
Self-Positivity or Self-Negativity as a Function of the Medial Prefrontal Cortex



Abstract

1. Introduction
2. Methods
2.1. Participants
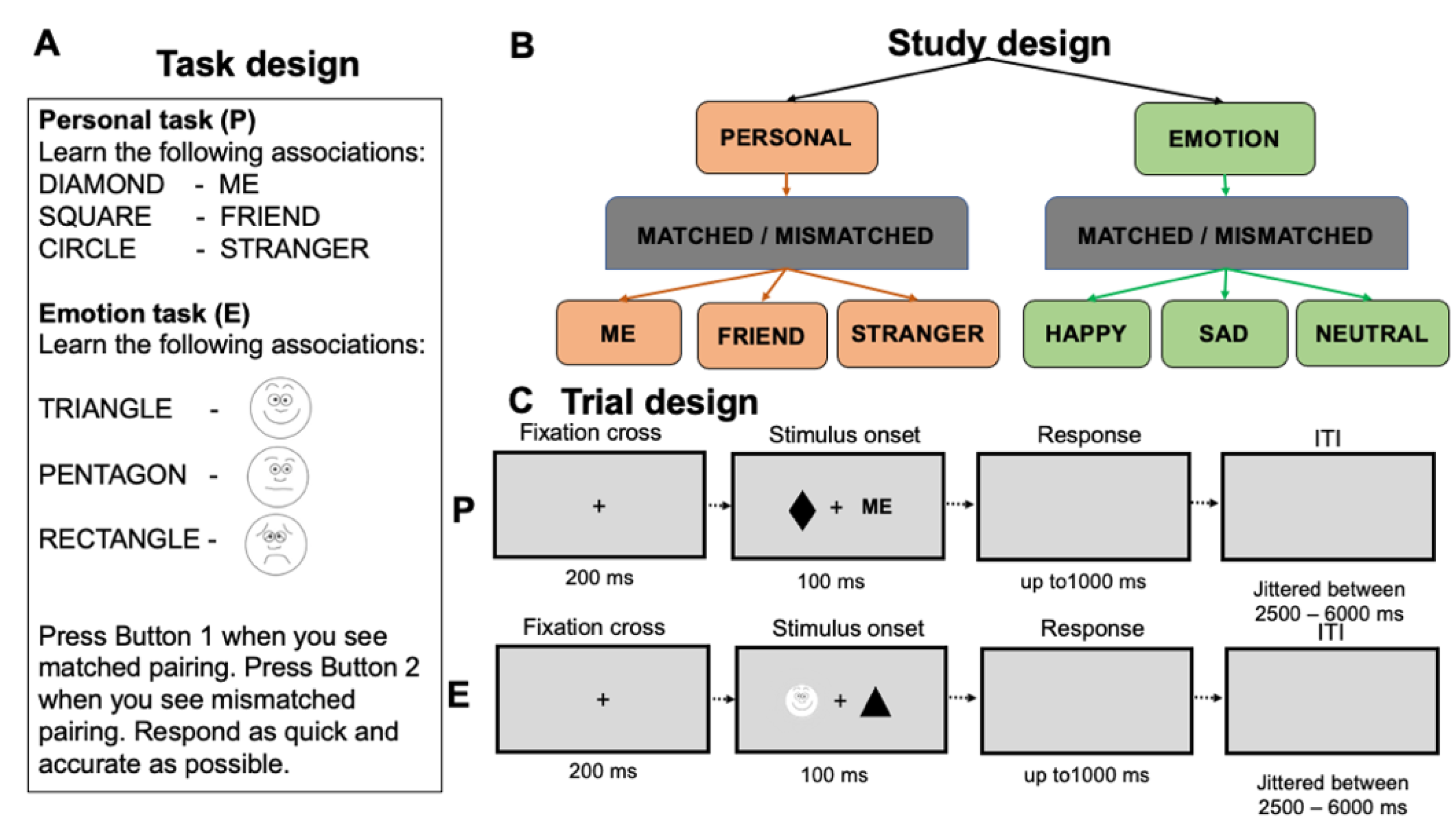
2.2. Task and Stimuli
2.3. fMRI Data Acquisition and Pre-Processing
2.4. Data Analysis
3. Results
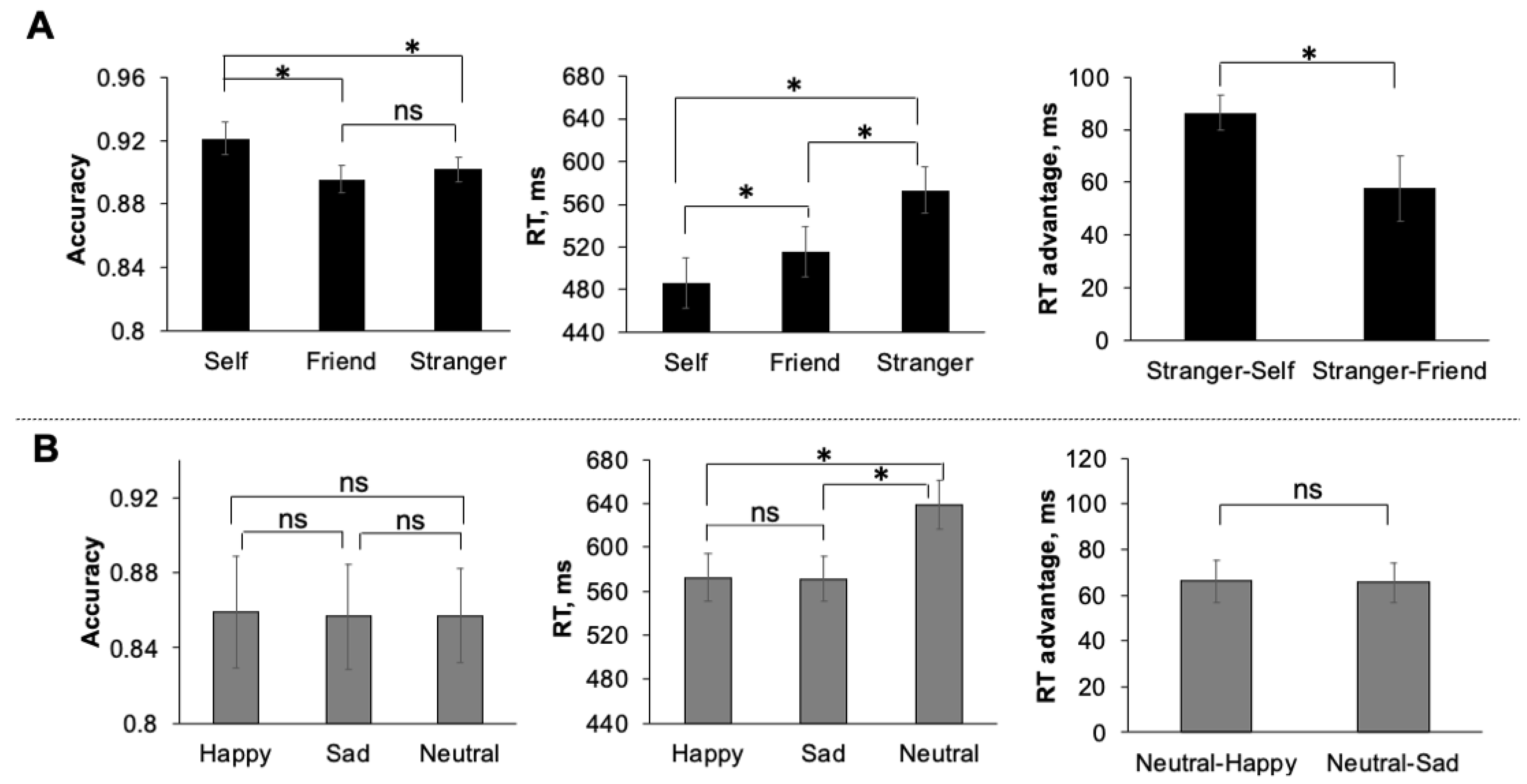
3.1. Behavioural Performance
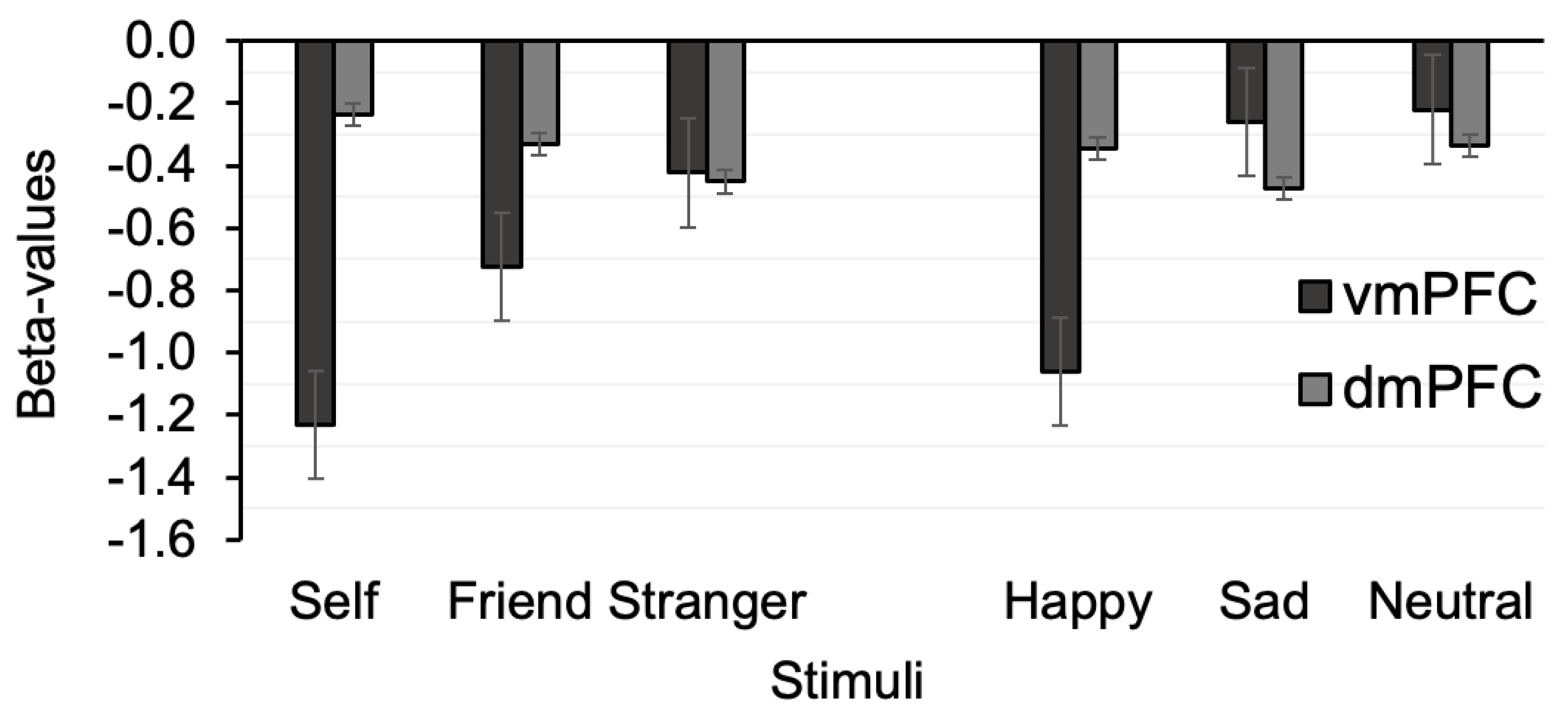
3.2. Univariate fMRI
3.3. The Effects of Personal and Emotion Associations in the MPFC
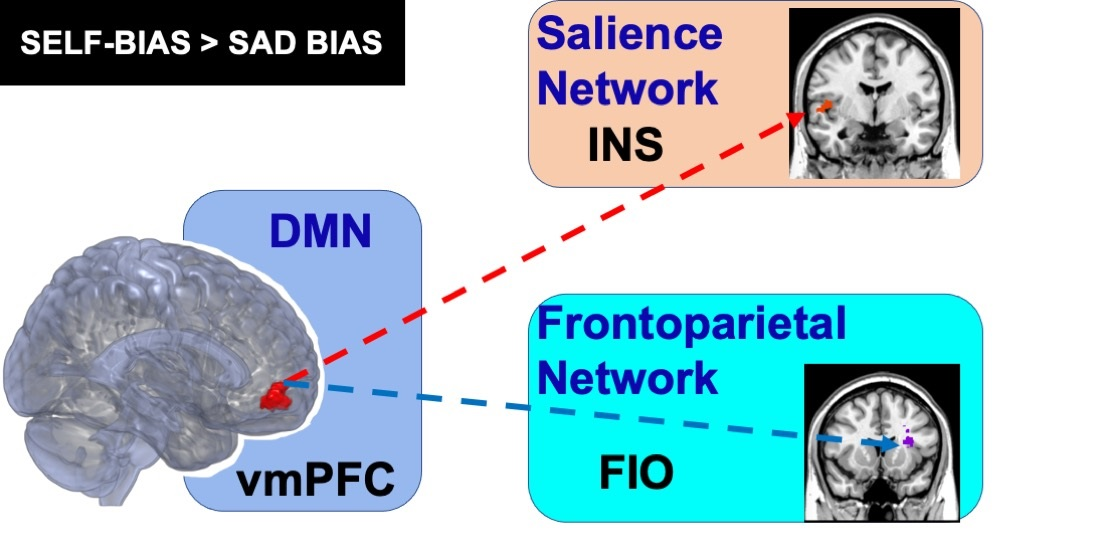
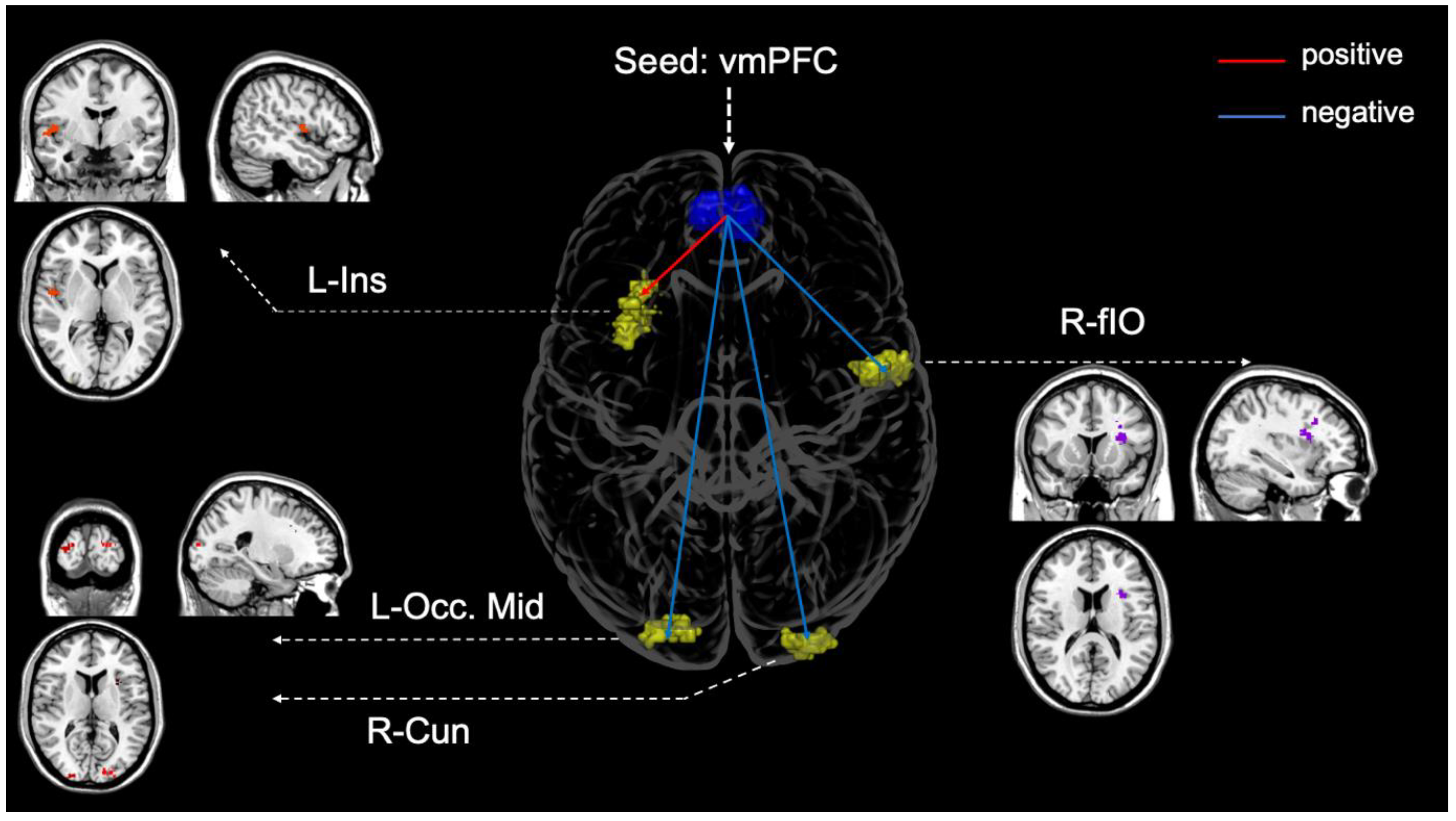
3.4. Seed-To Voxel Functional Connectivity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Feinberg, T.E.; Keenan, J.P. Where in the brain is the self? Conscious. Cogn. 2005, 14, 661–678. [Google Scholar] [CrossRef] [PubMed]
- Rottenberg, J. Emotions in Depression: What Do We Really Know? Annu. Rev. Clin. Psychol. 2017, 13, 241–263. [Google Scholar] [CrossRef]
- Northoff, G. Psychopathology and pathophysiology of the self in depression—Neuropsychiatric hypothesis. J. Affect. Disord. 2007, 104, 1–14. [Google Scholar] [CrossRef]
- Pick, S.; Goldstein, L.H.; Perez, D.L.; Nicholson, T.R. Emotional processing in functional neurological disorder: A review, biopsychosocial model and research agenda. J. Neurol. Neurosurg. Psychiatry 2019, 90, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q. The self in autism: An emerging view from neuroimaging. Neurocase 2011, 17, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Burkitt, I. The emotional self: Embodiment, reflexivity, and emotion regulation. Soc. Pers. Psychol. Compass 2018, 12, e12389. [Google Scholar] [CrossRef]
- Northoff, G. From Emotions to Consciousness—A Neuro-Phenomenal and Neuro-Relational Approach. Front. Psychol. 2012, 3, 303. [Google Scholar] [CrossRef] [PubMed]
- N’Diaye, K.; Sander, D.; Vuilleumier, P. Self-relevance processing in the human amygdala: Gaze direction, facial expression, and emotion intensity. Emotion 2009, 9, 798–806. [Google Scholar] [CrossRef]
- Lambie, J.A.; Marcel, A.J. Consciousness and the varieties of emotion experience: A theoretical framework. Psychol. Rev. 2002, 109, 219–259. [Google Scholar] [CrossRef]
- Herbert, B.M.; Pollatos, O.; Schandry, R. Interoceptive sensitivity and emotion processing: An EEG study. Int. J. Psychophysiol. 2007, 65, 214–227. [Google Scholar] [CrossRef]
