
Neural Correlates of Music Listening: Does the Music Matter?



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Does the Music Matter? Acoustic Features as Candidate Elicitors of Induced Responses
- TIMBRAL FEATURES
- Zero Crossing rate (ZCR): number of time-domain zero crossings or sign-changes of the signal per unit of time
- Spectral centroid: geometric center on the frequency scale of the amplitude spectrum, indicating the average amount of the energy
- High-energy-low energy ratio: ratio of energy content below and above 1500 Hz.
- Spectral entropy: quantifies the spectral complexity and irregularity of energy distribution in the frequency domain
- Spectral roll-off: the frequency below which a specified percentage (cutoff) of the total spectral energy exists (85% by default)
- Spectral flux: dynamic variation of spectral information by measuring how quickly the power spectrum of a signal is changing
- Spectral spread: standard deviation of a spectrum
- Spectral flatness: tonality coefficient or Wiener entropy of a spectrum that qualifies how tone-like a sound is as opposed to being noise-like
- Sub-Band Flux: measure of fluctuation of frequency content (prominent partials) in different sub-bands of the spectrum
- Roughness: estimate of sensory dissonance
- LOUDNESS
- Root Mean Square Energy (RMS): measure of instantaneous energy contained in the signal, obtained by taking the square root of sum of the squares of the amplitude
- TONAL FEATURES
- Mode: strength of major or minor mode
- Key clarity: measure of tonal clarity
- RHYTHMIC FEATURES
- Fluctuation centroid: geometric mean of the fluctuation spectrum that represents the global repartition of rhythm periodicities within the range of 0–10 Hz, indicating the average frequency of these periodicities
- Fluctuation entropy: Shannon entropy of the fluctuation spectrum that represents the global repartition of rhythm periodicities, measuring the noisiness of the fluctuation spectrum.
