult age differences in cognition. Psychol. Rev. 1996, 103, 403–428. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, R.; Albert, M.; Belleville, S.; Craik, F.I.M.; Duarte, A.; Grady, C.L.; Lindenberger, U.; Nyberg, L.; Park, D.C.; Reuter-Lorenz, P.A.; et al. Maintenance, reserve and compensation: The cognitive neuroscience of healthy ageing. Nat. Rev. Neurosci. 2018, 19, 701–710. [Google Scholar] [CrossRef]
- Humes, L.E.; Dubno, J.R.; Gordon-Salant, S.; Lister, J.J.; Cacace, A.T.; Cruickshanks, K.J.; Gates, G.A.; Wilson, R.H.; Wingfield, A. Central Presbycusis: A Review and Evaluation of the Evidence. J. Am. Acad. Audiol. 2012, 23, 635–666. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.; Roque, L.; Gaskins, C.R.; Gordon-Salant, S.; Goupell, M.J. Age-Related Compensation Mechanism Revealed in the Cortical Representation of Degraded Speech. J. Assoc. Res. Otolaryngol. 2020, 21, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Pichora-Fuller, M.K. Use of supportive context by younger and older adult listeners: Balancing bottom-up and top-down information processing. Int. J. Audiol. 2008, 47, S72–S82. [Google Scholar] [CrossRef]
- Saija, J.D.; Akyürek, E.G.; Andringa, T.C.; Başkent, D. Perceptual restoration of degraded speech is preserved with advancing age. J. Assoc. Res. Otolaryngol. 2014, 15, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Wolpe, N.; Ingram, J.N.; Tsvetanov, K.A.; Geerligs, L.; Kievit, R.A.; Henson, R.N.; Wolpert, D.M.; Rowe, J.B. Ageing increases reliance on sensorimotor prediction through structural and functional differences in frontostriatal circuits. Nat. Commun. 2016, 7, 13034. [Google Scholar] [CrossRef] [PubMed]
- Arlinger, S.; Lunner, T.; Lyxell, B.; Pichora-Fuller, M.K. The emergence of cognitive hearing science. Scand. J. Psychol. 2009, 50, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Peelle, J.E. Listening Effort: How the Cognitive Consequences of Acoustic Challenge Are Reflected in Brain and Behavior. Ear Hear. 2018, 39, 204–214. [Google Scholar] [CrossRef]
- O’Brien, J.L.; Lister, J.J.; Fausto, B.A.; Morgan, D.G.; Maeda, H.; Andel, R.; Edwards, J.D. Are auditory processing and cognitive performance assessments overlapping or distinct? Parsing the auditory behaviour of older adults. Int. J. Audiol. 2021, 60, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Auksztulewicz, R.; Friston, K. Attentional Enhancement of Auditory Mismatch Responses: A DCM/MEG Study. Cereb. Cortex 2015, 25, 4273–4283. [Google Scholar] [CrossRef] [PubMed]
- Foxe, J.J.; Snyder, A.C. The Role of Alpha-Band Brain Oscillations as a Sensory Suppression Mechanism during Selective Attention. Front. Psychol. 2011, 2, 154. [Google Scholar] [CrossRef] [PubMed]
- Jacquemot, C.; Scott, S.K. What is the relationship between phonological short-term memory and speech processing? Trends Cogn. Sci. 2006, 10, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Erb, J.; Henry, M.J.; Eisner, F.; Obleser, J. Auditory skills and brain morphology predict individual differences in adaptation to degraded speech. Neuropsychologia 2012, 50, 2154–2164. [Google Scholar] [CrossRef]
- Puschmann, S.; Baillet, S.; Zatorre, R.J. Musicians at the Cocktail Party: Neural Substrates of Musical Training During Selective Listening in Multispeaker Situations. Cereb. Cortex 2019, 29, 3253–3265. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Choi, I.; Schwalje, A.T.; Kim, K.; Lee, J.H. Auditory Working Memory Explains Variance in Speech Recognition in Older Listeners Under Adverse Listening Conditions. Clin. Interv. Aging 2020, 15, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.J.; Wostmann, M.; Obleser, J. Selective Attention to Auditory Memory Neurally Enhances Perceptual Precision. J. Neurosci. 2015, 35, 16094–16104. [Google Scholar] [CrossRef]
- Akeroyd, M.A. Are individual differences in speech reception related to individual differences in cognitive ability? A survey of twenty experimental studies with normal and hearing-impaired adults. Int. J. Audiol. 2008, 47 (Suppl. 2), S53–S71. [Google Scholar] [CrossRef] [PubMed]
- Souza, P.; Arehart, K. Robust relationship between reading span and speech recognition in noise. Int. J. Audiol. 2015, 54, 705–713. [Google Scholar] [CrossRef]
- Donhauser, P.W.; Baillet, S. Two Distinct Neural Timescales for Predictive Speech Processing. Neuron 2020, 105, 385–393. [Google Scholar] [CrossRef]
- Alain, C.; Zendel, B.R.; Hutka, S.; Bidelman, G.M. Turning down the noise: The benefit of musical training on the aging auditory brain. Hear. Res. 2014, 308, 162–173. [Google Scholar] [CrossRef]
- Bidelman, G.M.; Krishnan, A. Effects of reverberation on brainstem representation of speech in musicians and non-musicians. Brain Res. 2010, 1355, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.L.; Lerch, J.; Norton, A.; Forgeard, M.; Winner, E.; Evans, A.C.; Schlaug, G. Musical Training Shapes Structural Brain Development. J. Neurosci. 2009, 29, 3019–3025. [Google Scholar] [CrossRef] [PubMed]
- Koelsch, S.; Schröger, E.; Tervaniemi, M. Superior pre-attentive auditory processing in musicians. Neuroreport 1999, 10, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Kraus, N.; Skoe, E.; Parbery-Clark, A.; Ashley, R. Experience-induced malleability in neural encoding of pitch, timbre, and timing. Ann. N. Y. Acad. Sci. 2009, 1169, 543–557. [Google Scholar] [CrossRef]
- Kraus, N.; Strait, D.L.; Parbery-Clark, A. Cognitive factors shape brain networks for auditory skills: Spotlight on auditory working memory. Ann. N. Y. Acad. Sci. 2012, 1252, 100–107. [Google Scholar] [CrossRef]
