Multimodal Sensorimotor Integration of Visual and Kinaesthetic Afferents Modulates Motor Circuits in Humans



Abstract

1. Introduction
2. Materials and Methods
2.1. Subjects
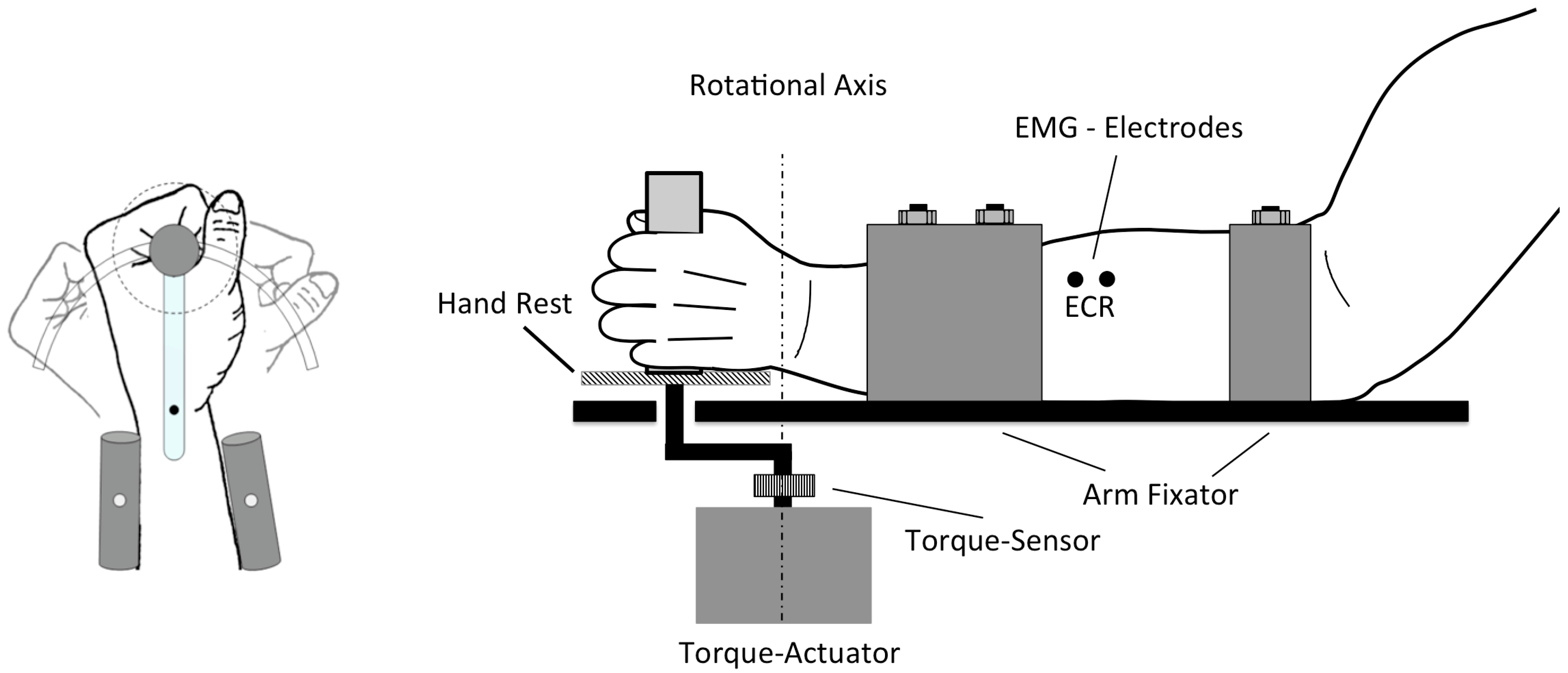
2.2. Protocol
2.3. Brain Stimulation
2.4. Electromyography (EMG)
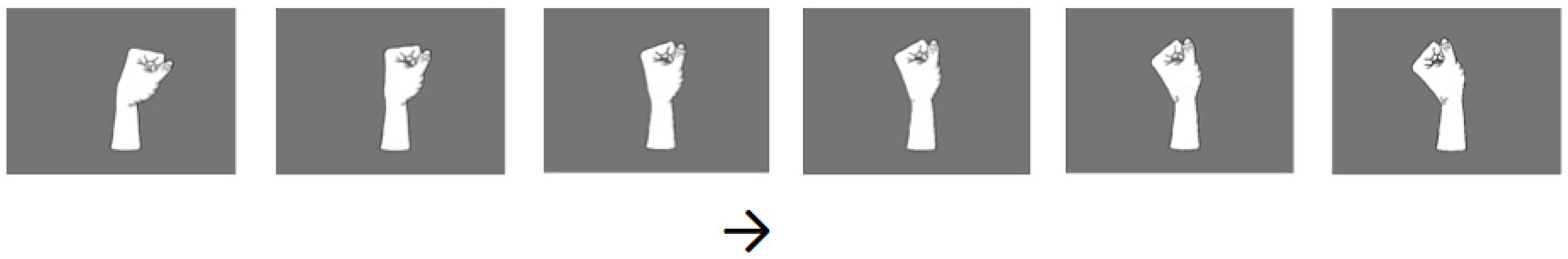
2.5. Visual Stimuli
2.6. Kinaesthetic Stimuli
2.7. Measurement and Data Processing
2.8. Statistics
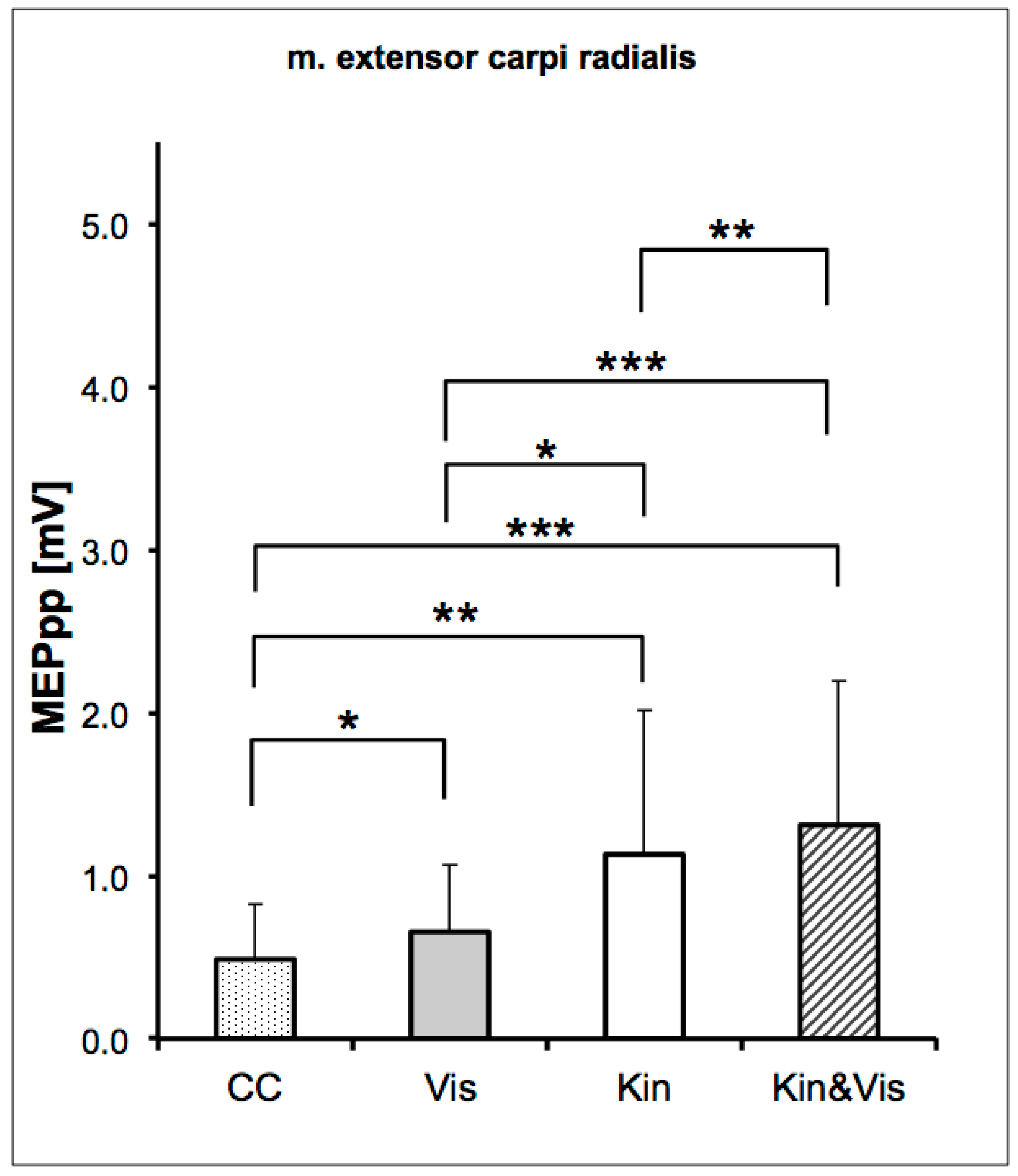
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rock, I.; Victor, J. Vision and touch: An experimentally created conflict between the two senses. Science 1964, 143, 594–596. [Google Scholar] [CrossRef]
- McGurk, H.; MacDonald, J. Hearing lips and seeing voices. Nature 1976, 264, 746–748. [Google Scholar] [CrossRef]
- Shams, L.; Kamitani, Y.; Shimojo, S. Illusions: What you see is what you hear. Nature 2000, 408, 788. [Google Scholar] [CrossRef]
- Ernst, M.O.; Bülthoff, H.H. Merging the senses into a robust percept. Trends Cogn. Sci. 2004, 8, 162–169. [Google Scholar] [CrossRef]
- Van Beers, R.J.; Sittig, A.C.; van der Gon Denier, J.J. How humans combine simultaneous proprioceptive and visual position information. Exp. Brain Res. 1996, 111, 253–261. [Google Scholar] [CrossRef]
- Van der Kooij, K.; Brenner, E.; van Beers, R.J.; Schot, W.D.; Smeets, J.B.J. Alignment to natural and imposed mismatches between the senses. J. Neurophysiol. 2013, 109, 1890–1899. [Google Scholar] [CrossRef]
- Di Pellegrino, G.; Fadiga, L.; Fogassi, L.; Gallese, V.; Rizzolatti, G. Understanding motor events: A neurophysiological study. Exp. Brain Res. 1992, 91, 176–180. [Google Scholar] [CrossRef]
- Mukamel, R.; Ekstrom, A.D.; Kaplan, J.; Iacoboni, M.; Fried, I. Single-neuron responses in humans during execution and observation of actions. Curr. Biol. 2010, 20, 750–756. [Google Scholar] [CrossRef]
- Gallese, V.; Fadiga, L.; Fogassi, L.; Rizzolatti, G. Action recognition in the premotor cortex. Brain 1996, 119, 593–609. [Google Scholar] [CrossRef]
- Umilta, M.A.; Kohler, E.; Gallese, V.; Fogassi, L.; Fadiga, L.; Keysers, C.; Rizzolatti, G. I know what you are doing: A neurophysiological study. Neuron 2001, 31, 155–165. [Google Scholar] [CrossRef]
- Dushanova, J.; Donoghue, J. Neurons in primary motor cortex engaged during action observation. Eur. J. Neurosci. 2010, 31, 386–398. [Google Scholar] [CrossRef]