- Pulse clarity: estimate of clarity of the pulse
3. From Incidental Listening to Full-Fledged Aesthetic Experience: The Role of Individual Factors
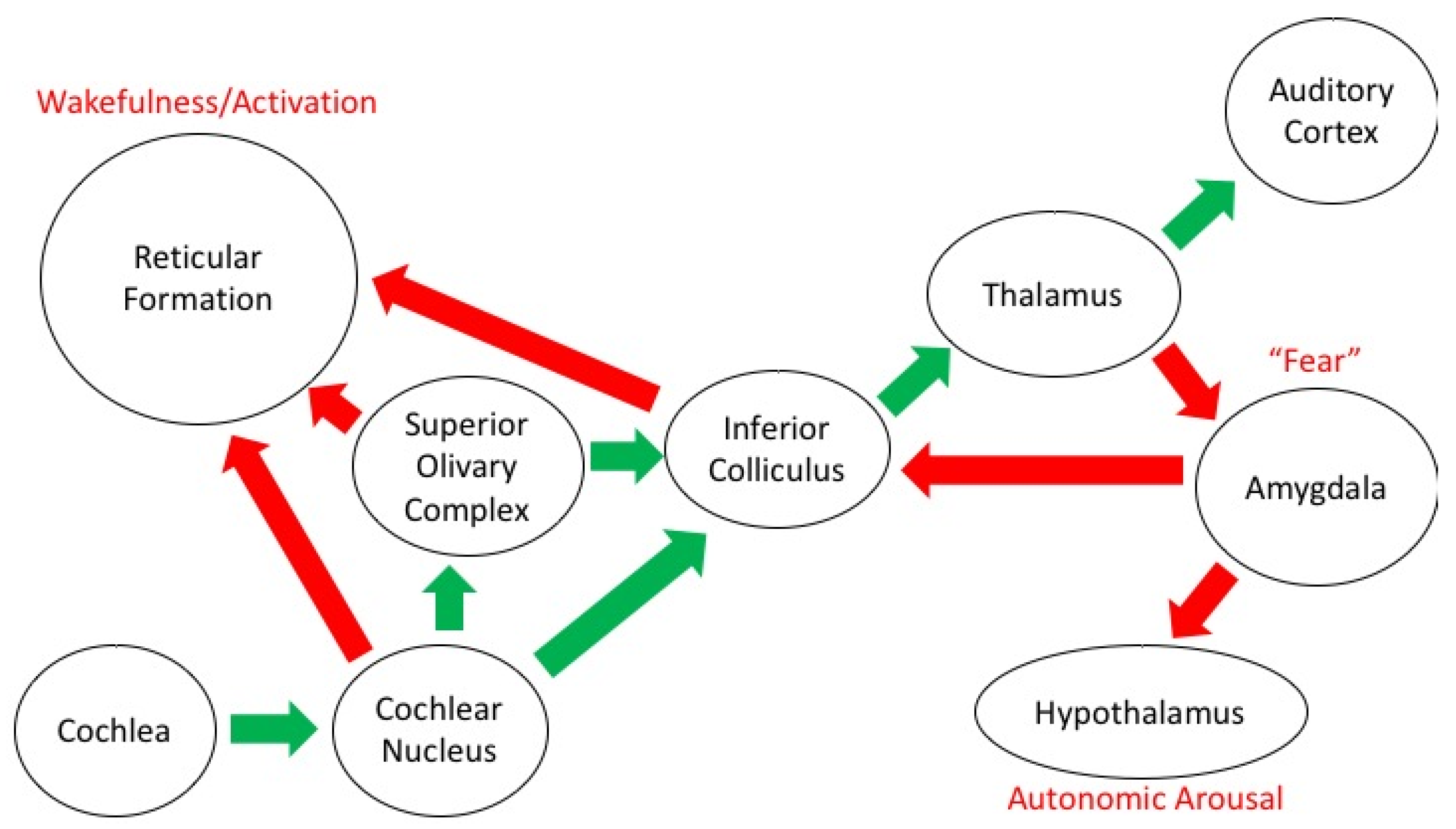
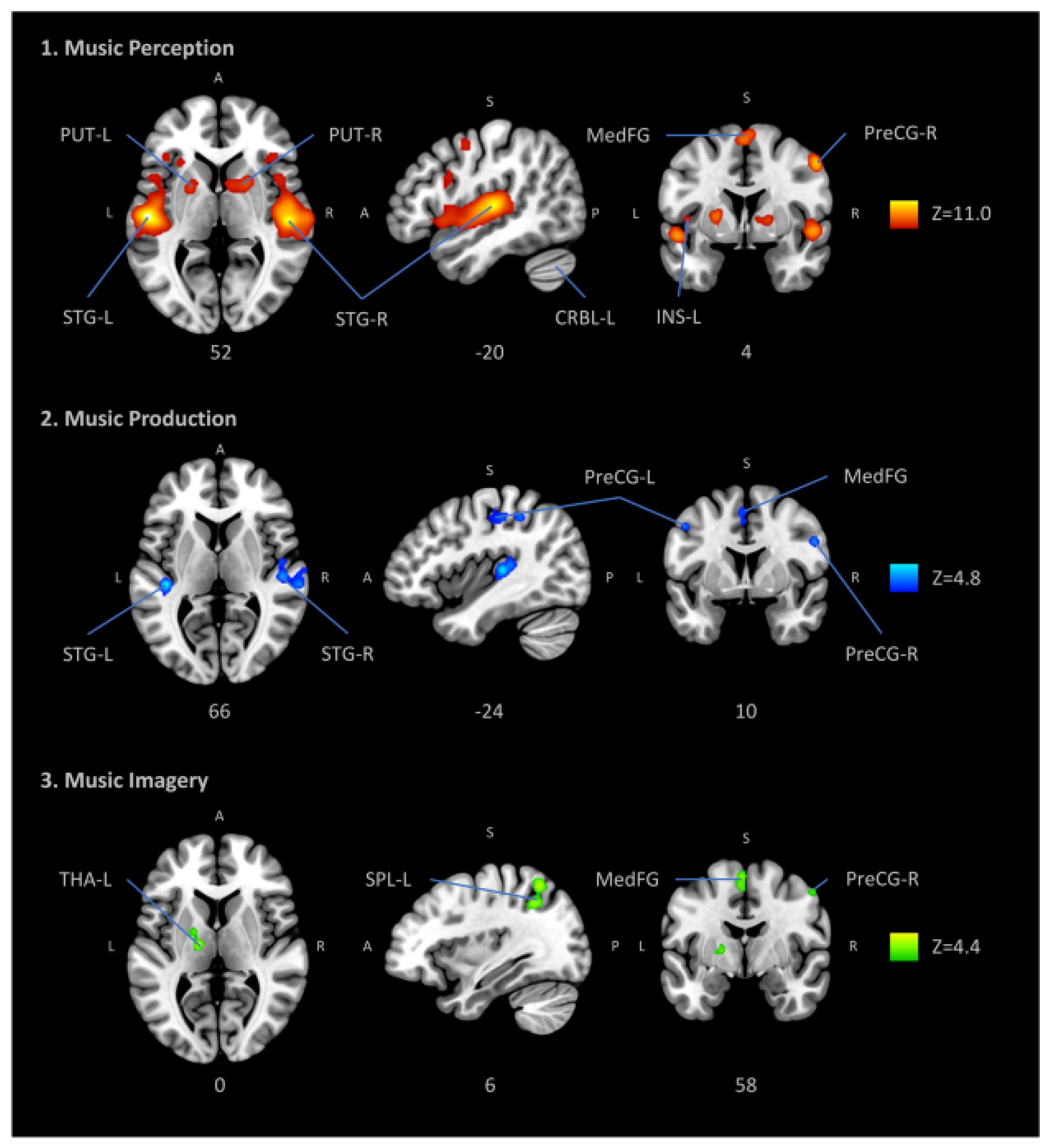
4. Neural Mechanisms of Coping with the Sounds
4.1. Aesthetic Listening and the Generation of Pleasure
4.2. Music Listening, Stress, and Allostatic Load
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Reybrouck, M.; Brattico, E. Neuroplasticity beyond Sounds: Neural Adaptations Following Long-Term Musical Aesthetic Experiences. Brain Sci. 2015, 5, 69–91. [Google Scholar] [CrossRef] [PubMed]
- Reybrouck, M.; Vuust, P.; Brattico, E. Brain Connectivity Networks and the Aesthetic Experience of Music. Brain Sci. 2018, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Reybrouck, M.; Vuust, P.; Brattico, E. Music and Brain Plasticity: How Sounds Trigger Neurogenerative Adaptations. In Neuroplasticity: Insights of Neural Reorganization; Chaban, V., Ed.; InTech: Rijeka, Croatia, 2018; pp. 85–103. [Google Scholar]
- Münte, T.F.; Altenmüller, E.; Jäncke, L. The musician’s brain as a model of neuroplasticity. Nat. Rev. Neurosci. 2002, 3, 473–478. [Google Scholar] [CrossRef]
- Sachs, M.E.; Ellis, R.E.; Schlaug, G.; Loui, P. Brain connectivity reflects human aesthetic responses. Soc. Cogn. Affect. Neurosci. 2016, 11, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Vuust, P.; Ostergaard, L.; Pallesen, K.J.; Bailey, C.; Roepstorff, A. Predictive coding of music–brain responses to rhythmic incongruity. Cortex 2009, 45, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Koelsch, S.; Vuust, P.; Friston, K. Predictive Processes and the Peculiar Case of Music. Trends Cogn. Sci. 2019, 23, P63–P77. [Google Scholar] [CrossRef]
- Brattico, E.; Pearce, M. The Neuroaesthetics of Music. Psychol. Aesthet. Creat. Arts 2013, 7, 48–61. [Google Scholar] [CrossRef]