- Magne, C.; Schön, D.; Besson, M. Musician children detect pitch violations in both music and language better than nonmusician children: Behavioral and electrophysiological approaches. J. Cogn. Neurosci. 2006, 18, 199–211. [Google Scholar] [CrossRef]
- Musacchia, G.; Sams, M.; Skoe, E.; Kraus, N. Musicians have enhanced subcortical auditory and audiovisual processing of speech and music. Proc. Natl. Acad. Sci. USA 2007, 104, 15894–15898. [Google Scholar] [CrossRef] [PubMed]
- Schön, D.; Magne, C.; Besson, M. The music of speech: Music training facilitates pitch processing in both music and language. Psychophysiology 2004, 41, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.C.; Skoe, E.; Russo, N.M.; Dees, T.; Kraus, N. Musical experience shapes human brainstem encoding of linguistic pitch patterns. Nat. Neurosci. 2007, 10, 420–422. [Google Scholar] [CrossRef]
- Başkent, D.; Gaudrain, E. Musician advantage for speech-on-speech perception. J. Acoust. Soc. Am. 2016, 139, El51–E156. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, J.; Mason, C.R.; Streeter, T.M.; Best, V.; Kidd, J.G.; Patel, A.D. Musical training, individual differences and the cocktail party problem. Sci. Rep. 2015, 5, 11628. [Google Scholar] [CrossRef]
- Zendel, B.R.; Alain, C. Musicians experience less age-related decline in central auditory processing. Psychol. Aging 2012, 27, 410–417. [Google Scholar] [CrossRef]
- Zendel, B.R.; Alain, C. Concurrent sound segregation is enhanced in musicians. J. Cogn. Neurosci. 2009, 21, 1488–1498. [Google Scholar] [CrossRef]
- Jin, S.-H.; Liu, C. English sentence recognition in speech-shaped noise and multi-talker babble for English-, Chinese-, and Korean-native listeners. J. Acoust. Soc. Am. 2012, 132, EL391–EL397. [Google Scholar] [CrossRef]
- Lucks Mendel, L.; Widner, H. Speech perception in noise for bilingual listeners with normal hearing. Int. J. Audiol. 2016, 55, 126–134. [Google Scholar] [CrossRef]
- Rammell, C.S.; Cheng, H.; Pisoni, D.B.; Newman, S.D. L2 speech perception in noise: An fMRI study of advanced Spanish learners. Brain Res. 2019, 1720, 146316. [Google Scholar] [CrossRef]
- Skoe, E.; Karayanidi, K. Bilingualism and Speech Understanding in Noise: Auditory and Linguistic Factors. J. Am. Acad. Audiol. 2019, 30, 115–130. [Google Scholar] [CrossRef]
- Bradlow, A.R.; Alexander, J.A. Semantic and phonetic enhancements for speech-in-noise recognition by native and non-native listeners. J. Acoust. Soc. Am. 2007, 121, 2339–2349. [Google Scholar] [CrossRef] [PubMed]
- Hervais-Adelman, A.; Pefkou, M.; Golestani, N. Bilingual speech-in-noise: Neural bases of semantic context use in the native language. Brain Lang. 2014, 132, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kousaie, S.; Baum, S.; Phillips, N.A.; Gracco, V.; Titone, D.; Chen, J.K.; Chai, X.J.; Klein, D. Language learning experience and mastering the challenges of perceiving speech in noise. Brain Lang. 2019, 196, 104645. [Google Scholar] [CrossRef]
- Eisner, F.; McGettigan, C.; Faulkner, A.; Rosen, S.; Scott, S.K. Inferior frontal gyrus activation predicts individual differences in perceptual learning of cochlear-implant simulations. J. Neurosci. 2010, 30, 7179–7186. [Google Scholar] [CrossRef]
- Goldstone, R.L. PERCEPTUAL LEARNING. Annu. Rev. Psychol. 1998, 49, 585–612. [Google Scholar] [CrossRef]
- Samuel, A.G.; Kraljic, T. Perceptual learning for speech. Atten. Percept. Psychophys. 2009, 71, 1207–1218. [Google Scholar] [CrossRef]
- Eisner, F.; McQueen, J.M. Perceptual learning in speech: Stability over time. J. Acoust. Soc. Am. 2006, 119, 1950–1953. [Google Scholar] [CrossRef] [PubMed]
- Norris, D. Perceptual learning in speech. Cogn. Psychol. 2003, 47, 204–238. [Google Scholar] [CrossRef]
- Petrov, A.A.; Dosher, B.A.; Lu, Z.L. The dynamics of perceptual learning: An incremental reweighting model. Psychol. Rev. 2005, 112, 715–743. [Google Scholar] [CrossRef]
- Tsodyks, M.; Gilbert, C. Neural networks and perceptual learning. Nature 2004, 431, 775–781. [Google Scholar] [CrossRef]
- Bradlow, A.R.; Bent, T. Perceptual adaptation to non-native speech. Cognition 2008, 106, 707–729. [Google Scholar] [CrossRef]
- Eisner, F.; McQueen, J.M. The specificity of perceptual learning in speech processing. Percept. Psychophys. 2005, 67, 224–238. [Google Scholar] [CrossRef]
- Nygaard, L.C.; Pisoni, D.B. Talker-specific learning in speech perception. Percept. Psychophys. 1998, 60, 355–376. [Google Scholar] [CrossRef]
- Clopper, C.G.; Pisoni, D.B. Effects of talker variability on perceptual learning of dialects. Lang. Speech 2004, 47, 207–239. [Google Scholar] [CrossRef]
- Gordon-Salant, S.; Yeni-Komshian, G.H.; Fitzgibbons, P.J.; Schurman, J. Short-term adaptation to accented English by younger and older adults. J. Acoust. Soc. Am. 2010, 128, El200–E1204. [Google Scholar] [CrossRef]
- Sidaras, S.K.; Alexander, J.E.; Nygaard, L.C. Perceptual learning of systematic variation in Spanish-accented speech. J. Acoust. Soc. Am. 2009, 125, 3306–3316. [Google Scholar] [CrossRef] [PubMed]
- Stacey, P.C.; Summerfield, A.Q. Effectiveness of computer-based auditory training in improving the perception of noise-vocoded speech. J. Acoust. Soc. Am. 2007, 121, 2923–2935. [Google Scholar] [CrossRef] [PubMed]
- Banai, K.; Lavner, Y. The effects of training length on the perceptual learning of time-compressed speech and its generalization. J. Acoust. Soc. Am. 2014, 136, 1908–1917. [Google Scholar] [CrossRef] [PubMed]