- Emolnar-Szakacs, I.; Uddin, L.Q. Self-Processing and the Default Mode Network: Interactions with the Mirror Neuron System. Front. Hum. Neurosci. 2013, 7, 571. [Google Scholar] [CrossRef]
- Davey, C.G.; Pujol, J.; Harrison, B.J. Mapping the self in the brain’s default mode network. NeuroImage 2016, 132, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Gutchess, A.; Kensinger, E.A. Shared Mechanisms May Support Mnemonic Benefits from Self-Referencing and Emotion. Trends Cogn. Sci. 2018, 22, 712–724. [Google Scholar] [CrossRef] [PubMed]
- Fossati, P.; Hevenor, S.J.; Graham, S.J.; Grady, C.; Keightley, M.L.; Craik, F.; Mayberg, H. In Search of the Emotional Self: An fMRI Study Using Positive and Negative Emotional Words. Am. J. Psychiatry 2003, 160, 1938–1945. [Google Scholar] [CrossRef]
- Fossati, P.; Hevenor, S.J.; Lepage, M.; Graham, S.J.; Grady, C.; Keightley, M.L.; Craik, F.; Mayberg, H. Distributed self in episodic memory: Neural correlates of successful retrieval of self-encoded positive and negative personality traits. NeuroImage 2004, 22, 1596–1604. [Google Scholar] [CrossRef]
- Gusnard, D.A.; Akbudak, E.; Shulman, G.L.; Raichle, M.E. Medial prefrontal cortex and self-referential mental activity: Relation to a default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 4259–4264. [Google Scholar] [CrossRef]
- Jo, H.; Ou, Y.-Y.; Kung, C.-C. The neural substrate of self- and other-concerned wellbeing: An fMRI study. PLoS ONE 2019, 14, e0203974. [Google Scholar] [CrossRef] [PubMed]
- Etkin, A.; Egner, T.; Kalisch, R. Emotional processing in anterior cingulate and medial prefrontal cortex. Trends Cogn. Sci. 2011, 15, 85–93. [Google Scholar] [CrossRef]
- Lee, K.H.; Siegle, G.J. Common and distinct brain networks underlying explicit emotional evaluation: A meta-analytic study. Soc. Cogn. Affect. Neurosci. 2009, 7, 521–534. [Google Scholar] [CrossRef]
- Northoff, G.; Heinzel, A.; de Greck, M.; Bermpohl, F.; Dobrowolny, H.; Panksepp, J. Self-referential processing in our brain—A meta-analysis of imaging studies on the self. NeuroImage 2006, 31, 440–457. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, S.; Ueda, K.; Suzuki, S.-I.; Onoda, K.; Okamoto, Y.; Yamawaki, S. Self-referential processing of negative stimuli within the ventral anterior cingulate gyrus and right amygdala. Brain Cogn. 2009, 69, 218–225. [Google Scholar] [CrossRef]
