
Action Video Games Enhance Attentional Control and Phonological Decoding in Children with Developmental Dyslexia

 ,
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Reading Tasks
2.2.1. Phonological Decoding Tasks
2.2.2. Word Reading Tasks
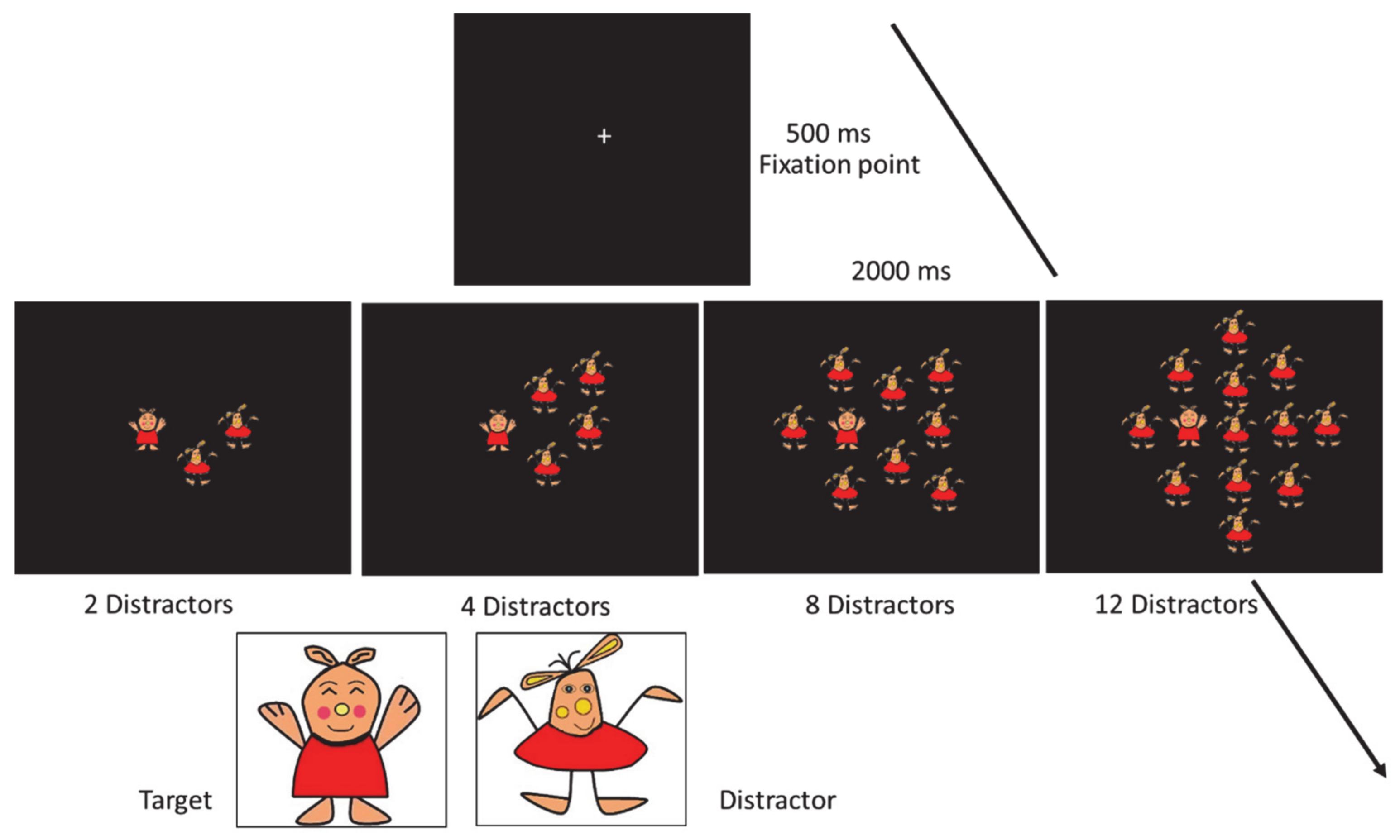
2.3. Visual Search Task
2.4. Non-Alphanumeric RAN Task
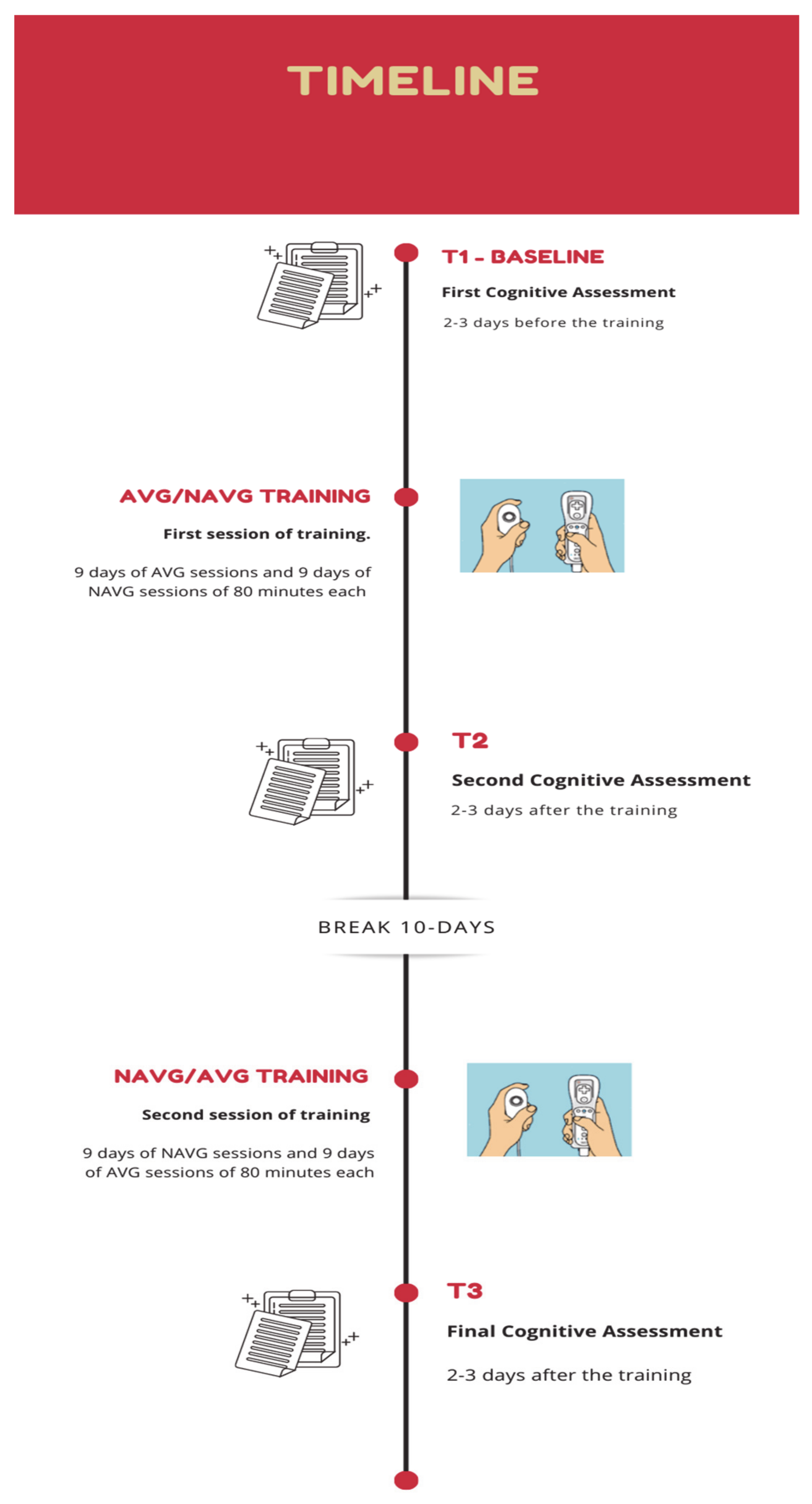
2.5. Training Procedure
3. Results
3.1. Within-Subject Analysis: Pre vs. Post AVG and Pre vs. Post NAVG
3.1.1. Reading Task
Phonological Decoding Tasks
Word Reading Tasks
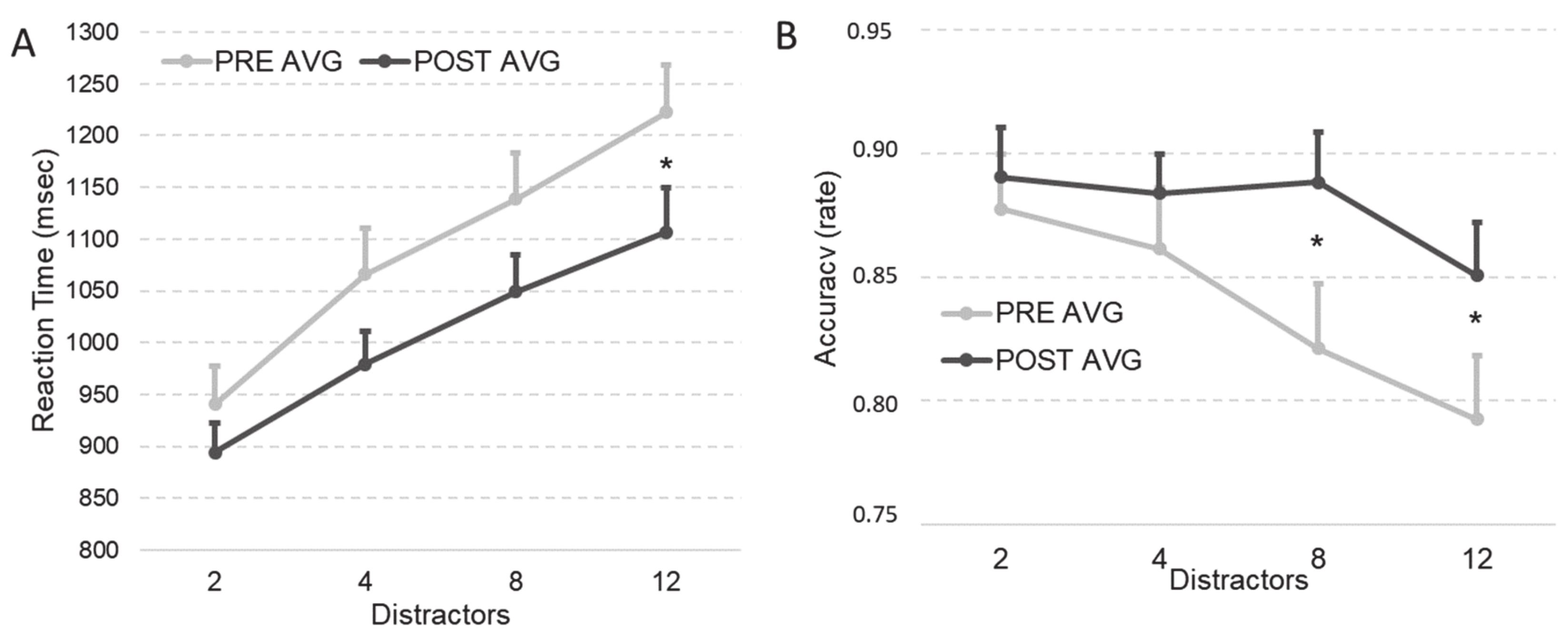
3.1.2. Visual Search Task
RTs
Accuracy
3.1.3. Non-Alphanumeric RAN Task
3.1.4. Action Video Game Ability after Training
3.2. Between-Subjects Analysis
3.2.1. T1 vs. T2
Phonological Decoding Tasks
Visual Search Task
3.2.2. T2 vs. T3
Phonological Decoding Tasks
Visual Search Task
3.2.3. Comparison between First NAVG Group vs. Second NAVG Group
Reading Task: Phonological Decoding Tasks
Visual Search Task
3.2.4. Comparison between Total AVG and First NAVG Groups
Reading Task: Phonological Decoding Tasks
Visual Search Task
Comparison between HSP and LSP Groups
Reading Task: Phonological Decoding Tasks
Visual Search Task
4. Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Share, D.L. On the Anglocentricities of current reading research and practice: The perils of overreliance on an "outlier" orthography. Psychol. Bull. 2008, 134, 584–615. [Google Scholar] [CrossRef] [PubMed]
- Hornickel, J.; Kraus, N. Unstable Representation of Sound: A Biological Marker of Dyslexia. J. Neurosci. 2013, 33, 3500–3504. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.L.; Pennington, B.F. Developmental Dyslexia. Annu. Rev. Clin. Psychol. 2015, 11, 283–307. [Google Scholar] [CrossRef] [PubMed]
- Vellutino, F.R.; Fletcher, J.M.; Snowling, M.J.; Scanlon, D.M. Specific reading disability (dyslexia): What have we learned in the past four decades? J. Child Psychol. Psychiatry 2004, 45, 2–40. [Google Scholar] [CrossRef] [PubMed]
- McArthur, G.; Eve, P.M.; Jones, K.; Banales, E.; Kohnen, S.; Anandakumar, T.; Larsen, L.; Marinus, E.; Wang, H.-C.; Castles, A. Phonics training for English-speaking poor readers. Cochrane Database Syst. Rev. 2012, 12, CD009115. [Google Scholar] [CrossRef]