- Francis, A.L.; Ciocca, V.; Ma, L.; Fenn, K. Perceptual learning of Cantonese lexical tones by tone and non-tone language speakers. J. Phon. 2008, 36, 268–294. [Google Scholar] [CrossRef]
- Peelle, J.E.; Wingfield, A. Dissociations in perceptual learning revealed by adult age differences in adaptation to time-compressed speech. J. Exp. Psychol. Hum. Percept. Perform. 2005, 31, 1315–1330. [Google Scholar] [CrossRef]
- Huyck, J.J.; Johnsrude, I.S. Rapid perceptual learning of noise-vocoded speech requires attention. J. Acoust. Soc. Am. 2012, 131, EL236–EL242. [Google Scholar] [CrossRef] [PubMed]
- Bradlow, A.R.; Pisoni, D.B.; Akahane-Yamada, R.; Tohkura, Y. Training Japanese listeners to identify English /r/ and /l/: IV. Some effects of perceptual learning on speech production. J. Acoust. Soc. Am. 1997, 101, 2299–2310. [Google Scholar] [CrossRef]
- Kraljic, T.; Samuel, A.G. Perceptual learning for speech: Is there a return to normal? Cogn. Psychol. 2005, 51, 141–178. [Google Scholar] [CrossRef]
- Houde, J.F.; Chang, E.F. The cortical computations underlying feedback control in vocal production. Curr. Opin. Neurobiol. 2015, 33, 174–181. [Google Scholar] [CrossRef]
- Houde, J.F.; Nagarajan, S.S. Speech production as state feedback control. Front. Hum. Neurosci. 2011, 5, 82. [Google Scholar] [CrossRef] [PubMed]
- Hickok, G. The cortical organization of speech processing: Feedback control and predictive coding the context of a dual-stream model. J. Commun. Disord. 2012, 45, 393–402. [Google Scholar] [CrossRef]
- Lincoln, M.; Packman, A.; Onslow, M. Altered auditory feedback and the treatment of stuttering: A review. J. Fluen. Disord. 2006, 31, 71–89. [Google Scholar] [CrossRef] [PubMed]
- Lombard, E. Le signe de l’elevation de la voix. Ann. Mal. De L’oreille Et Du Larynx 1911, 18, 101–119. [Google Scholar]
- Takaso, H.; Eisner, F.; Wise, R.J.; Scott, S.K. The effect of delayed auditory feedback on activity in the temporal lobe while speaking: A positron emission tomography study. J. Speech Lang. Hear. Res. 2010, 53, 226–236. [Google Scholar] [CrossRef]
- Meekings, S.; Evans, S.; Lavan, N.; Boebinger, D.; Krieger-Redwood, K.; Cooke, M.; Scott, S.K. Distinct neural systems recruited when speech production is modulated by different masking sounds. J. Acoust. Soc. Am. 2016, 140, 8. [Google Scholar] [CrossRef] [PubMed]
- Elder, G.A. Update on TBI and Cognitive Impairment in Military Veterans. Curr. Neurol. Neurosci. Rep. 2015, 15, 68. [Google Scholar] [CrossRef]
- Menon, D.K.; Schwab, K.; Wright, D.W.; Maas, A.I. Position statement: Definition of traumatic brain injury. Arch. Phys. Med. Rehabil. 2010, 91, 1637–1640. [Google Scholar] [CrossRef]
- Hoffer, M.E.; Balaban, C.; Nicholas, R.; Marcus, D.; Murphy, S.; Gottshall, K. Neurosensory Sequelae of Mild Traumatic Brain Injury. Psychiatr. Ann. 2013, 43, 5. [Google Scholar] [CrossRef]
- Gallun, F.J.; Papesh, M.A.; Lewis, M.S. Hearing complaints among veterans following traumatic brain injury. Brain Inj. 2017, 31, 1183–1187. [Google Scholar] [CrossRef]
- Turgeon, C.; Champoux, F.; Lepore, F.; Leclerc, S.; Ellemberg, D. Auditory processing after sport-related concussions. Ear Hear. 2011, 32, 667–670. [Google Scholar] [CrossRef]
- Hoover, E.C.; Souza, P.E.; Gallun, F.J. Auditory and Cognitive Factors Associated with Speech-in-Noise Complaints following Mild Traumatic Brain Injury. J. Am. Acad. Audiol. 2017, 28, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Boller, F.; Vrtunski, P.B.; Kim, Y.; Mack, J.L. Delayed auditory feedback and aphasia. Cortex 1978, 14, 212–226. [Google Scholar] [CrossRef]
- Chapin, C.; Blumstein, S.E.; Meissner, B.; Boller, F. Speech production mechanisms in aphasia: A delayed auditory feedback study. Brain Lang. 1981, 14, 106–113. [Google Scholar] [CrossRef]
- Fifer, R.C. Insular stroke causing unilateral auditory processing disorder: Case report. J. Am. Acad. Audiol. 1993, 4, 364–369. [Google Scholar]
- Huang, X.; Chen, X.; Yan, N.; Jones, J.A.; Wang, E.Q.; Chen, L.; Guo, Z.; Li, W.; Liu, P.; Liu, H. The impact of parkinson’s disease on the cortical mechanisms that support auditory-motor integration for voice control. Hum. Brain Mapp. 2016, 37, 4248–4261. [Google Scholar] [CrossRef]
- Tolosa, E.; Wenning, G.; Poewe, W. The diagnosis of Parkinson’s disease. Lancet Neurol. 2006, 5, 75–86. [Google Scholar] [CrossRef]
- Williams-Gray, C.H.; Mason, S.L.; Evans, J.R.; Foltynie, T.; Brayne, C.; Robbins, T.W.; Barker, R.A. The CamPaIGN study of Parkinson’s disease: 10-year outlook in an incident population-based cohort. J. Neurol. Neurosurg. Psychiatry 2013, 84, 1258–1264. [Google Scholar] [CrossRef]
- Sapir, S.; Ramig, L.; Fox, C. Speech and swallowing disorders in Parkinson disease. Curr. Opin. Otolaryngol. Head Neck Surg. 2008, 16, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.K.; Bradshaw, J.L.; Iansek, T. Volume perception in parkinsonian speech. Mov. Disord. 2000, 15, 1125–1131. [Google Scholar] [CrossRef]
- Arnold, C.; Gehrig, J.; Gispert, S.; Seifried, C.; Kell, C.A. Pathomechanisms and compensatory efforts related to Parkinsonian speech. Neuroimage Clin. 2014, 4, 82–97. [Google Scholar] [CrossRef]
- Ho, A.K.; Bradshaw, J.L.; Iansek, R.; Alfredson, R. Speech volume regulation in Parkinson’s disease: Effects of implicit cues and explicit instructions. Neuropsychologia 1999, 37, 1453–1460. [Google Scholar] [CrossRef]