- Fadiga, L.; Buccino, G.; Craighero, L.; Fogassi, L.; Gallese, V.; Pavesi, G. Corticospinal excitability is specifically modulated by motor imagery: A magnetic stimulation study. Neuropsychologia 1999, 37, 147–158. [Google Scholar] [CrossRef]
- Strafella, A.P.; Paus, T. Modulation of cortical excitability during action observation: A transcranial magnetic stimulation study. Neuroreport 2000, 11, 2289–2292. [Google Scholar] [CrossRef]
- Alaerts, K.; Senot, P.; Swinnen, S.P.; Craighero, L.; Wenderoth, N.; Fadiga, L. Force requirements of observed object lifting are encoded by the observer’s motor system: A TMS study. Eur. J. Neurosci. 2010, 31, 1144–1153. [Google Scholar] [CrossRef]
- Behrendt, F.; de Lussanet, M.H.E.; Zentgraf, K.; Zschorlich, V.R. Motor-evoked potentials in the lower back are modulated by visual perception of lifted weight. PLoS ONE 2016, 11, e0157811. [Google Scholar] [CrossRef]
- Buccino, G.; Binkofski, F.; Fink, G.R.; Fadiga, L.; Fogassi, L.; Gallese, V.; Seitz, R.J.; Zilles, K.; Rizzolatti, G.; Freund, H.J. Action observation activates premotor and parietal areas in a somatotopic manner: An fMRI study. Eur. J. Neurosci. 2001, 13, 400–404. [Google Scholar] [CrossRef]
- Saygin, A.P.; Wilson, S.M.; Hagler, D.J.; Bates, E.; Sereno, M.I. Point-light biological motion perception activates human premotor cortex. J. Neurosci. 2004, 24, 6181–6188. [Google Scholar] [CrossRef]
- de Lussanet, M.H.E.; Fadiga, L.; Michels, L.; Seitz, R.J.; Kleiser, R.; Lappe, M. Interaction of visual hemifield and body view in biological motion perception. Eur. J. Neurosci. 2008, 27, 514–522. [Google Scholar] [CrossRef]
- Michels, L.; Kleiser, R.; de Lussanet, M.H.E.; Seitz, R.J.; Lappe, M. Brain activity for peripheral biological motion in the posterior superior temporal gyrus and the fusiform gyrus: Dependence on visual hemifield and view orientation. Neuroimage 2009, 45, 151–159. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Sinigaglia, C. The functional role of the parieto-frontal mirror circuit: Interpretations and misinterpretations. Nat. Rev. Neurosci. 2010, 11, 264–274. [Google Scholar] [CrossRef]
- Jeannerod, M. Neural simulation of action: A unifying mechanism for motor cognition. Neuroimage 2001, 14, S103–S109. [Google Scholar] [CrossRef]
- Proske, U.; Gandevia, S.C. The kinaesthetic senses. J. Physiol. 2009, 587, 4139–4146. [Google Scholar] [CrossRef]
- Qi, F.; Nitsche, M.A.; Zschorlich, V.R. Interaction between transcranial random noise stimulation and observation-execution matching activity promotes motor cortex excitability. Front. Neurosci. 2019, 13, 69. [Google Scholar] [CrossRef]