- Peters, J.L.; De Losa, L.; Bavin, E.L.; Crewther, S.G. Efficacy of dynamic visuo-attentional interventions for reading in dyslexic and neurotypical children: A systematic review. Neurosci. Biobehav. Rev. 2019, 100, 58–76. [Google Scholar] [CrossRef]
- Menghini, D.; Finzi, A.; Benassi, M.; Bolzani, R.; Facoetti, A.; Giovagnoli, S.; Ruffino, M.; Vicari, S. Different underlying neurocognitive deficits in developmental dyslexia: A comparative study. Neuropsychologia 2010, 48, 863–872. [Google Scholar] [CrossRef]
- Bertoni, S.; Franceschini, S.; Ronconi, L.; Gori, S.; Facoetti, A. Is excessive visual crowding causally linked to developmental dyslexia? Neuropsychologia 2019, 130, 107–117. [Google Scholar] [CrossRef]
- Carroll, J.M.; Solity, J.; Shapiro, L.R. Predicting dyslexia using prereading skills: The role of sensorimotor and cognitive abilities. J. Child Psychol. Psychiatry 2015, 57, 750–758. [Google Scholar] [CrossRef]
- Franceschini, S.; Gori, S.; Ruffino, M.; Pedrolli, K.; Facoetti, A. A Causal Link between Visual Spatial Attention and Reading Acquisition. Curr. Biol. 2012, 22, 814–819. [Google Scholar] [CrossRef]
- Snowling, M.J.; Lervåg, A.; Nash, H.M.; Hulme, C. Longitudinal relationships between speech perception, phonological skills and reading in children at high-risk of dyslexia. Dev. Sci. 2018, 22, e12723. [Google Scholar] [CrossRef] [PubMed]
- Facoetti, A.; Paganoni, P.; Turatto, M.; Marzola, V.; Mascetti, G.G. Visual-Spatial Attention in Developmental Dyslexia. Cortex 2000, 36, 109–123. [Google Scholar] [CrossRef]
- Vidyasagar, T.R.; Pammer, K. Dyslexia: A deficit in visuo-spatial attention, not in phonological processing. Trends Cogn. Sci. 2010, 14, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Stein, J. Dyslexia: The Role of Vision and Visual Attention. Curr. Dev. Disord. Rep. 2014, 1, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Grainger, J.; Dufau, S.; Ziegler, J.C. A Vision of Reading. Trends Cogn. Sci. 2016, 20, 171–179. [Google Scholar] [CrossRef]
- Franceschini, S.; Bertoni, S.; Facoetti, A. Manual dexterity predicts phonological decoding speed in typical reading adults. Psychol. Res. 2021, 1–10. [Google Scholar] [CrossRef]
- Franceschini, S.; Bertoni, S.; Puccio, G.; Mancarella, M.; Gori, S.; Facoetti, A. Local perception impairs the lexical reading route. Psychol. Res. 2020, 1–9. [Google Scholar] [CrossRef]