- Kiran, S.; Larson, C.R. Effect of duration of pitch-shifted feedback on vocal responses in patients with Parkinson’s disease. J. Speech Lang. Hear. Res. 2001, 44, 975–987. [Google Scholar] [CrossRef]
- Mollaei, F.; Shiller, D.M.; Baum, S.R.; Gracco, V.L. Sensorimotor control of vocal pitch and formant frequencies in Parkinson’s disease. Brain Res. 2016, 1646, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Downie, A.W.; Low, J.M.; Lindsay, D.D. Speech Disorder in Parkinsonism—Usefulness of Delayed Auditory Feedback in Selected Cases. Int. J. Lang. Commun. Disord. 1981, 16, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Cools, R.; Rogers, R.; Barker, R.A.; Robbins, T.W. Top-down attentional control in Parkinson’s disease: Salient considerations. J. Cogn. Neurosci. 2010, 22, 848–859. [Google Scholar] [CrossRef]
- Zarkali, A.; Adams, R.A.; Psarras, S.; Leyland, L.A.; Rees, G.; Weil, R.S. Increased weighting on prior knowledge in Lewy body-associated visual hallucinations. Brain Commun. 2019, 1, fcz007. [Google Scholar] [CrossRef] [PubMed]
- Agosta, F.; Pievani, M.; Geroldi, C.; Copetti, M.; Frisoni, G.B.; Filippi, M. Resting state fMRI in Alzheimer’s disease: Beyond the default mode network. Neurobiol. Aging 2012, 33, 1564–1578. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Greicius, M.D.; Gennatas, E.D.; Growdon, M.E.; Jang, J.Y.; Rabinovici, G.D.; Kramer, J.H.; Weiner, M.; Miller, B.L.; Seeley, W.W. Divergent network connectivity changes in behavioural variant frontotemporal dementia and Alzheimer’s disease. Brain 2010, 133, 1352–1367. [Google Scholar] [CrossRef]
- Bouma, A.; Gootjes, L. Effects of attention on dichotic listening in elderly and patients with dementia of the Alzheimer type. Brain Cogn. 2011, 76, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Idrizbegovic, E.; Hederstierna, C.; Dahlquist, M.; Rosenhall, U. Short-term longitudinal study of central auditory function in Alzheimer’s disease and mild cognitive impairment. Dement. Geriatr. Cogn. Dis. Extra 2013, 3, 468–471. [Google Scholar] [CrossRef]
- Utoomprurkporn, N.; Hardy, C.J.D.; Stott, J.; Costafreda, S.G.; Warren, J.; Bamiou, D.E. “The Dichotic Digit Test” as an Index Indicator for Hearing Problem in Dementia: Systematic Review and Meta-Analysis. J. Am. Acad. Audiol. 2020, 31, 646–655. [Google Scholar] [CrossRef]
- Goll, J.C.; Kim, L.G.; Ridgway, G.R.; Hailstone, J.C.; Lehmann, M.; Buckley, A.H.; Crutch, S.J.; Warren, J.D. Impairments of auditory scene analysis in Alzheimer’s disease. Brain 2012, 135, 190–200. [Google Scholar] [CrossRef]
- Hardy, C.J.D.; Yong, K.X.X.; Goll, J.C.; Crutch, S.J.; Warren, J.D. Impairments of auditory scene analysis in posterior cortical atrophy. Brain 2020, 143, 2689–2695. [Google Scholar] [CrossRef]
- Gates, G.A.; Anderson, M.L.; McCurry, S.M.; Feeney, M.P.; Larson, E.B. Central auditory dysfunction as a harbinger of Alzheimer dementia. Arch. Otolaryngol. Head Neck Surg. 2011, 137, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Gates, G.A.; Gibbons, L.E.; McCurry, S.M.; Crane, P.K.; Feeney, M.P.; Larson, E.B. Executive dysfunction and presbycusis in older persons with and without memory loss and dementia. Cogn. Behav. Neurol. 2010, 23, 218–223. [Google Scholar] [CrossRef]
- Pronk, M.; Lissenberg-Witte, B.I.; Van der Aa, H.P.A.; Comijs, H.C.; Smits, C.; Lemke, U.; Zekveld, A.A.; Kramer, S.E. Longitudinal Relationships Between Decline in Speech-in-Noise Recognition Ability and Cognitive Functioning: The Longitudinal Aging Study Amsterdam. J. Speech Lang. Hear. Res. 2019, 62, 1167–1187. [Google Scholar] [CrossRef] [PubMed]
- Burda, A.N.; Hageman, C.F.; Brousard, K.T.; Miller, A.L. Dementia and identification of words and sentences produced by native and nonnative English speakers. Percept. Mot. Ski. 2004, 98, 1359–1362. [Google Scholar] [CrossRef]
- Hardy, C.J.D.; Hwang, Y.T.; Bond, R.L.; Marshall, C.R.; Ridha, B.H.; Crutch, S.J.; Rossor, M.N.; Warren, J.D. Donepezil enhances understanding of degraded speech in Alzheimer’s disease. Ann. Clin. Transl. Neurol. 2017, 4, 835–840. [Google Scholar] [CrossRef]
- Ranasinghe, K.G.; Kothare, H.; Kort, N.; Hinkley, L.B.; Beagle, A.J.; Mizuiri, D.; Honma, S.M.; Lee, R.; Miller, B.L.; Gorno-Tempini, M.L.; et al. Neural correlates of abnormal auditory feedback processing during speech production in Alzheimer’s disease. Sci. Rep. 2019, 9, 5686. [Google Scholar] [CrossRef]
- Marshall, C.R.; Hardy, C.J.D.; Volkmer, A.; Russell, L.L.; Bond, R.L.; Fletcher, P.D.; Clark, C.N.; Mummery, C.J.; Schott, J.M.; Rossor, M.N.; et al. Primary progressive aphasia: A clinical approach. J. Neurol. 2018, 265, 1474–1490. [Google Scholar] [CrossRef]
- Gorno-Tempini, M.L.; Hillis, A.E.; Weintraub, S.; Kertesz, A.; Mendez, M.; Cappa, S.F.; Ogar, J.M.; Rohrer, J.D.; Black, S.; Boeve, B.F.; et al. Classification of primary progressive aphasia and its variants. Neurology 2011, 76, 1006–1014. [Google Scholar] [CrossRef]
- Goll, J.C.; Kim, L.G.; Hailstone, J.C.; Lehmann, M.; Buckley, A.; Crutch, S.J.; Warren, J.D. Auditory object cognition in dementia. Neuropsychologia 2011, 49, 2755–2765. [Google Scholar] [CrossRef]
- Goll, J.C.; Crutch, S.J.; Loo, J.H.; Rohrer, J.D.; Frost, C.; Bamiou, D.E.; Warren, J.D. Non-verbal sound processing in the primary progressive aphasias. Brain 2010, 133, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Grube, M.; Bruffaerts, R.; Schaeverbeke, J.; Neyens, V.; De Weer, A.S.; Seghers, A.; Bergmans, B.; Dries, E.; Griffiths, T.D.; Vandenberghe, R. Core auditory processing deficits in primary progressive aphasia. Brain 2016, 139, 1817–1829. [Google Scholar] [CrossRef]