- De Pisapia, N.; Barchiesi, G.; Jovicich, J.; Cattaneo, L. The role of medial prefrontal cortex in processing emotional self-referential information: A combined TMS/fMRI study. Brain Imaging Behav. 2018, 13, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Mezulis, A.H.; Abramson, L.Y.; Hyde, J.S.; Hankin, B.L. Is There a Universal Positivity Bias in Attributions? A Meta-Analytic Review of Individual, Developmental, and Cultural Differences in the Self-Serving Attributional Bias. Psychol. Bull. 2004, 130, 711–747. [Google Scholar] [CrossRef]
- Frewen, P.; Schroeter, M.L.; Riva, G.; Cipresso, P.; Fairfield, B.; Padulo, C.; Kemp, A.H.; Palaniyappan, L.; Owolabi, M.; Kusi-Mensah, K.; et al. Neuroimaging the consciousness of self: Review, and conceptual-methodological framework. Neurosci. Biobehav. Rev. 2020, 112, 164–212. [Google Scholar] [CrossRef]
- Beer, J.S.; Hughes, B.L. Neural systems of social comparison and the “above-average” effect. NeuroImage 2010, 49, 2671–2679. [Google Scholar] [CrossRef]
- Hughes, B.L.; Beer, J.S. Protecting the Self: The Effect of Social-evaluative Threat on Neural Representations of Self. J. Cogn. Neurosci. 2013, 25, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Fields, E.C.; Kuperberg, G.R. Dynamic Effects of Self-Relevance and Task on the Neural Processing of Emotional Words in Context. Front. Psychol. 2016, 6, 2003. [Google Scholar] [CrossRef] [PubMed]
- Fields, E.C.; Weber, K.; Stillerman, B.; Delaney-Busch, N.; Kuperberg, G.R. Functional MRI reveals evidence of a self-positivity bias in the medial prefrontal cortex during the comprehension of social vignettes. Soc. Cogn. Affect. Neurosci. 2019, 14, 613–621. [Google Scholar] [CrossRef]
- Sui, J.; He, X.; Humphreys, G.W. Perceptual effects of social salience: Evidence from self-prioritization effects on perceptual matching. J. Exp. Psychol. Hum. Percept. Perform. 2012, 38, 1105–1117. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; Humphreys, G.W. The Integrative Self: How Self-Reference Integrates Perception and Memory. Trends Cogn. Sci. 2015, 19, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Golubickis, M.; Falben, J.K.; Sahraie, A.; Visokomogilski, A.; Cunningham, W.A.; Sui, J.; Macrae, C.N. Self-prioritization and perceptual matching: The effects of temporal construal. Mem. Cogn. 2017, 45, 1223–1239. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-P.; Lan, Y.; Macrae, C.N.; Sui, J. Good Me Bad Me: Prioritisation of the Good-Self during Perceptual Decision-Making. Collabra Psychol. 2020, 6, 20. [Google Scholar] [CrossRef]
- Macrae, C.N.; Visokomogilski, A.; Golubickis, M.; Cunningham, W.A.; Sahraie, A. Self-relevance prioritizes access to visual awareness. J. Exp. Psychol. Hum. Percept. Perform. 2017, 43, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, M.; Knoblich, G. Self-prioritization of fully unfamiliar stimuli. Q. J. Exp. Psychol. 2019, 72, 2110–2120. [Google Scholar] [CrossRef]
- Reuther, J.; Chakravarthi, R. Does self-prioritization affect perceptual processes? Vis. Cogn. 2017, 25, 381–398. [Google Scholar] [CrossRef]
- Stolte, M.; Humphreys, G.W.; Yankouskaya, A.; Sui, J. Dissociating Biases towards the Self and Positive Emotion. Q. J. Exp. Psychol. 2017, 70, 1011–1022. [Google Scholar] [CrossRef]
- Sui, J.; Ohrling, E.; Humphreys, G.W. Negative mood disrupts self- and reward-biases in perceptual matching. Q. J. Exp. Psychol. 2016, 69, 1438–1448. [Google Scholar] [CrossRef] [PubMed]
- Sui, J. Self-Reference Acts as a Golden Thread in Binding. Trends Cogn. Sci. 2016, 20, 482–483. [Google Scholar] [CrossRef]
- Sui, J.; Gu, X. Self as Object: Emerging Trends in Self Research. Trends Neurosci. 2017, 40, 643–653. [Google Scholar] [CrossRef]
- Sui, J.; Rotshtein, P.; Humphreys, G.W. Coupling social attention to the self forms a network for personal significance. Proc. Natl. Acad. Sci. USA 2013, 110, 7607–7612. [Google Scholar] [CrossRef] [PubMed]
- Yankouskaya, A.; Humphreys, G.; Stolte, M.; Stokes, M.; Moradi, Z.; Sui, J. An anterior–posterior axis within the ventromedial prefrontal cortex separates self and reward. Soc. Cogn. Affect. Neurosci. 2017, 12, 1859–1868. [Google Scholar] [CrossRef]
- Watson, D.; Weber, K.; Assenheimer, J.S.; Clark, L.A.; Al, E. Testing a tripartite model: I. Evaluating the convergent and discriminant validity of anxiety and depression symptom scales. J. Abnorm. Psychol. 1995, 104, 3–14. [Google Scholar] [CrossRef]
