
Auditory Beat Stimulation Modulates Memory-Related Single-Neuron Activity in the Human Medial Temporal Lobe

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.1.1. Participants
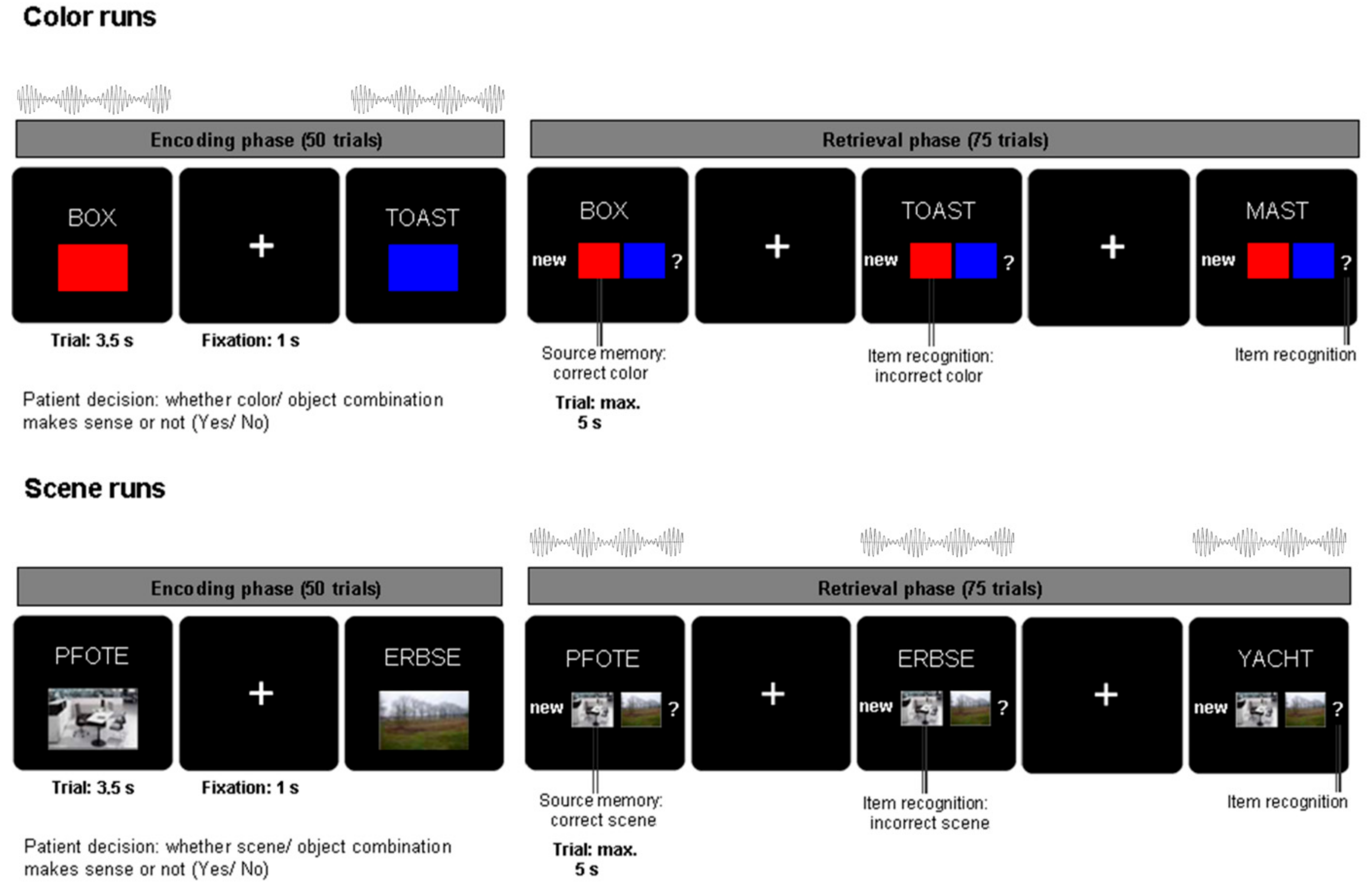
2.1.2. Experimental Paradigm
2.1.3. Data Recording and Preprocessing
2.2. Statistical Analysis
2.2.1. Evaluation of Firing Rates
2.2.2. Relationship between Beat Stimulation and Memory Performance: Behavioral Data
2.2.3. Relationship between Beat Stimulation and Memory Performance: Neural Data
2.2.4. Data and Code Availability
3. Results
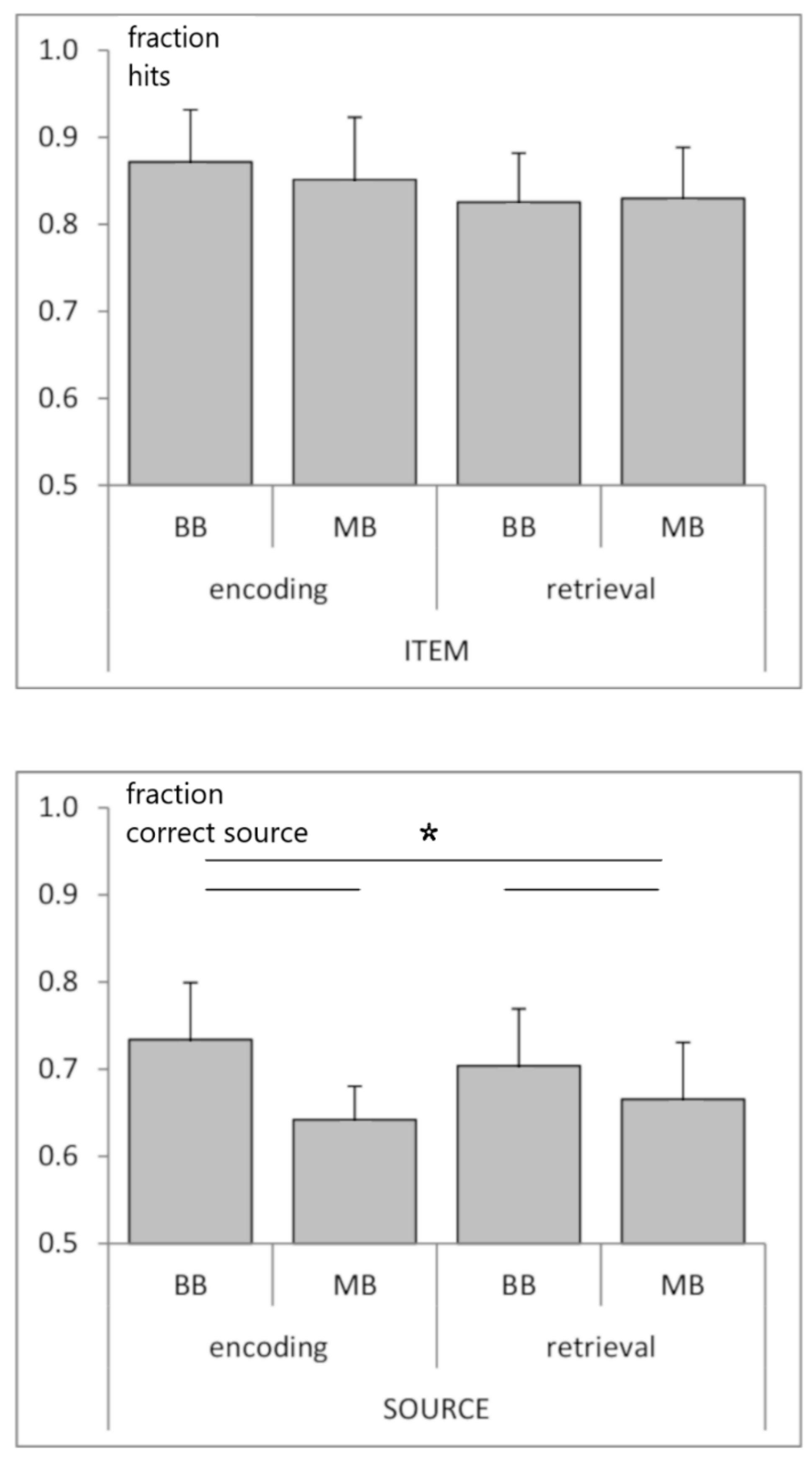
3.1. Behavioral Data
3.2. Relationship between Beat Stimulation and Memory Performance: Behavioral Data
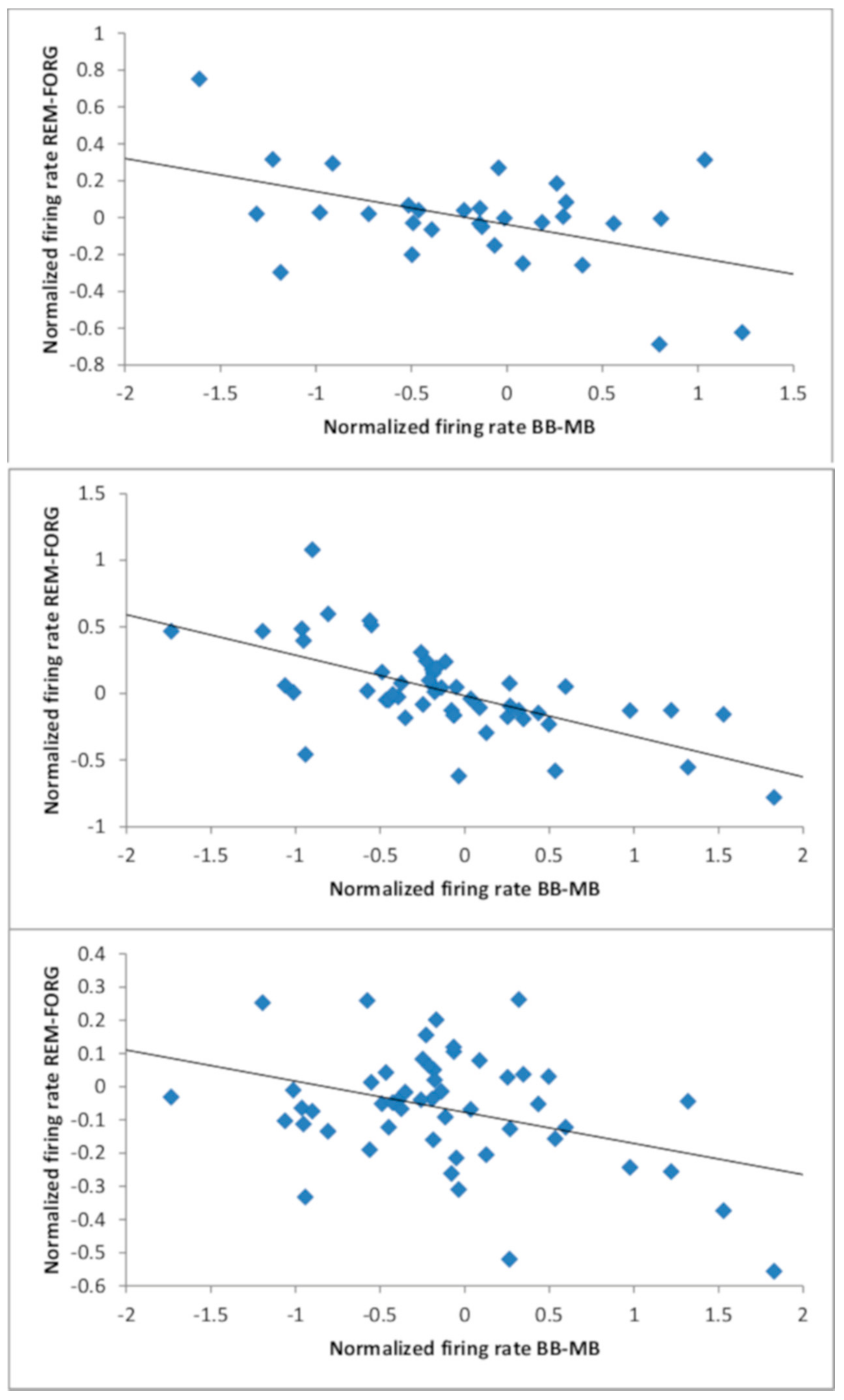
3.3. Relationship Between Beat Stimulation and Memory Performance: Neural Data
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chaieb, L.; Wilpert, E.C.; Reber, T.P.; Fell, J. Auditory Beat Stimulation and its Effects on Cognition and Mood States. Front. Psychiatry 2015, 6, 70. [Google Scholar] [CrossRef]
- Garcia-Argibay, M.; Santed, M.A.; Reales, J.M. Efficacy of binaural auditory beats in cognition, anxiety, and pain perception: A meta-analysis. Psychol. Res. 2019, 83, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Wernick, J.S.; Starr, A. Binaural interaction in the superior olivary complex of the cat: An analysis of field potentials evoked by binaural-beat stimuli. J. Neurophysiol. 1968, 31, 428–441. [Google Scholar] [CrossRef]
- Becher, A.K.; Höhne, M.; Axmacher, N.; Chaieb, L.; Elger, C.E.; Fell, J. Intracranial electroencephalogram power and phase synchronization changes during monaural and binaural beat stimulation. Eur. J. Neurosci. 2015, 41, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Derner, M.; Chaieb, L.; Surges, R.; Staresina, B.P.; Fell, J. Modulation of item and source memory by auditory beat stimulation: A pilot study with intracranial EEG. Front. Hum. Neurosci. 