- Hardy, C.J.D.; Frost, C.; Sivasathiaseelan, H.; Johnson, J.C.S.; Agustus, J.L.; Bond, R.L.; Benhamou, E.; Russell, L.L.; Marshall, C.R.; Rohrer, J.D.; et al. Findings of Impaired Hearing in Patients With Nonfluent/Agrammatic Variant Primary Progressive Aphasia. JAMA Neurol. 2019, 76, 607–611. [Google Scholar] [CrossRef]
- Johnson, J.C.S.; Jiang, J.; Bond, R.L.; Benhamou, E.; Requena-Komuro, M.C.; Russell, L.L.; Greaves, C.; Nelson, A.; Sivasathiaseelan, H.; Marshall, C.R.; et al. Impaired phonemic discrimination in logopenic variant primary progressive aphasia. Ann. Clin. Transl. Neurol. 2020, 7, 1252–1257. [Google Scholar] [CrossRef] [PubMed]
- Ruksenaite, J.; Volkmer, A.; Jiang, J.; Johnson, J.C.; Marshall, C.R.; Warren, J.D.; Hardy, C.J. Primary Progressive Aphasia: Toward a Pathophysiological Synthesis. Curr. Neurol. Neurosci. Rep. 2021, 21. [Google Scholar] [CrossRef]
- Hardy, C.J.D.; Bond, R.L.; Jaisin, K.; Marshall, C.R.; Russell, L.L.; Dick, K.; Crutch, S.J.; Rohrer, J.D.; Warren, J.D. Sensitivity of Speech Output to Delayed Auditory Feedback in Primary Progressive Aphasias. Front. Neurol. 2018, 9, 894. [Google Scholar] [CrossRef] [PubMed]
- Hardy, C.J.D.; Agustus, J.L.; Marshall, C.R.; Clark, C.N.; Russell, L.L.; Bond, R.L.; Brotherhood, E.V.; Thomas, D.L.; Crutch, S.J.; Rohrer, J.D.; et al. Behavioural and neuroanatomical correlates of auditory speech analysis in primary progressive aphasias. Alzheimer’s Res. Ther. 2017, 9. [Google Scholar] [CrossRef]
- Hardy, C.J.D.; Agustus, J.L.; Marshall, C.R.; Clark, C.N.; Russell, L.L.; Brotherhood, E.V.; Bond, R.L.; Fiford, C.M.; Ondobaka, S.; Thomas, D.L.; et al. Functional neuroanatomy of speech signal decoding in primary progressive aphasias. Neurobiol. Aging 2017, 56, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Conway, E.R.; Chenery, H.J. Evaluating the MESSAGE Communication Strategies in Dementia training for use with community-based aged care staff working with people with dementia: A controlled pretest-post-test study. J. Clin. Nurs. 2016, 25, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Liddle, J.; Smith-Conway, E.R.; Baker, R.; Angwin, A.J.; Gallois, C.; Copland, D.A.; Pachana, N.A.; Humphreys, M.S.; Byrne, G.J.; Chenery, H.J. Memory and communication support strategies in dementia: Effect of a training program for informal caregivers. Int. Psychogeriatr. 2012, 24, 1927–1942. [Google Scholar] [CrossRef]
- Sparks, R.; Helm, N.; Albert, M. Aphasia rehabilitation resulting from melodic intonation therapy. Cortex 1974, 10, 303–316. [Google Scholar] [CrossRef]
- Zumbansen, A.; Peretz, I.; Hébert, S. Melodic Intonation Therapy: Back to Basics for Future Research. Front. Neurol. 2014, 5. [Google Scholar] [CrossRef]
- Bieber, R.E.; Gordon-Salant, S. Improving older adults’ understanding of challenging speech: Auditory training, rapid adaptation and perceptual learning. Hear. Res. 2021, 402, 108054. [Google Scholar] [CrossRef]
- Shafiro, V.; Sheft, S.; Gygi, B.; Ho, K.T. The influence of environmental sound training on the perception of spectrally degraded speech and environmental sounds. Trends Amplif. 2012, 16, 83–101. [Google Scholar] [CrossRef]
- Sehm, B.; Schnitzler, T.; Obleser, J.; Groba, A.; Ragert, P.; Villringer, A.; Obrig, H. Facilitation of inferior frontal cortex by transcranial direct current stimulation induces perceptual learning of severely degraded speech. J. Neurosci. 2013, 33, 15868–15878. [Google Scholar] [CrossRef]
- Rampersad, S.; Roig-Solvas, B.; Yarossi, M.; Kulkarni, P.P.; Santarnecchi, E.; Dorval, A.D.; Brooks, D.H. Prospects for transcranial temporal interference stimulation in humans: A computational study. Neuroimage 2019, 202, 116124. [Google Scholar] [CrossRef] [PubMed]
- Wayne, R.V.; Hamilton, C.; Jones Huyck, J.; Johnsrude, I.S. Working Memory Training and Speech in Noise Comprehension in Older Adults. Front. Aging Neurosci. 2016, 8, 49. [Google Scholar] [CrossRef]
- Escobar, J.; Mussoi, B.S.; Silberer, A.B. The Effect of Musical Training and Working Memory in Adverse Listening Situations. Ear Hear. 2020, 41, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Fu, X.; Luo, D.; Xing, L.; Du, Y. Musical Experience Offsets Age-Related Decline in Understanding Speech-in-Noise: Type of Training Does Not Matter, Working Memory Is the Key. Ear Hear. 2020, 42, 258–270. [Google Scholar] [CrossRef]
- Zendel, B.R.; West, G.L.; Belleville, S.; Peretz, I. Musical training improves the ability to understand speech-in-noise in older adults. Neurobiol. Aging 2019, 81, 102–115. [Google Scholar] [CrossRef]
- Cardin, V.; Rosen, S.; Konieczny, L.; Coulson, K.; Lametti, D.; Edwards, M.; Woll, B. The effect of dopamine on the comprehension of spectrally-shifted noise-vocoded speech: A pilot study. Int. J. Audiol. 2020, 59, 674–681. [Google Scholar] [CrossRef]
- De Boer, B. Self-Organisation and Evolution of Biological and Social Systems. In Self-Organisation and Evolution of Biological and Social Systems; Hemelrijk, C., Ed.; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Williams, D. Predictive coding and thought. Synthese 2020, 197, 1749–1775. [Google Scholar] [CrossRef]
- Koelsch, S.; Vuust, P.; Friston, K. Predictive Processes and the Peculiar Case of Music. Trends Cogn. Sci. 2019, 23, 63–77. [Google Scholar] [CrossRef]
- Sohoglu, E.; Peelle, J.E.; Carlyon, R.P.; Davis, M.H. Predictive top-down integration of prior knowledge during speech perception. J. Neurosci. 2012, 32, 8443–8453. [Google Scholar] [CrossRef]