- Wong, D.L.; Baker, C.M. Pain in children: Comparison of assessment scales. Pediatr. Nurs. 1988, 14, 9–17. [Google Scholar] [PubMed]
- Glaser, D.; Friston, K. Covariance Components; Elsevier: Amsterdam, The Netherlands, 2007; pp. 140–147. [Google Scholar]
- Ashburner, J. A fast diffeomorphic image registration algorithm. NeuroImage 2007, 38, 95–113. [Google Scholar] [CrossRef]
- Ashburner, J.; Friston, K.J. Unified segmentation. NeuroImage 2005, 26, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. Conn: A Functional Connectivity Toolbox for Correlated and Anticorrelated Brain Networks. Brain Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef]
- Behzadi, Y.; Restom, K.; Liau, J.; Liu, T.T. A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. NeuroImage 2007, 37, 90–101. [Google Scholar] [CrossRef]
- Friston, K.; Stephan, K.; Lund, T.; Morcom, A.; Kiebel, S. Mixed-effects and fMRI studies. NeuroImage 2005, 24, 244–252. [Google Scholar] [CrossRef]
- Pernet, C.R. Misconceptions in the use of the General Linear Model applied to functional MRI: A tutorial for junior neuro-imagers. Front. Neurosci. 2014, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.; Holmes, A.; Price, C.; Büchel, C.; Worsley, K. Multisubject fMRI Studies and Conjunction Analyses. NeuroImage 1999, 10, 385–396. [Google Scholar] [CrossRef]
- Heinzel, A.; Northoff, G. The Relationship of Self-Relatedness and Emotional Processing. J. Conscious. Stud. 2014, 21, 30–48. [Google Scholar]
- Chumbley, J.R.; Friston, K.J. False discovery rate revisited: FDR and topological inference using Gaussian random fields. NeuroImage 2009, 44, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.P.; Macrae, C.N.; Banaji, M.R. Dissociable Medial Prefrontal Contributions to Judgments of Similar and Dissimilar Others. Neuron 2006, 50, 655–663. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Ekoschack, J.; Epohlers, H.; Ehullemann, S.; Epaulus, W.; Ehappe, S. Effects of Frontal Transcranial Direct Current Stimulation on Emotional State and Processing in Healthy Humans. Front. Psychiatry 2012, 3, 58. [Google Scholar] [CrossRef]