- Heuninckx, S.; Wenderoth, N.; Swinnen, S.P. Systems neuroplasticity in the aging brain: Recruiting additional neural resources for successful motor performance in elderly persons. J. Neurosci. 2008, 28, 91–99. [Google Scholar] [CrossRef]
- Carp, J.; Park, J.; Hebrank, A.; Park, D.C.; Polk, T.A. Age-related neural dedifferentiation in the motor system. PLoS ONE 2011, 6, e29411. [Google Scholar] [CrossRef]
- Bernard, J.A.; Seidler, R.D. Evidence for motor cortex dedifferentiation in older adults. Neurobiol. Aging 2012, 33, 1890–1899. [Google Scholar] [CrossRef]
- Reuter, E.-M.; Behrens, M.; Zschorlich, V.R. Age-related differences in corticomotor facilitation indicate dedifferentiation in motor planning. Exp. Gerontol. 2015, 65, 79–84. [Google Scholar] [CrossRef]
- Flament, D.; Hall, E.; Lemon, R. The development of cortico-motoneuronal projections investigated using magnetic brain stimulation in the infant macaque. J. Physiol. 1992, 447, 755–768. [Google Scholar] [CrossRef]
- Olivier, E.; Edgley, S.A.; Armand, J.; Lemon, R.N. An electrophysiological study of the postnatal development of the corticospinal system in the macaque monkey. J. Neurosci. 1997, 17, 267–276. [Google Scholar] [CrossRef]
- Graziano, M.; Gandhi, S. Location of the polysensory zone in the precentral gyrus of anesthetized monkeys. Exp. Brain Res. 2000, 135, 259–266. [Google Scholar] [CrossRef]
- Bremmer, F.; Schlack, A.; Shah, N.J.; Zafiris, O.; Kubischik, M.; Hoffmann, K.-P.; Zilles, K.; Fink, G.R. Polymodal motion processing in posterior parietal and premotor cortex: A human fMRI study strongly implies equivalencies between humans and monkeys. Neuron 2001, 29, 287–296. [Google Scholar] [CrossRef]
- Maravita, A.; Iriki, A. Tools for the body (schema). Trends Cogn. Sci. 2004, 8, 79–86. [Google Scholar] [CrossRef]
- Makin, T.R.; Holmes, N.P.; Ehrsson, H.H. On the other hand: Dummy hands and peripersonal space. Behav. Brain Res. 2008, 191, 1–10. [Google Scholar] [CrossRef]
- Lee, C.R.; Yonk, A.J.; Wiskerke, J.; Paradiso, K.G.; Tepper, J.M.; Margolis, D.J. Opposing influence of sensory and motor cortical input on striatal circuitry and choice behavior. Curr. Biol. 2019, 29, 1313–1323. [Google Scholar] [CrossRef]
- Loporto, M.; Holmes, P.S.; Wright, D.J.; McAllister, C.J. Reflecting on mirror mechanisms: Motor resonance effects during action observation only present with low-intensity transcranial magnetic stimulation. PLoS ONE 2013, 8, e64911. [Google Scholar] [CrossRef]