- Eimer, M. The neural basis of attentional control in visual search. Trends Cogn. Sci. 2014, 18, 526–535. [Google Scholar] [CrossRef]
- McClelland, J.L.; Rumelhart, D.E. An interactive activation model of context effects in letter perception: I. An account of basic findings. Psychol. Rev. 1981, 88, 375–407. [Google Scholar] [CrossRef]
- Pelli, D.; Farell, B.; Moore, D.C. The remarkable inefficiency of word recognition. Nat. Cell Biol. 2003, 423, 752–756. [Google Scholar] [CrossRef]
- Perry, C.; Ziegler, J.C.; Zorzi, M. Nested incremental modeling in the development of computational theories: The CDP+ model of reading aloud. Psychol. Rev. 2007, 114, 273–315. [Google Scholar] [CrossRef] [PubMed]
- Bosse, M.-L.; Tainturier, M.J.; Valdois, S. Developmental dyslexia: The visual attention span deficit hypothesis. Cognition 2007, 104, 198–230. [Google Scholar] [CrossRef] [PubMed]
- Valdois, S.; Roulin, J.-L.; Bosse, M.L. Visual attention modulates reading acquisition. Vis. Res. 2019, 165, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Facoetti, A.; Trussardi, A.N.; Ruffino, M.; Lorusso, M.L.; Cattaneo, C.; Galli, R.; Molteni, M.; Zorzi, M. Multisensory Spatial Attention Deficits Are Predictive of Phonological Decoding Skills in Developmental Dyslexia. J. Cogn. Neurosci. 2010, 22, 1011–1025. [Google Scholar] [CrossRef] [PubMed]
- White, A.L.; Boynton, G.M.; Yeatman, J.D. The link between reading ability and visual spatial attention across development. Cortex 2019, 121, 44–59. [Google Scholar] [CrossRef]
- Gori, S.; Seitz, A.R.; Ronconi, L.; Franceschini, S.; Facoetti, A. Multiple Causal Links between Magnocellular–Dorsal Pathway Deficit and Developmental Dyslexia. Cereb. Cortex 2016, 26, 4356–4369. [Google Scholar] [CrossRef]
- Franceschini, S.; Bertoni, S.; Gianesini, T.; Gori, S.; Facoetti, A. A different vision of dyslexia: Local precedence on global perception. Sci. Rep. 2017, 7, 17462. [Google Scholar] [CrossRef]
- Facoetti, A.; Lorusso, M.L.; Paganoni, P.; Umiltà, C.; Mascetti, G.G. The role of visuospatial attention in developmental dyslexia: Evidence from a rehabilitation study. Cogn. Brain Res. 2003, 15, 154–164. [Google Scholar] [CrossRef]
- Dye, M.W.G.; Green, C.S.; Bavelier, D. Increasing Speed of Processing With Action Video Games. Curr. Dir. Psychol. Sci. 2009, 18, 321–326. [Google Scholar] [CrossRef]