- Wacongne, C.; Changeux, J.P.; Dehaene, S. A neuronal model of predictive coding accounting for the mismatch negativity. J. Neurosci. 2012, 32, 3665–3678. [Google Scholar] [CrossRef]
- Rysop, A.U.; Schmitt, L.M.; Obleser, J.; Hartwigsen, G. Neural modelling of the semantic predictability gain under challenging listening conditions. Hum. Brain Mapp. 2021, 42, 110–127. [Google Scholar] [CrossRef]
- Kogo, N.; Trengove, C. Is predictive coding theory articulated enough to be testable? Front. Comput. Neurosci. 2015, 9, 111. [Google Scholar] [CrossRef]
- Benhamou, E.; Marshall, C.R.; Russell, L.L.; Hardy, C.J.D.; Bond, R.L.; Sivasathiaseelan, H.; Greaves, C.V.; Friston, K.J.; Rohrer, J.D.; Warren, J.D.; et al. The neurophysiological architecture of semantic dementia: Spectral dynamic causal modelling of a neurodegenerative proteinopathy. Sci. Rep. 2020, 10, 16321. [Google Scholar] [CrossRef]
- Shaw, A.D.; Hughes, L.E.; Moran, R.; Coyle-Gilchrist, I.; Rittman, T.; Rowe, J.B. In Vivo Assay of Cortical Microcircuitry in Frontotemporal Dementia: A Platform for Experimental Medicine Studies. Cereb. Cortex 2021, 31, 1837–1847. [Google Scholar] [CrossRef]




{kind=link}
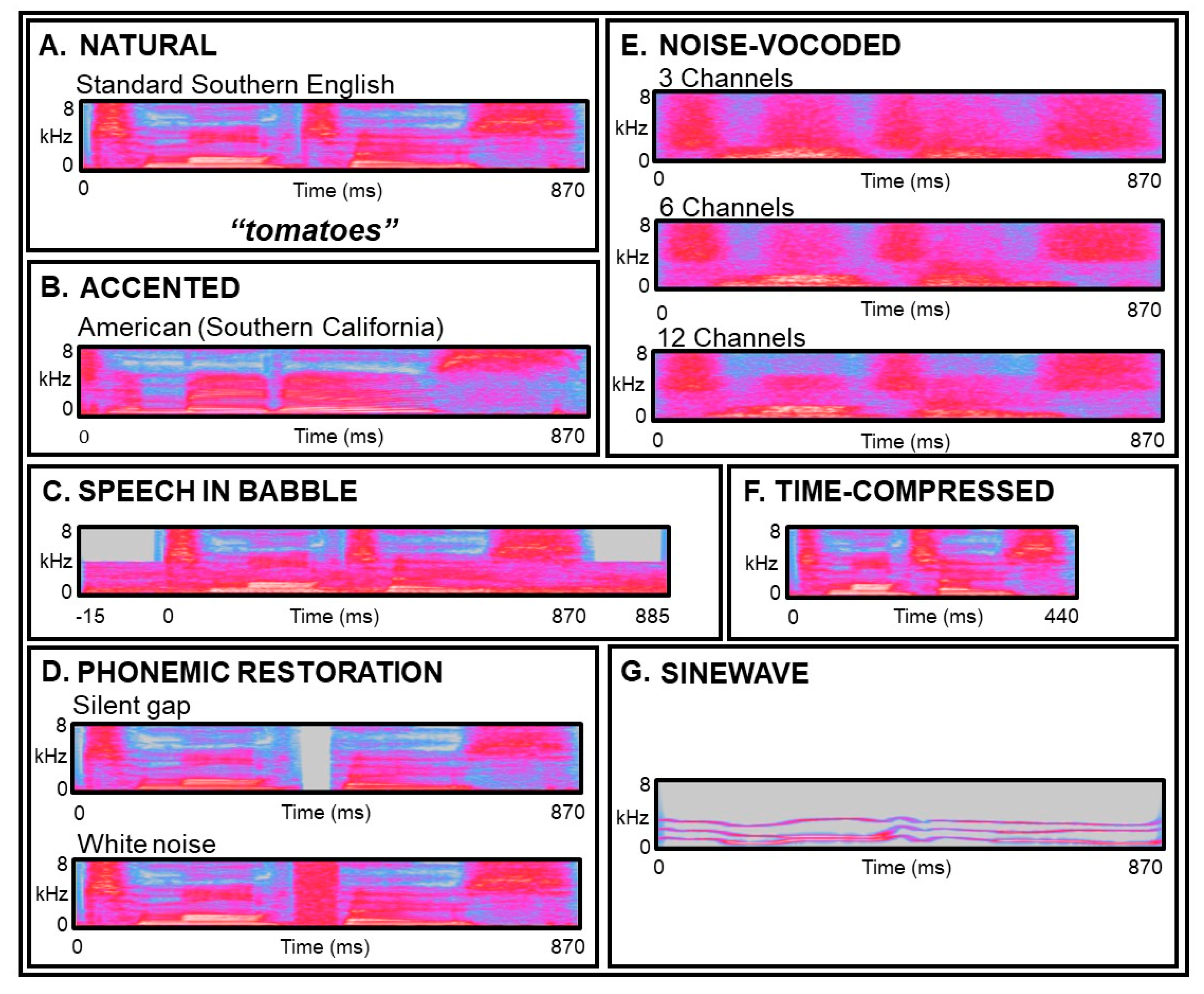{kind=link}
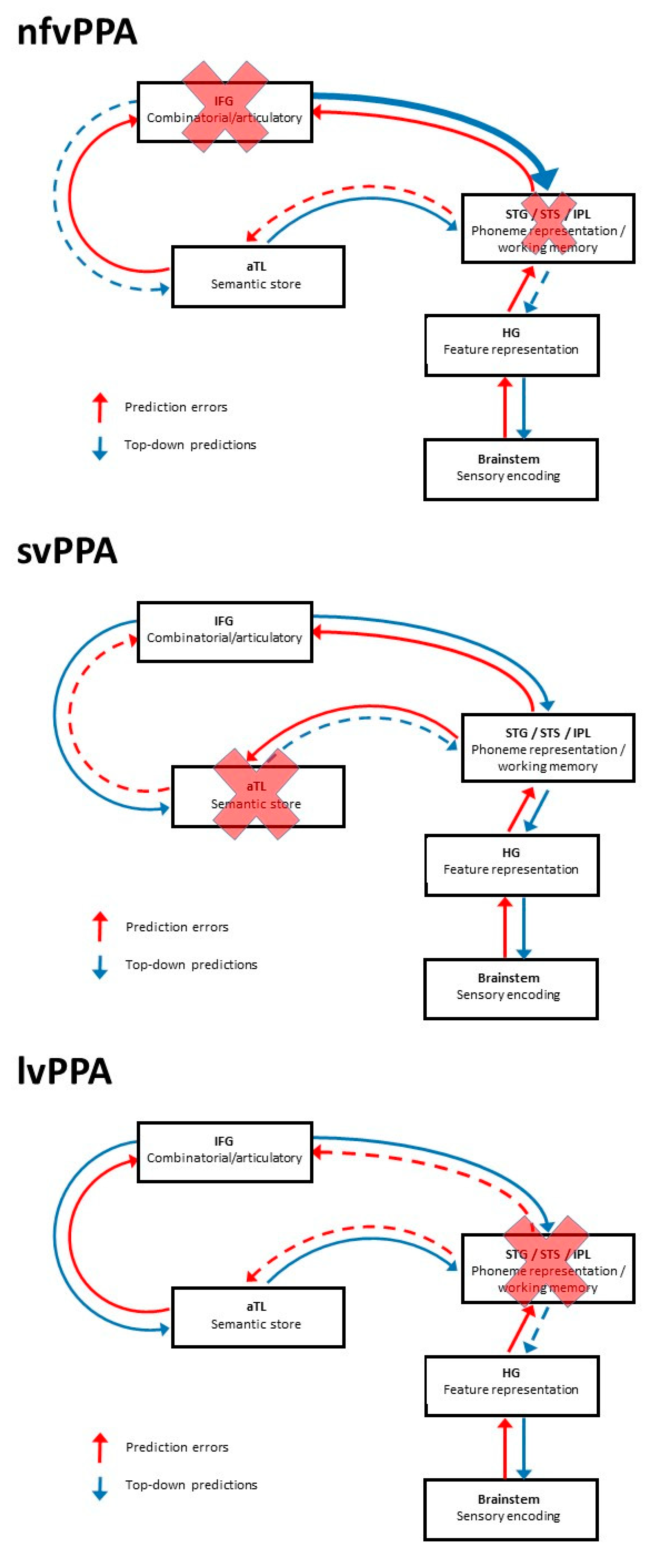{kind=link}
| Degradation Type | Study | Participants | Methodology | Major Findings |
|---|---|---|---|---|
| ACCENTS Target process: phonemic and intonational representations Ecological relevance: Understanding messages conveyed via non-canonical spoken phonemes and suprasegmental intonation | Bent and Bradlow [65] | 65 healthy participants (age: 19.1) | Participants listened to English sentences spoken by Chinese, Korean, and English native speakers. | Non-native listeners found speech from non-native English speakers as intelligible as from a native speaker. |
| Clarke and Garrett [66] | 164 healthy participants (American English) | Participants listened to English sentences spoken with a Spanish, Chinese, and English accent. | Processing speed initially slower for accented speech, but this deficit diminished with exposure. | |
| Floccia, Butler, Goslin and Ellis [54] | 54 healthy participants (age 19.7; Southern British English) | Participants had to say if the last word in a spoken sentence was real or not. | Changing accent caused a delay in word identification, whether accent change was regional or foreign. | |
| ALTERED AUDITORY FEEDBACK Target process: Influence of auditory feedback on speech production Ecological relevance: Ability to hear, process, and regulate speech from own production. | Siegel and Pick [67] | 20 healthy participants | Participants produced speech whilst hearing amplified feedback of their own voice. | Participants lowered their voices (displaying the sidetone amplification effect) in all conditions. |
| Jones and Munhall [68] | 18 healthy participants (age: 22.4; Canadian English) | Participants produced vowels with altered feedback of F0 shifted up or down. | Participants compensated for change in F0. | |
| Donath et al. [69] | 22 healthy participants (age: 23; German) | Participants said a nonsense word with feedback of their frequency randomly shifting downwards. | Participants adjusted their voice F0 after a set period of time due to processing the feedback first. | |
| Stuart et al. [70] | 17 healthy participants (age: 32.9; American English) | Participants spoke under DAF at 0, 25, 50, 200 ms at normal and fast rates of speech. | There were more dysfluencies at 200 ms, and more dysfluencies at the fast rate of speech. | |
| DICHOTIC LISTENING Target process: Auditory scene analysis (auditory attention) Ecological relevance: Processing of spoken information with competing verbal material | Moray [71] | Healthy participants, no other information given | Participants were told to focus on a message played to one ear, with a competing message in the other ear. | Participants did not recognize the content in the unattended message. |
| Lewis [72] | 12 healthy participants | Participants were told to attend to message presented in one ear, with a competing message in the other. | Participants could not recall the unattended message, but semantic similarity affected reaction times. | |
| Ding and Simon [73] | 10 healthy participants (age 19–25) | Under MEG, participants heard competing messages in each ear, and asked to attend to each in turn. | Auditory cortex tracked temporal modulations of both signals, but was stronger for the attended one. | |
| NOISE-VOCODED SPEECH Target process: Phonemic spectral detail Ecological relevance: Understanding whisper (similar quality to speech heard by cochlear implant users) | Shannon, Zeng, Kamath, Wygonski and Ekelid [59] | 8 healthy participants | Participants listened to and repeated simple sentences that had been noise-vocoded to different degrees. | Performance improved with number of channels; high speech recognition was achieved with only 3 channels. |
| Davis, Johnsrude, Hervais-Adelman, Taylor and McGettigan [58] | 12 healthy participants (age 18–25; British English) | Participants listened to and then transcribed 6-channel noise-vocoded sentences. | Participants showed rapid improvement over the course of 30-sentence exposure. | |
| Scott, Rosen, Lang and Wise [35] | 7 healthy participants (age 38) | Under PET, participants listened to spoken sentences that were noise-vocoded to various degrees. | Selective response to speech intelligibility in left anterior STS. | |