- Zschorlich, V.R.; Köhling, R. How thoughts give rise to action-conscious motor intention increases the excitability of target-specific motor circuits. PLoS ONE 2013, 8, e83845. [Google Scholar] [CrossRef]
- Bruton, A.M.; Mellalieu, S.D.; Shearer, D.A. Observation as a method to enhance collective efficacy: An integrative review. Psychol. Sport Exerc. 2016, 24, 1–8. [Google Scholar] [CrossRef]
- Gueugneau, N.; Bove, M.; Ballay, Y.; Papaxanthis, C. Interhemispheric inhibition is dynamically regulated during action observation. Cortex 2016, 78, 138–149. [Google Scholar] [CrossRef]
- Schubert, M.; Curt, A.; Jensen, L.; Dietz, V. Corticospinal input in human gait: Modulation of magnetically evoked motor responses. Exp. Brain Res. 1997, 115, 234–246. [Google Scholar] [CrossRef]
- Holl, N.; Zschorlich, V.R. Neural control of joint stability during a ballistic force production task. Exp. Brain Res. 2011, 210, 229–242. [Google Scholar] [CrossRef]
- Zschorlich, V.R. Digital filtering of emg-signals. Electromyogr. Clin. Neurophysiol. 1989, 29, 81–86. [Google Scholar] [PubMed]
- Merletti, R.; Hermens, H. Detection and conditioning of the surface emg signal. Electromyography 2004, 107–131. [Google Scholar] [CrossRef]
- Fadiga, L.; Fogassi, L.; Pavesi, G.; Rizzolatti, G. Motor facilitation during action observation: A magnetic stimulation study. J. Neurophysiol. 1995, 73, 2608–2611. [Google Scholar] [CrossRef]
- Stinear, C.M.; Byblow, W.D. Motor imagery of phasic thumb abduction temporally and spatially modulates corticospinal excitability. Clin. Neurophysiol. 2003, 114, 909–914. [Google Scholar] [CrossRef]
- Kumru, H.; Albu, S.; Rothwell, J.; Leon, D.; Flores, C.; Opisso, E.; Tormos, J.M.; Valls-Sole, J. Modulation of motor cortex excitability by paired peripheral and transcranial magnetic stimulation. Clin. Neurophysiol. 2017, 128, 2043–2047. [Google Scholar] [CrossRef]
- Behrens, M.; Husmann, F.; Mau-Moeller, A.; Schlegel, J.; Reuter, E.M.; Zschorlich, V. Neuromuscular properties of the human wrist flexors as a function of the wrist joint angle. Front. Bioeng. Biotechnol. 2019, 7, 181. [Google Scholar]
- Longo, M.R.; Schüür, F.; Kammers, M.P.; Tsakiris, M.; Haggard, P. What is embodiment? A psychometric approach. Cognition 2008, 107, 978–998. [Google Scholar] [CrossRef]
- Stein, B.E.; Meredith, M.A. The Merging of the Senses; The MIT Press: Cambridge, MA, USA, 1993. [Google Scholar]