2018, 12, 500. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Taylor, P. Human auditory steady state responses to binaural and monaural beats. Clin. Neurophysiol. 2005, 116, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Fell, J.; Axmacher, N. The role of phase synchronization in memory processes. Nat. Rev. Neurosci. 2011, 12, 105–118. [Google Scholar] [CrossRef]
- Eichenbaum, H. A cortical–hippocampal system for declarative memory. Nat. Rev. Neurosci. 2000, 1, 41–50. [Google Scholar] [CrossRef]
- Derner, M.; Dehnen, G.; Chaieb, L.; Reber, T.P.; Borger, V.; Surges, R.; Staresina, B.P.; Mormann, F.; Fell, J. Patterns of sin-gle-neuron activity during associative recognition memory in the human medial temporal lobe. Neuroimage 2020, 221, 117214. [Google Scholar] [CrossRef]
- Bienenstock, E.L.; Cooper, L.N.; Munro, P.W. Theory for the development of neuron selectivity: Orientation specificity and binocular interaction in visual cortex. J. Neurosci. 1982, 2, 32–48. [Google Scholar] [CrossRef]
- Keck, T.; Hübener, M.; Bonhoeffer, T. Interactions between synaptic homeostatic mechanisms: An attempt to reconcile BCM theory, synaptic scaling, and changing excitation/inhibition balance. Curr. Opin. Neurobiol. 2017, 43, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Josse, G.; Tzourio-Mazoyer, N. Hemispheric specialization for language. Brain Res. Rev. 2004, 44, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Staresina, B.P.; Fell, J.; Do Lam, A.T.A.; Axmacher, N.; Henson, R.N. Memory signals are temporally dissociated in and across human hippocampus and perirhinal cortex. Nat. Neurosci. 2012, 15, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Niediek, J.; Boström, J.; Elger, C.E.; Mormann, F. Reliable Analysis of Single-Unit Recordings from the Human Brain under Noisy Conditions: Tracking Neurons over Hours. PLoS ONE 2016, 11, e0166598. [Google Scholar] [CrossRef] [PubMed]