- Nejati, V.; Majdi, R.; Salehinejad, M.A.; Nitsche, M.A. The role of dorsolateral and ventromedial prefrontal cortex in the processing of emotional dimensions. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- McIvor, L.; Sui, J.; Malhotra, T.; Drury, D.; Kumar, S. Self-referential processing and emotion context insensitivity in major depressive disorder. Eur. J. Neurosci. 2021, 53, 311–329. [Google Scholar] [CrossRef] [PubMed]
- Beer, J.S. Exaggerated Positivity in Self-Evaluation: A Social Neuroscience Approach to Reconciling the Role of Self-esteem Protection and Cognitive Bias. Soc. Pers. Psychol. Compass 2014, 8, 583–594. [Google Scholar] [CrossRef]
- Fan, W.; Zhong, Y.; Li, J.; Yang, Z.; Zhan, Y.; Cai, R.; Fu, X. Negative Emotion Weakens the Degree of Self-Reference Effect: Evidence from ERPs. Front. Psychol. 2016, 7, 1408. [Google Scholar] [CrossRef]
- D’Argembeau, A. On the Role of the Ventromedial Prefrontal Cortex in Self-Processing: The Valuation Hypothesis. Front. Hum. Neurosci. 2013, 7, 372. [Google Scholar] [CrossRef]
- Wild, B.; Erb, M.; Bartels, M.; Grodd, W. Why are smiles contagious? An fMRI study of the interaction between perception of facial affect and facial movements. Psychiatry Res. Neuroimaging 2003, 123, 17–36. [Google Scholar] [CrossRef]
- Todd, R.M.; Lee, W.; Evans, J.W.; Lewis, M.D.; Taylor, M.J. Withholding response in the face of a smile: Age-related differences in prefrontal sensitivity to Nogo cues following happy and angry faces. Dev. Cogn. Neurosci. 2012, 2, 340–350. [Google Scholar] [CrossRef][Green Version]
- Hiser, J.; Koenigs, M. The Multifaceted Role of the Ventromedial Prefrontal Cortex in Emotion, Decision Making, Social Cognition, and Psychopathology. Biol. Psychiatry 2018, 83, 638–647. [Google Scholar] [CrossRef]
- Scalabrini, A.; Vai, B.; Poletti, S.; Damiani, S.; Mucci, C.; Colombo, C.; Zanardi, R.; Benedetti, F.; Northoff, G. All roads lead to the default-mode network—global source of DMN abnormalities in major depressive disorder. Neuropsychopharmacol. 2020, 45, 2058–2069. [Google Scholar] [CrossRef]
- Compare, A.; Brugnera, A.; Adorni, R.; Sakatani, K. Effects of Positive and Negative Mood Induction on the Prefrontal Cortex Activity Measured by Near Infrared Spectroscopy. Single Mol. Single Cell Seq. 2016, 923, 151–157. [Google Scholar] [CrossRef]