- Van Beers, R.J.; Sittig, A.C.; van der Gon, J.J.D. Integration of proprioceptive and visual position-information: An experimentally supported model. J. Neurophysiol. 1999, 81, 1355–1364. [Google Scholar] [CrossRef]
- Hay, J.C.; Pick, H.L.; Ikeda, K. Visual capture produced by prism spectacles. Psychon. Sci. 1965, 2, 215–216. [Google Scholar] [CrossRef]
- Shams, L.; Kim, R. Crossmodal influences on visual perception. Phys. Life Rev. 2010, 7, 269–284. [Google Scholar] [CrossRef]
- Guerraz, M.; Provost, S.; Narison, R.; Brugnon, A.; Virolle, S.; Bresciani, J.P. Integration of visual and proprioceptive afferents in kinesthesia. Neuroscience 2012, 223, 258–268. [Google Scholar] [CrossRef]
- Maravita, A.; Spence, C.; Driver, J. Multisensory integration and the body schema: Close to hand and within reach. Curr. Biol. 2003, 13, R531–R539. [Google Scholar] [CrossRef]
- Giroux, M.; Barra, J.; Zrelli, I.E.; Barraud, P.-A.; Cian, C.; Guerraz, M. The respective contributions of visual and proprioceptive afferents to the mirror illusion in virtual reality. PLoS ONE 2018, 13, e0203086. [Google Scholar] [CrossRef]
- Stinear, C.M.; Byblow, W.D.; Steyvers, M.; Levin, O.; Swinnen, S.P. Kinesthetic, but not visual, motor imagery modulates corticomotor excitability. Exp. Brain Res. 2006, 168, 157–164. [Google Scholar] [CrossRef]
- Alaerts, K.; Swinnen, S.P.; Wenderoth, N. Interaction of sound and sight during action perception: Evidence for shared modality-dependent action representations. Neuropsychologia 2009, 47, 2593–2599. [Google Scholar] [CrossRef]
- Strick, P.L.; Preston, J.B. Two representations of the hand in area 4 of a primate. I. Motor output organization. J. Neurophysiol. 1982, 48, 139–149. [Google Scholar] [CrossRef]
- Dum, R.P.; Strick, P.L. Motor areas in the frontal lobe: The anatomical substrate for the central control of movement. In Motor Cortex in Voluntary Movements: A Distributed System for Distributed Functions; CRC Press: Boca Raton, FL, USA, 2005; pp. 3–47. [Google Scholar]
- Ladda, A.M.; Wallwork, S.B.; Lotze, M. Multimodal sensory-spatial integration and retrieval of trained motor patterns for body coordination in musicians and dancers. Front. Psychol. 2020, 11, 3201. [Google Scholar] [CrossRef]
- Gruber, M.; Linnamo, V.; Strojnik, V.; Rantalainen, T.; Avela, J. Excitability at the motoneuron pool and motor cortex is specifically modulated in lengthening compared to isometric contractions. J. Neurophysiol. 2009, 101, 2030–2040. [Google Scholar] [CrossRef]