- Chopin, A.; Bediou, B.; Bavelier, D. Altering perception: The case of action video gaming. Curr. Opin. Psychol. 2019, 29, 168–173. [Google Scholar] [CrossRef]
- Feng, J.; Spence, I.; Pratt, J. Playing an Action Video Game Reduces Gender Differences in Spatial Cognition. Psychol. Sci. 2007, 18, 850–855. [Google Scholar] [CrossRef] [PubMed]
- Mancarella, M.; Antzaka, A.; Bertoni, S.; Facoetti, A.; Lallier, M. Enhanced Disengagement of Auditory Attention and Phonological Skills in Action Video Gamers. Sci. Rep. 2021. Under Review. [Google Scholar]
- Nava, E.; Föcker, J.; Gori, M. Children can optimally integrate multisensory information after a short action-like mini game training. Dev. Sci. 2020, 23, e12840. [Google Scholar] [CrossRef] [PubMed]
- Oei, A.C.; Patterson, M.D. Enhancing Cognition with Video Games: A Multiple Game Training Study. PLoS ONE 2013, 8, e58546. [Google Scholar] [CrossRef]
- Green, C.S.; Bavelier, D. Action video game training for cognitive enhancement. Curr. Opin. Behav. Sci. 2015, 4, 103–108. [Google Scholar] [CrossRef]
- Franceschini, S.; Gori, S.; Ruffino, M.; Viola, S.; Molteni, M.; Facoetti, A. Action Video Games Make Dyslexic Children Read Better. Curr. Biol. 2013, 23, 462–466. [Google Scholar] [CrossRef]
- Franceschini, S.; Trevisan, P.; Ronconi, L.; Bertoni, S.; Colmar, S.; Double, K.; Facoetti, A.; Gori, S. Action video games improve reading abilities and visual-to-auditory attentional shifting in English-speaking children with dyslexia. Sci. Rep. 2017, 7, 5863. [Google Scholar] [CrossRef]
- Cancer, A.; Bonacina, S.; Antonietti, A.; Salandi, A.; Molteni, M.; Lorusso, M.L. The Effectiveness of Interventions for Developmental Dyslexia: Rhythmic Reading Training Compared With Hemisphere-Specific Stimulation and Action Video Games. Front. Psychol. 2020, 11, 11. [Google Scholar] [CrossRef]
- Łuniewska, M.; Chyl, K.; Dębska, A.; Kacprzak, A.; Plewko, J.; Szczerbiński, M.; Szewczyk, J.; Grabowska, A.; Jednoróg, K. Neither action nor phonological video games make dyslexic children read better. Sci. Rep. 2018, 8, 549. [Google Scholar] [CrossRef]
- Franceschini, S.; Bertoni, S.; Ronconi, L.; Molteni, M.; Gori, S.; Facoetti, A. “Shall We Play a Game?”: Improving Reading Through Action Video Games in Developmental Dyslexia. Curr. Dev. Disord. Rep. 2015, 2, 318–329. [Google Scholar] [CrossRef]