- Mormann, F.; Dubois, J.; Kornblith, S.; Milosavljevic, M.; Cerf, M.; Ison, M.; Tsuchiya, N.; Kraskov, A.; Quiroga, R.Q.; Adolphs, R.; et al. A category-specific response to animals in the right human amygdala. Nat. Neurosci. 2011, 14, 1247–1249. [Google Scholar] [CrossRef] [PubMed]
- Rutishauser, U.; Schuman, E.M.; Mamelak, A.N. Online detection and sorting of extracellularly recorded action potentials in human medial temporal lobe recordings, in vivo. J. Neurosci. Methods 2006, 154, 204–224. [Google Scholar] [CrossRef]
- Staresina, B.P.; Reber, T.P.; Niediek, J.; Boström, J.; Elger, C.E.; Mormann, F. Recollection in the human hippocam-pal-entorhinal cell circuitry. Nat. Commun. 2019, 10, 1503. [Google Scholar] [CrossRef]
- Insausti, R.; Juottonen, K.; Soininen, H.; Insausti, A.M.; Partanen, K.; Vainio, P.; Laakso, M.P.; Pitkänen, A. MR volumetric analysis of the human entorhinal, perirhinal, and temporopolar cortices. Am. J. Neuroradiol. 1998, 19, 659–671. [Google Scholar]
- Pruessner, J.C.; Li, L.M.; Serles, W.; Pruessner, M.; Collins, D.L.; Kabani, N.; Lupien, S.; Evans, A.C. Volumetry of hippo-campus and amygdala with high-resolution MRI and three-dimensional analysis software: Minimizing the discrepancies between laboratories. Cereb. Cortex 2000, 10, 433–442. [Google Scholar] [CrossRef]
- Pruessner, J.C.; Köhler, S.; Crane, J.; Pruessner, M.; Lord, C.; Byrne, A.; Kabani, N.; Collins, D.L.; Evans, A.C. Volumetry of temporopolar, perirhinal, entorhinal and parahippocampal cortex from high-resolution MR images: Considering the variabil-ity of the collateral sulcus. Cereb. Cortex 2002, 12, 1342–1353. [Google Scholar] [CrossRef]