- Mouthon, A.; Ruffieux, J.; Wälchli, M.; Keller, M.; Taube, W. Task-dependent changes of corticospinal excitability during observation and motor imagery of balance tasks. Neuroscience 2015, 303, 535–543. [Google Scholar] [CrossRef]
- Duchateau, J.; Enoka, R.M. Neural control of lengthening contractions. J. Exp. Biol. 2016, 219, 197–204. [Google Scholar] [CrossRef]
- Calvert, G.; Spence, C.; Stein, B.E. The Handbook of Multisensory Processes; MIT Press: Cambridge, MA, USA, 2004. [Google Scholar]
- Keysers, C.; Gazzola, V. Expanding the mirror: Vicarious activity for actions, emotions, and sensations. Curr. Opin. Neurobiol. 2009, 19, 666–671. [Google Scholar] [CrossRef]
- Carrillo, M.; Han, Y.; Migliorati, F.; Liu, M.; Gazzola, V.; Keysers, C. Emotional mirror neurons in the rat’s anterior cingulate cortex. Curr. Biol. 2019, 29, 1301–1312. [Google Scholar] [CrossRef]
- Barraclough, N.E.; Xiao, D.; Baker, C.I.; Oram, M.W.; Perrett, D.I. Integration of visual and auditory information by superior temporal sulcus neurons responsive to the sight of actions. J. Cogn. Neurosci. 2005, 17, 377–391. [Google Scholar] [CrossRef]
- Dahl, C.D.; Logothetis, N.K.; Kayser, C. Spatial organization of multisensory responses in temporal association cortex. J. Neurosci. 2009, 29, 11924–11932. [Google Scholar] [CrossRef]
- Kaplan, J.T.; Iacoboni, M. Multimodal action representation in human left ventral premotor cortex. Cogn. Process. 2007, 8, 103–113. [Google Scholar] [CrossRef]
- Nakashita, S.; Saito, D.N.; Kochiyama, T.; Honda, M.; Tanabe, H.C.; Sadato, N. Tactile–visual integration in the posterior parietal cortex: A functional magnetic resonance imaging study. Brain Res. Bull. 2008, 75, 513–525. [Google Scholar] [CrossRef]
- Wallace, M.T.; Wilkinson, L.K.; Stein, B.E. Representation and integration of multiple sensory inputs in primate superior colliculus. J. Neurophysiol. 1996, 76, 1246–1266. [Google Scholar] [CrossRef]
- Murray, M.M.; Wallace, M.T. The Neural Bases of Multisensory Processes; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Calvert, G.A.; Campbell, R.; Brammer, M.J. Evidence from functional magnetic resonance imaging of crossmodal binding in the human heteromodal cortex. Curr. Biol. 2000, 10, 649–657. [Google Scholar] [CrossRef]
- Parker, P.R.; Brown, M.A.; Smear, M.C.; Niell, C.M. Movement-related signals in sensory areas: Roles in natural behavior. Trends Neurosci. 2020, 43, 581–595. [Google Scholar] [CrossRef]