- Franceschini, S.; Bertoni, S. Improving action video games abilities increases the phonological decoding speed and phonological short-term memory in children with developmental dyslexia. Neuropsychologia 2019, 130, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Bavelier, D.; Green, C.S. Enhancing Attentional Control: Lessons from Action Video Games. Neuron 2019, 104, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Ma, W.; Gong, J.; He, H.; Dong, L.; Zhang, D.; Li, J.; Luo, C.; Yao, D. Action Video Game Experience Related to Altered Large-Scale White Matter Networks. Neural Plast. 2017, 2017, 7543686. [Google Scholar] [CrossRef]
- Etanaka, S.; Ikeda, H.; Kasahara, K.; Kato, R.; Tsubomi, H.; Sugawara, S.K.; Mori, M.; Hanakawa, T.; Sadato, N.; Honda, M.; et al. Larger Right Posterior Parietal Volume in Action Video Game Experts: A Behavioral and Voxel-Based Morphometry (VBM) Study. PLoS ONE 2013, 8, e66998. [Google Scholar] [CrossRef]
- Franceschini, S.; Mascheretti, S.; Bertoni, S.; Trezzi, V.; Andreola, C.; Gori, S.; Facoetti, A. Sluggish dorsally-driven inhibition of return during orthographic processing in adults with dyslexia. Brain Lang. 2018, 179, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gori, S.; Cecchini, P.; Bigoni, A.; Molteni, M.; Facoetti, A. Magnocellular-dorsal pathway and sub-lexical route in developmental dyslexia. Front. Hum. Neurosci. 2014, 8, 460. [Google Scholar] [CrossRef]
- Stein, J. The current status of the magnocellular theory of developmental dyslexia. Neuropsychologia 2019, 130, 66–77. [Google Scholar] [CrossRef]
- Lawton, T. Improving Dorsal Stream Function in Dyslexics by Training Figure/Ground Motion Discrimination Improves Attention, Reading Fluency, and Working Memory. Front. Hum. Neurosci. 2016, 10, 397. [Google Scholar] [CrossRef]
- Weissman, D.; Roberts, K.C.; Visscher, K.M.; Woldorff, M.G. The neural bases of momentary lapses in attention. Nat. Neurosci. 2006, 9, 971–978. [Google Scholar] [CrossRef]
- Ahissar, M.; Hochstein, S. The reverse hierarchy theory of visual perceptual learning. Trends Cogn. Sci. 2004, 8, 457–464. [Google Scholar] [CrossRef]