- Jirakittayakorn, N.; Wongsawat, Y. Brain responses to a 6-Hz binaural beat: Effects on general theta rhythm and frontal mid-line theta activity. Front. Neurosci. 2017, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Mormann, F.; Kornblith, S.; Quiroga, R.Q.; Kraskov, A.; Cerf, M.; Fried, I.; Koch, C. Latency and Selectivity of Single Neurons Indicate Hierarchical Processing in the Human Medial Temporal Lobe. J. Neurosci. 2008, 28, 8865–8872. [Google Scholar] [CrossRef]
- Kirkwood, A.; Rioult, M.G.; Bear, M.F. Experience-dependent modification of synaptic plasticity in visual cortex. Nature 1996, 381, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Bortoletto, M.; Pellicciari, M.C.; Rodella, C.; Miniussi, C. The Interaction With Task-induced Activity is More Important Than Polarization: A tDCS Study. Brain Stimul. 2015, 8, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Antal, A.; Begemeier, S.; Nitsch, M.A.; Paulus, W. Prior state of cortical activity influences subsequent practicing of a visuo-motor coordination task. Neuropsychologia 2008, 46, 3157–3161. [Google Scholar] [CrossRef]
- Lee, K.J.; Queenan, B.N.; Rozeboom, A.M.; Bellmore, R.; Lim, S.T.; Vicini, S.; Pak, D.T.S. Mossy fiber-CA3 synapses mediate homeostatic plasticity in mature hippocampal neurons. Neuron 2013, 77, 99–114. [Google Scholar] [CrossRef]
- Mendez, P.; Stefanelli, T.; Flores, C.E.; Muller, D.; Lüscher, C. Homeostatic plasticity in the hippocampus facilitates memory extinction. Cell. Rep. 2018, 22, 1451–1461. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Entorhinal | Parahippocampal | ||||
|---|---|---|---|---|---|
| Amygdala | Hippocampus | Cortex | Cortex | ||
| ENCODING | LEFT | 26% (1 × 10−4) | 38% (3 × 10−4) | 29% (8 × 10−8) | 16% (n.s.) |
| RIGHT | 22% (0.011) | 11% (n.s.) | 14% (n.s.) | 18% (n.s.) | |
| RETRIEVAL | LEFT | 58% (4 × 10−16) | 46% (2 × 10−5) | 35% (1 × 10−10) | 58% (3 × 10−10) |
| RIGHT | 28% (0.002) | 11% (n.s.) | 43% (3 × 10−5) | 59% (1 × 10−9) | |
| TIME WINDOW | |||||