| PERCEPTUAL RESTORATION Target process: Message interpolation Ecological relevance: Understanding messages in intermittent or varying noise (e.g., a poor telephone line) | Warren [57] | 20 healthy participants | Participants identified where the gap was in sentences where a phoneme was replaced by silence/white noise. | Participants were more likely to mislocalize a missing phoneme that was replaced by noise. |
| Samuel [74] | 20 healthy participants (English) | Participants heard sentences in which white noise was either “Added” to or “Replaced” a phoneme. | Phonemic restoration was more common for longer words and certain phone classes. | |
| Leonard, Baud, Sjerps and Chang [43] | 5 healthy participants (age 38.6; English/Italian) | Subdural electrode arrays recorded while participants listened to words with noise-replaced phonemes. | Electrode responses were comparable to intact words vs. words with a phoneme replaced. | |
| SINEWAVE SPEECH Target process: Speech reconstruction and adaptation from very impoverished cues Ecological relevance: Synthetic model for impoverished speech signal and perceptual learning | Remez, Rubin, Pisoni and Carrell [63] | 54 control participants | Naïve listeners heard SWS replicas of spoken sentences and were later asked to transcribe the sentences. | Most listeners did not initially identify the SWS as speech, but were able to transcribe them when told this. |
| Barker and Cooke [64] | 12 control participants | Participants were asked to transcribe SWS or amplitude-comodulated SWS sentences. | Recognition for SWS ranged from 35–90%, and amplitude-comodulated SWS ranged from 50–95%. | |
| Möttönen, Calvert, Jääskeläinen, Matthews, Thesen, Tuomainen and Sams [37] | 21 control participants (18–36; English) | Participants underwent two fMRI scans: one before training on SWS, and one post-training. | Activity in left posterior STS was increased after SWS training. | |
| SPEECH-IN-NOISE Target process: Auditory scene analysis (parsing of phonemes from acoustic background) Ecological relevance: Understanding messages in background noise (e.g., “cocktail party effect”) | Pichora-Fuller et al. [75] | 24 participants in three groups (age 23.9; 70.4; 75.8; English) | Participants repeated the last word of sentences in 8-talker babble. Half had predictable endings. | Both groups of older listeners derived more benefit from context than younger listeners. |
| Parbery-Clark et al. [76] | 31 control participants (incl. 16 musicians; age: 23; English) | Participants were assessed via clinical measures of speech perception in noise. | Musicians outperformed the non-musicians on both QuickSIN and HINT. | |
| Anderson et al. [77] | 120 control participants (age 63.9) | Peripheral auditory function, cognitive ability, speech-in-noise, and life experience were examined. | Central processing and cognitive function predicted variance in speech-in-noise perception. | |
| TIME-COMPRESSED SPEECH Target process: Phoneme duration (rate of presentation) Ecological relevance: Understanding rapid speech | Dupoux and Green [60] | 160 control participants (English) | Participants transcribed spoken sentence were compressed to 38% and 45% of their original durations. | Participants improved over time. This happened more rapidly for the 45% compressed sentences. |
| Poldrack et al. [78] | 8 control participants (age: 20–29; English) | Participants listened to time-compressed speech. Brain responses were tracked using fMRI. | Activity in bilateral IFG and left STG increased with compression, until speech became incomprehensible. | |
| Peelle et al. [79] | 8 control participants (age: 22.6; English) | Participants listened to sentences manipulated for complexity and time-compression in an fMRI study. | Time-compressed sentences recruited AC and premotor cortex, regardless of complexity. |
| Population | Study, Degradation | Participants | Methodology | Major Findings |
|---|---|---|---|---|
| Traumatic brain injury | Gallun et al. [80]: Central auditory processing | 36 blast-exposed military veterans (age: 32.8); 29 controls (age: 32.1) | Participants went through a battery of standardised behavioural tests of central auditory function: temporal pattern perception, GIN, MLD, DDT, SSW, and QuickSIN. | While no participant performed poorly on all behavioural testing, performance was impaired in central auditory processing for the blast-exposed veterans in comparison to matched-controls. |
| Saunders et al. [81]: Central auditory processing | 99 military veterans (age: 34.1) | Participants went through self-reported measures as well as a battery of standardised behavioural measures: HINT, NA LiSN-S, ATTR, TCST, and SSW. | Participants in this study showed measurable performance deficits on speech-in-noise perception, binaural processing, temporal resolution, and speech segregation. | |
| Gallun et al. [82]: Central auditory processing | 30 blast-exposed military veterans, with a least one blast occurring 10 years prior to study (age: 37.3); 29 controls (age: 39.2) | Participants went through a battery of standardised behavioural tests of central auditory function: GIN, DDT, SSW, FPT, and MLD. | Replicating the findings from Gallun et al., 2012, this study found that the central auditory processing deficits persisted in individuals tested an average of more than 7 years after blast exposure. | |