- Dehaene, S.; Cohen, L.; Morais, J.; Kolinsky, R. Illiterate to literate: Behavioural and cerebral changes induced by reading acquisition. Nat. Rev. Neurosci. 2015, 16, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Föcker, J.; Cole, D.; Beer, A.L.; Bavelier, D. Neural bases of enhanced attentional control: Lessons from action video game players. Brain Behav. 2018, 8, e01019. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Spence, I. Playing shooter and driving videogames improves top-down guidance in visual search. Atten. Percept. Psychophys. 2013, 75, 673–686. [Google Scholar] [CrossRef]
- Martinez-Trujillo, J.C.; Treue, S. Feature-Based Attention Increases the Selectivity of Population Responses in Primate Visual Cortex. Curr. Biol. 2004, 14, 744–751. [Google Scholar] [CrossRef]
- Gottlieb, J.P.; Kusunoki, M.; Goldberg, M.E. The representation of visual salience in monkey parietal cortex. Nat. Cell Biol. 1998, 391, 481–484. [Google Scholar] [CrossRef]
- Schall, J.D. On the role of frontal eye field in guiding attention and saccades. Vis. Res. 2004, 44, 1453–1467. [Google Scholar] [CrossRef] [PubMed]
- Bundesen, C.; Habekost, T.; Kyllingsbaek, S. A Neural Theory of Visual Attention: Bridging Cognition and Neurophysiology. Psychol. Rev. 2005, 112, 291–328. [Google Scholar] [CrossRef]
- Luck, S.J.; Vogel, E.K. Visual working memory capacity: From psychophysics and neurobiology to individual differences. Trends Cogn. Sci. 2013, 17, 391–400. [Google Scholar] [CrossRef]
- American Psychiatric Association (APA). Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Publishing: Arlington, VA, USA, 2013; ISBN 978-0-89042-554-1. [Google Scholar]
- Franceschini, S.; Bertoni, S.; Ronconi, L.; Molteni, M.; Gori, S.; Facoetti, A. Batteria De.Co.Ne. per la lettura. Dislessia 2016, 13, 247–319. [Google Scholar]
- Calvino, I. Marcovaldo; Giulio Einaudi: Milano, Italy, 1966. [Google Scholar]
- Mascheretti, S.; Gori, S.; Trezzi, V.; Ruffino, M.; Facoetti, A.; Marino, C. Visual motion and rapid auditory processing are solid endophenotypes of developmental dyslexia. Genes Brain Behav. 2018, 17, 70–81. [Google Scholar] [CrossRef]
- Rakhlin, N.; Cardoso-Martins, C.; Kornilov, S.A.; Grigorenko, E.L. Spelling well despite developmental language disorder: What makes it possible? Ann. Dyslexia 2013, 63, 253–273. [Google Scholar] [CrossRef] [PubMed]
- Lervåg, A.; Hulme, C. Rapid Automatized Naming (RAN) Taps a Mechanism That Places Constraints on the Development of Early Reading Fluency. Psychol. Sci. 2009, 20, 1040–1048. [Google Scholar] [CrossRef]