- Reybrouck, M.; Eerola, T. Music and its inductive power: A psychobiological and evolutionary approach to musical emotions. Front. Psychol. 2017, 8, 494. [Google Scholar] [CrossRef]
- Reybrouck, M.; Podlipniak, P.; Welch, D. Music Listening as Coping Behavior: From Reactive Response to Sense-Making. Behav. Sci. 2020, 10, 119. [Google Scholar] [CrossRef]
- Ingarden, R. Aesthetic Experience and Aesthetic Object. Philos. Phenomenol. Res. 1961, 21, 289. [Google Scholar] [CrossRef]
- Ingarden, R. Phenomenological Aesthetics: An Attempt at Defining Its Range. J. Aesthet. Art Crit. 1975, 33, 257–269. [Google Scholar] [CrossRef]
- Szyszkowska, M. Roman Ingarden’s Theory of Aesthetic Experience. From Idea to Experience and Back. In Roman Ingarden and His Times; Czakon, D., Michna, N., Sosnowski, L., Eds.; Księgarnia Akademicka: Krakow, Poland, 2020; pp. 225–240. [Google Scholar]
- Dewey, J. Art as Experience; Minton, Balch & Company: New York, NY, USA, 1934. [Google Scholar]
- Alexander, T. Dewey’s Philosophy of Art and Aesthetic Experience. Arts Teach. 2016, 2, 59–67. [Google Scholar]
- Mead, G. The nature of aesthetic experience. Int. J. Ethics 1926, 36, 382–393. [Google Scholar] [CrossRef]
- Dufrenne, M. Phénoménologie de L’expérience Esthétique; Presses Universitaires de France: Paris, France, 1953. [Google Scholar]
- Milhorim, T.; Telles, T. Aesthetic perception in Dufrenne’s phenomenology: Possible contributions to psychology. Psic. Estud. 2018, 23, 137–146. [Google Scholar] [CrossRef][Green Version]
- Marković, S. Components of aesthetic experience: Aesthetic fascination, aesthetic appraisal, and aesthetic emotion. i-Perception 2012, 3, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Pelowski, M.; Markey, P.; Forster, M.; Gerger, G.; Leder, H. Move me, astonish me … delight my eyes and brain: The Vienna Integrated Model of top-down and bottom-up processes in Art Perception (VIMAP) and corresponding affective, evaluative, and neurophysiological correlates. Phys. Life Rev. 2017, 21, 80–125. [Google Scholar] [CrossRef] [PubMed]