- Posse, S.; Ackley, E.; Mutihac, R.; Zhang, T.; Hummatov, R.; Akhtari, M.; Chohan, M.O.; Fisch, B.; Yonas, H. High-Speed Real-Time Resting-State fMRI Using Multi-Slab Echo-Volumar Imaging. Front. Hum. Neurosci. 2013, 7, 479. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q.; Nomi, J.S.; Hébert-Seropian, B.; Ghaziri, J.; Boucher, O. Structure and Function of the Human Insula. J. Clin. Neurophysiol. 2017, 34, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Northoff, G. How is our self related to midline regions and the default-mode network? NeuroImage 2011, 57, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Soch, J.; Deserno, L.; Assmann, A.; Barman, A.; Walter, H.; Richardson-Klavehn, A.; Schott, B.H. Inhibition of Information Flow to the Default Mode Network During Self-Reference Versus Reference to Others. Cereb. Cortex 2016, 27, 3930–3942. [Google Scholar] [CrossRef] [PubMed]
- Barrett, L.F. The theory of constructed emotion: An active inference account of interoception and categorization. Soc. Cogn. Affect. Neurosci. 2016, 12, 1–23. [Google Scholar] [CrossRef]
- Hoemann, K.; Devlin, M.; Barrett, L.F. Comment: Emotions Are Abstract, Conceptual Categories That Are Learned by a Predicting Brain. Emot. Rev. 2020, 12, 253–255. [Google Scholar] [CrossRef]
- Cohn, M.A.; Fredrickson, B.L.; Brown, S.L.; Mikels, J.A.; Conway, A.M. Happiness unpacked: Positive emotions increase life satisfaction by building resilience. Emotion 2009, 9, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Phan, K.; Taylor, S.F.; Welsh, R.C.; Ho, S.-H.; Britton, J.C.; Liberzon, I. Neural correlates of individual ratings of emotional salience: A trial-related fMRI study. NeuroImage 2004, 21, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Arias, J.A.; Williams, C.; Raghvani, R.; Aghajani, M.; Baez, S.; Belzung, C.; Booij, L.; Busatto, G.; Chiarella, J.; Fu, C.H.; et al. The neuroscience of sadness: A multidisciplinary synthesis and collaborative review. Neurosci. Biobehav. Rev. 2020, 111, 199–228. [Google Scholar] [CrossRef]
- Wager, T.D.; Kang, J.; Johnson, T.D.; Nichols, T.E.; Satpute, A.B.; Barrett, L.F. A Bayesian Model of Category-Specific Emotional Brain Responses. PLoS Comput. Biol. 2015, 11, e1004066. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Han, X.; Wu, B.; Shi, Z.; Marks, M.; Wang, S.; Wu, X.; Han, S. Neural activation in response to the two sides of emotion. Neurosci. Lett. 2018, 684, 140–144. [Google Scholar] [CrossRef]