- Tressoldi, P.; Stella, G.; Faggella, M. The Development of Reading Speed in Italians with Dyslexia. J. Learn. Disabil. 2001, 34, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Hochstein, S.; Ahissar, M. View from the Top. Neuron 2002, 36, 791–804. [Google Scholar] [CrossRef]
- Posner, M.I. Cognitive Neuroscience of Attention; Guilford Press: New York, NY, USA, 2011. [Google Scholar]
- Cardoso-Leite, P.; Joessel, A.; Bavelier, D. Games for Enhancing Cognitive Abilities. Handbook of Game-based Learning. In Handbook of Game-based Learning; Mit Press: London, UK, 2020; Volume 18, p. 437. [Google Scholar]
- Gori, S.; Facoetti, A. How the visual aspects can be crucial in reading acquisition: The intriguing case of crowding and de-velopmental dyslexia. J. Vis. 2015, 15, 8. [Google Scholar] [CrossRef] [PubMed]
- Ekstrand, C.; Neudorf, J.; Gould, L.; Mickleborough, M.; Borowsky, R. Where words and space collide: The overlapping neural activation of lexical and sublexical reading with voluntary and reflexive spatial attention. Brain Res. 2019, 1706, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hari, R.; Renvall, H. Impaired processing of rapid stimulus sequences in dyslexia. Trends Cogn. Sci. 2001, 5, 525–532. [Google Scholar] [CrossRef]
- Sperling, A.J.; Lu, Z.-L.; Manis, F.R.; Seidenberg, M.S. Deficits in perceptual noise exclusion in developmental dyslexia. Nat. Neurosci. 2005, 8, 862–863. [Google Scholar] [CrossRef]
- Sperling, A.J.; Lu, Z.-L.; Manis, F.R.; Seidenberg, M.S. Motion-Perception Deficits and Reading Impairment. Psychol. Sci. 2006, 17, 1047–1053. [Google Scholar] [CrossRef]
- Hancock, R.; Pugh, K.R.; Hoeft, F. Neural Noise Hypothesis of Developmental Dyslexia. Trends Cogn. Sci. 2017, 21, 434–448. [Google Scholar] [CrossRef]
- Tressoldi, P.; Lorusso, M.L.; Brenbati, F.; Donini, R. Fluency remediation in dyslexic children: Does age make a difference? Dyslexia 2008, 14, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Hari, R.; Renvall, H.; Tanskanen, T. Left minineglect in dyslexic adults. Brain 2001, 124, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Facoetti, A.; Turatto, M.; Lorusso, M.L.; Mascetti, G.G. Orienting of visual attention in dyslexia: Evidence for asymmetric