| Papesh et al. [83]: Central auditory processing | 16 blast-exposed veterans (age 36.9); 13 veteran controls (age 38) with normal peripheral hearing | Participants competed self-reported measures and standardised tests of speech-in-noise perception, DDT, SSW, TCST, plus auditory event-related potential studies. | Impaired cortical sensory gating was primarily influenced by a diagnosis of TBI and reduced habituation by a diagnosis of post-traumatic stress disorder. Cortical sensory gating and habituation to acoustic startle strongly predicted degraded speech perception | |
| Stroke aphasia | Bamiou et al. [84]: Dichotic listening | 8 patients with insular strokes (age: 63); 8 control participants (age: 63) | Participants heard pairs of spoken digits presented simultaneously to each ear, and were asked to repeat all four digits. | Dichotic listening was abnormal in five of the eight stroke patients. |
| Dunton et al. [85]: Accents | 16 participants with aphasia (age: 59); 16 controls (age: 59; English) | Participants heard English sentences spoken with a familiar (South-East British England) or unfamiliar (Nigerian) accent. | Aphasia patients made more errors in comprehending sentences spoken in an unfamiliar accent vs. a familiar accent. | |
| Jacks and Haley [86]: AAF (MAF) | 10 aphasia patients (age: 53.1); 10 controls (age: 63.1; English) | Participants produced spoken sentences with no feedback, DAF, FAF or noise-masked auditory feedback (MAF). | Speech rate increased under MAF but decreased with DAF and FAF in most participants with aphasia. | |
| Parkinson’s disease | Liu et al. [87]: AAF (MAF and FAF) | 12 PD participants (ge: 62.3); 13 control participants (age: 68.7) | Participants sustained a vowel whilst receiving changes in feedback of loudness (±3/4 dB) or pitch (±100 cents). | All participants produced compensatory responses to AAF, but response sizes were larger in PD than controls. |
| Chen et al. [88]: AAF (FAF) | 15 people with PD (age: 61); 15 control participants (age 61; Cantonese) | Participants were asked to vocalize a vowel sound with AAF pitch-shifted upwards or downwards. | PD participants produced larger magnitudes of compensation. | |
| Alzheimer’s disease | Gates et al. [89]: Dichotic digits | 17 ADs (age: 84); 64 MCI (age: 82.3); 232 controls (age: 78.8) | Participants listened to 40 numbers presented in pairs to each ear simultaneously. | AD patients scored the worst in the dichotic digits, followed by the MCI group and then the controls. |
| Golden et al. [90]: Auditory scene analysis | 13 AD participants (age: 66); 17 control participants (age: 68) | In fMRI, participants listened to their own name interleaved with or superimposed on multi-talker babble. | Significantly enhanced activation of right supramarginal gyrus in the AD vs. control group for the cocktail party effect. | |
| Ranasinghe et al. [91]: AAF (FAF) | 19 AD participants; 16 control participants | Participants were asked to produce a spoken vowel in context of AAF, with perturbations of pitch. | AD patients showed enhanced compensatory response and poorer pitch-response persistence vs. controls. | |
| Primary progressive aphasia | Hailstone et al. [92]: Accents | 20 ADs (age: 66.4); 6 nfvPPA (age: 66); 35 controls (age: 65); British English | Accent comprehension and accent recognition was assessed. VBM examined grey matter correlates. | Reduced comprehension for phrases in unfamiliar vs. familiar accents in AD and for words in nfvPPA; in AD group, grey matter associations of accent comprehension and recognition in anterior superior temporal lobe |
| Cope et al. [93]: Noise-vocoding | 11 nfvPPA (age: 72); 11 control participants (age: 72) | During MEG, participants listened to vocoded words presented with written text that matched/mismatched. | People with nfvPPA compared to controls showed delayed resolution of predictions in temporal lobe, enhanced frontal beta power and top-down fronto-temporal connectivity; precision of predictions correlated with beta power across groups | |
| Hardy et al. [94]: SWS | 9 nfvPPA (age: 69.6); 10 svPPA (age: 64.9); 7 lvPPA (age: 66.3); 17 control (age: 67.7) | Participants transcribed SWS of numbers/locations. VBM examined grey matter correlates in combined patient cohort. | Variable task performance groups; all showed spontaneous perceptual learning effects for SWS numbers; grey matter correlates in a distributed left hemisphere network extending beyond classical speech-processing cortices, perceptual learning effect in left inferior parietal cortex |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.; Benhamou, E.; Waters, S.; Johnson, J.C.S.; Volkmer, A.; Weil, R.S.; Marshall, C.R.; Warren, J.D.; Hardy, C.J.D. Processing of Degraded Speech in Brain Disorders. Brain Sci. 2021, 11, 394. https://doi.org/10.3390/brainsci11030394
Jiang J, Benhamou E, Waters S, Johnson JCS, Volkmer A, Weil RS, Marshall CR, Warren JD, Hardy CJD. Processing of Degraded Speech in Brain Disorders. Brain Sciences. 2021; 11(3):394. https://doi.org/10.3390/brainsci11030394
Chicago/Turabian StyleJiang, Jessica, Elia Benhamou, Sheena Waters, Jeremy C. S. Johnson, Anna Volkmer, Rimona S. Weil, Charles R. Marshall, Jason D. Warren, and Chris J. D. Hardy. 2021. "Processing of Degraded Speech in Brain Disorders" Brain Sciences 11, no. 3: 394. https://doi.org/10.3390/brainsci11030394
APA StyleJiang, J., Benhamou, E., Waters, S., Johnson, J. C. S., Volkmer, A., Weil, R. S., Marshall, C. R., Warren, J. D., & Hardy, C. J. D. (2021). Processing of Degraded Speech in Brain Disorders. Brain Sciences, 11(3), 394. https://doi.org/10.3390/brainsci11030394