- Banich, M.T.; Floresco, S. Reward systems, cognition, and emotion: Introduction to the special issue. Cogn. Affect. Behav. Neurosci. 2019, 19, 409–414. [Google Scholar] [CrossRef]
- Enzi, B.; De Greck, M.; Prösch, U.; Tempelmann, C.; Northoff, G. Is Our Self Nothing but Reward? Neuronal Overlap and Distinction between Reward and Personal Relevance and Its Relation to Human Personality. PLoS ONE 2009, 4, e8429. [Google Scholar] [CrossRef]
- Northoff, G.; Hayes, D.J. Is Our Self Nothing but Reward? Biol. Psychiatry 2011, 69, 1019–1025. [Google Scholar] [CrossRef]
- Yankouskaya, A.; Bührle, R.; Lugt, E.; Stolte, M.; Sui, J. Intertwining personal and reward relevance: Evidence from the drift-diffusion model. Psychol. Res. 2018, 84, 32–50. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Contrast | Label | x | y | z | k | Z |
|---|---|---|---|---|---|---|
| happy > neutral | Precentral_L | −26 | −22 | 64 | 289 | 3.67 |
| Parietal_Inf_l | −40 | −38 | 40 | 543 | 4.65 | |
| sad > neutral | Frontal_Inf_Tri_L | −54 | 14 | 28 | 421 | 4.49 |
| Supp Motor Area_L | −2 | 2 | 62 | 141 | 4.31 | |
| Frontal Sup_2_L | −24 | 0 | 64 | 306 | 4.09 | |
| Frontal_Inf_Tri_R | 46 | 16 | 22 | 207 | 3.93 |
| x | y | z | k | t-Value | |
|---|---|---|---|---|---|
| Left Insula * | −28 | 16 | 16 | 187 | 9.31 |
| Right frontal inferior opercular * | 46 | −04 | 04 | 103 | −5.72 |
| Right cuneus * | 24 | −96 | 10 | 61 | −5.51 |
| Left occipital middle gyrus | −14 | −90 | 14 | 48 | −5.44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yankouskaya, A.; Sui, J. Self-Positivity or Self-Negativity as a Function of the Medial Prefrontal Cortex. Brain Sci. 2021, 11, 264. https://doi.org/10.3390/brainsci11020264
Yankouskaya A, Sui J. Self-Positivity or Self-Negativity as a Function of the Medial Prefrontal Cortex. Brain Sciences. 2021; 11(2):264. https://doi.org/10.3390/brainsci11020264
Chicago/Turabian StyleYankouskaya, Alla, and Jie Sui. 2021. "Self-Positivity or Self-Negativity as a Function of the Medial Prefrontal Cortex" Brain Sciences 11, no. 2: 264. https://doi.org/10.3390/brainsci11020264
APA StyleYankouskaya, A., & Sui, J. (2021). Self-Positivity or Self-Negativity as a Function of the Medial Prefrontal Cortex. Brain Sciences, 11(2), 264. https://doi.org/10.3390/brainsci11020264