hemispheric control of attention. Exp. Brain Res. 2001, 138, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Hoeft, F.; McCandliss, B.D.; Black, J.M.; Gantman, A.; Zakerani, N.; Hulme, C.; Lyytinen, H.; Whitfield-Gabrieli, S.; Glover, G.H.; Reiss, A.L.; et al. Neural systems predicting long-term outcome in dyslexia. Proc. Natl. Acad. Sci. USA 2010, 108, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Ronconi, L.; Basso, D.; Gori, S.; Facoetti, A. TMS on Right Frontal Eye Fields Induces an Inflexible Focus of Attention. Cereb. Cortex 2012, 24, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, M.; Sabaroedin, K.; Silk, T.; Genc, S.; Newman, D.P.; Loughnane, G.M.; Fornito, A.; O’Connell, R.G.; Bellgrove, M.A. Evidence accumulation during perceptual decisions in humans varies as a function of dorsal frontoparietal organization. Nat. Hum. Behav. 2020, 4, 844–855. [Google Scholar] [CrossRef]
- Vandermosten, M.; Hoeft, F.; Norton, E.S. Integrating MRI brain imaging studies of pre-reading children with current theo-ries of developmental dyslexia: A review and quantitative meta-analysis. Curr. Opin. Behav. Sci. 2016, 10, 155–161. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertoni, S.; Franceschini, S.; Puccio, G.; Mancarella, M.; Gori, S.; Facoetti, A. Action Video Games Enhance Attentional Control and Phonological Decoding in Children with Developmental Dyslexia. Brain Sci. 2021, 11, 171. https://doi.org/10.3390/brainsci11020171
Bertoni S, Franceschini S, Puccio G, Mancarella M, Gori S, Facoetti A. Action Video Games Enhance Attentional Control and Phonological Decoding in Children with Developmental Dyslexia. Brain Sciences. 2021; 11(2):171. https://doi.org/10.3390/brainsci11020171
Chicago/Turabian StyleBertoni, Sara, Sandro Franceschini, Giovanna Puccio, Martina Mancarella, Simone Gori, and Andrea Facoetti. 2021. "Action Video Games Enhance Attentional Control and Phonological Decoding in Children with Developmental Dyslexia" Brain Sciences 11, no. 2: 171. https://doi.org/10.3390/brainsci11020171
APA StyleBertoni, S., Franceschini, S., Puccio, G., Mancarella, M., Gori, S., & Facoetti, A. (2021). Action Video Games Enhance Attentional Control and Phonological Decoding in Children with Developmental Dyslexia. Brain Sciences, 11(2), 171. https://doi.org/10.3390/brainsci11020171