- Meredith, M.A.; Stein, B.E. Visual, auditory, and somatosensory convergence on cells in superior colliculus results in multisensory integration. J. Neurophysiol. 1986, 56, 640–662. [Google Scholar] [CrossRef]
- Meredith, M.; Wallace, M.; Stein, B. Visual, auditory and somatosensory convergence in output neurons of the cat superior colliculus: Multisensory properties of the tecto-reticulo-spinal projection. Exp. Brain Res. 1992, 88, 181–186. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Sinigaglia, C. The mirror mechanism: A basic principle of brain function. Nat. Rev. Neurosci. 2016, 17, 757. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Cattaneo, L.; Fabbri-Destro, M.; Rozzi, S. Cortical mechanisms underlying the organization of goal-directed actions and mirror neuron-based action understanding. Physiol. Rev. 2014, 94, 655–706. [Google Scholar] [CrossRef]
- Fogassi, L.; Gallese, V.; Fadiga, L.; Luppino, G.; Matelli, M.; Rizzolatti, G. Coding of peripersonal space in inferior premotor cortex (area f4). J. Neurophysiol. 1996, 76, 141–157. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Matelli, M.; Pavesi, G. Deficits in attention and movement following the removal of postarcuate (area 6) and prearcuate (area 8) cortex in macaque monkeys. Brain 1983, 106, 655–673. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Control | Visual | KIN |
|---|---|---|---|
| VIS | 0.015687 | ||
| * | |||
| KIN | 0.001596 | 0.015454 | |
| ** | * | ||
| VIS & KIN | 0.000114 | 0.00081 | 0.004782 |
| *** | *** | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zschorlich, V.R.; Behrendt, F.; de Lussanet, M.H.E. Multimodal Sensorimotor Integration of Visual and Kinaesthetic Afferents Modulates Motor Circuits in Humans. Brain Sci. 2021, 11, 187. https://doi.org/10.3390/brainsci11020187
Zschorlich VR, Behrendt F, de Lussanet MHE. Multimodal Sensorimotor Integration of Visual and Kinaesthetic Afferents Modulates Motor Circuits in Humans. Brain Sciences. 2021; 11(2):187. https://doi.org/10.3390/brainsci11020187
Chicago/Turabian StyleZschorlich, Volker R., Frank Behrendt, and Marc H. E. de Lussanet. 2021. "Multimodal Sensorimotor Integration of Visual and Kinaesthetic Afferents Modulates Motor Circuits in Humans" Brain Sciences 11, no. 2: 187. https://doi.org/10.3390/brainsci11020187
APA StyleZschorlich, V. R., Behrendt, F., & de Lussanet, M. H. E. (2021). Multimodal Sensorimotor Integration of Visual and Kinaesthetic Afferents Modulates Motor Circuits in Humans. Brain Sciences, 11(2), 187. https://doi.org/10.3390/brainsci11020187