- Leder, H.; Belke, B.; Oeberst, A.; Augustin, D. A model of aesthetic appreciation and aesthetic judgments. Br. J. Psychol. 2004, 95, 489–508. [Google Scholar] [CrossRef]
- Maruskin, L.; Thrash, T. The Chills as a Psychological Construct: Content Universe, Factor Structure, Affective Composition, Elicitors, Trait Antededents, and Consequences. J. Pers. Soc. Psychol. 2012, 103, 135–157. [Google Scholar] [CrossRef]
- Brattico, E.; Bogert, B.; Alluri, V.; Tervaniemi, M.; Eerola, T.; Jacobsen, T. It’s sad but I like it: The neural dissociation between musical emotions and liking in experts and laypersons. Front. Hum. Neurosci. 2016, 9, 676. [Google Scholar] [CrossRef]
- Liu, C.; Abu-Jamous, B.; Brattico, E.; Nandi, A.K. Towards Tunable Consensus Clustering for Studying Functional Brain Connectivity During Affective Processing. Int. J. Neural. Syst. 2017, 27, 1650042. [Google Scholar] [CrossRef]
- Liu, C.; Brattico, E.; Abu-Jamous, B.; Pereira, C.; Jacobsen, T.; Nandi, A. Effect of Explicit Evaluation on Neural Connectivity Related to Listening to Unfamiliar Music. Front. Hum. Neurosci. 2017, 11, 611. [Google Scholar] [CrossRef] [PubMed]
- Habibi, A.; Damasio, A. Music, Feelings, and the Human Brain. Psychomusicology 2014, 24, 92–102. [Google Scholar] [CrossRef]
- Chartrand, J.-P.; Peretz, I.; Belin, P. Auditory recognition expertise and domain specificity. Brain Res. 2008, 1220, 191–198. [Google Scholar] [CrossRef]
- Lartillot, O.; Toiviainen, P. MIR in Matlab (II): A Toolbox for Musical Feature Extraction from Audio. In Proceedings of the 8th International Conference on Music Information Retrieval, Vienna, Austria, 23–27 September 2007; Available online: http://ismir2007.ismir.net/proceedings/ISMIR2007_p127_lartillot.pdf (accessed on 19 October 2021).