|---|---|---|---|---|---|
| 0–500 | 500–1000 | 1000–1500 | 1500–2000 | ||
| ENCODING | item | n.s. | n.s. | n.s. | −0.47 (0.009) |
| source | n.s. | n.s. | n.s. | n.s. | |
| RETRIEVAL | item | n.s. | −0.49 (2 × 10−4) | −0.64 (4 × 10−7) | n.s. |
| source | n.s. | n.s. | −0.38 (0.006) | n.s. | |
| TIME WINDOW | |||||
|---|---|---|---|---|---|
| 0–500 | 500–1000 | 1000–1500 | 1500–2000 | ||
| ENCODING | item | n.s. | n.s. | n.s. | −0.47 (0.009) |
| source | n.s. | n.s. | −0.47 (0.008) | n.s. | |
| RETRIEVAL | item | −0.36 (0.010) | −0.48 (3 × 10−4) | −0.67 (4 × 10−8) | n.s. |
| source | n.s. | n.s. | n.s. | n.s. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derner, M.; Chaieb, L.; Dehnen, G.; Reber, T.P.; Borger, V.; Surges, R.; Staresina, B.P.; Mormann, F.; Fell, J. Auditory Beat Stimulation Modulates Memory-Related Single-Neuron Activity in the Human Medial Temporal Lobe. Brain Sci. 2021, 11, 364. https://doi.org/10.3390/brainsci11030364
Derner M, Chaieb L, Dehnen G, Reber TP, Borger V, Surges R, Staresina BP, Mormann F, Fell J. Auditory Beat Stimulation Modulates Memory-Related Single-Neuron Activity in the Human Medial Temporal Lobe. Brain Sciences. 2021; 11(3):364. https://doi.org/10.3390/brainsci11030364
Chicago/Turabian StyleDerner, Marlene, Leila Chaieb, Gert Dehnen, Thomas P. Reber, Valeri Borger, Rainer Surges, Bernhard P. Staresina, Florian Mormann, and Juergen Fell. 2021. "Auditory Beat Stimulation Modulates Memory-Related Single-Neuron Activity in the Human Medial Temporal Lobe" Brain Sciences 11, no. 3: 364. https://doi.org/10.3390/brainsci11030364
APA StyleDerner, M., Chaieb, L., Dehnen, G., Reber, T. P., Borger, V., Surges, R., Staresina, B. P., Mormann, F., & Fell, J. (2021). Auditory Beat Stimulation Modulates Memory-Related Single-Neuron Activity in the Human Medial Temporal Lobe. Brain Sciences, 11(3), 364. https://doi.org/10.3390/brainsci11030364