- Alluri, V.; Toiviainen, P.; Jääskeläinen, I.P.; Glerean, E.; Sams, M.; Brattico, E. Large-scale brain networks emerge from dynamic processing of musical timbre, key and rhythm. NeuroImage 2012, 59, 3677–3689. [Google Scholar] [CrossRef]
- Toiviainen, P.; Alluri, V.; Brattico, E.; Wallentin, M.; Vuust, P. Capturing the musical brain with Lasso: Dynamic decoding of musical features from fMRI data. Neuroimage 2014, 88, 170–180. [Google Scholar] [CrossRef]
- Poikonen, H.; Alluri, V.; Brattico, E.; Lartillot, O.; Tervaniemi, M.; Huotilainen, M. Event-related brain responses while listening to entire pieces of music. Neuroscience 2016, 312, 58–73. [Google Scholar] [CrossRef] [PubMed]
- Haumann, N.T.; Lumaca, M.; Kliuchko, M.; Santacruz, J.L.; Vuust, P.; Brattico, E. Extracting human cortical responses to sound onsets and acoustic feature changes in real music, and their relation to event rate. Brain Res. 2021, 1754, 147248. [Google Scholar] [CrossRef] [PubMed]
- Vuust, P.; Brattico, E.; Seppänen, M.; Näätänen, R.; Tervaniemi, M. The sound of music: Differentiating musicians using a fast, musical multi-feature mismatch negativity paradigm. Neuropsychologia 2012, 50, 1432–1443. [Google Scholar] [CrossRef]
- Caclin, A.; Brattico, E.; Tervaniemi, M.; Näätänen, R.; Morlet, D.; Giard, M.H.; McAdams, S. Separate neural processing of timbre dimensions in auditory sensory memory. J. Cogn. Neurosci. 2006, 18, 1959–1972. [Google Scholar] [CrossRef]
- Yabe, H.; Tervaniemi, M.; Sinkkonen, J.; Huotilainen, M.; Ilmoniemi, R.J.; Näätänen, R. Temporal window of integration of auditory information in the human brain. Psychophysiology 1998, 35, 615–619. [Google Scholar] [CrossRef]
- Seppänen, M.; Brattico, E.; Tervaniemi, M. Practice strategies of musicians modulate neural processing and the learning of sound-patterns. Neurobiol. Learn. Mem. 2007, 87, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Toiviainen, P.; Burunat, I.; Brattico, E.; Vuust, P.; Alluri, V. The chronnectome of musical beat. NeuroImage 2020, 216, 116191. [Google Scholar] [CrossRef] [PubMed]
- Burunat, I.; Toiviainen, P.; Alluri, V.; Bogert, B.; Ristaniemi, T.; Sams, M.; Brattico, E. The reliability of continuous brain responses during naturalistic listening to music. NeuroImage 2016, 124, 224–231. [Google Scholar] [CrossRef]
- Burunat, I.; Tsatsishvili, V.; Brattico, E.; Toiviainen, P. Coupling of action-perception brain networks during musical pulse processing: Evidence from region-of-interest-based independent component analysis. Front. Hum. Neurosci. 2017, 11, 230. [Google Scholar] [PubMed]
- Brattico, P.; Brattico, E.; Vuust, P. Global Sensory Qualities and Aesthetic Experience in Music. Front. Neurosci. 2017, 11, 159. [Google Scholar] [CrossRef]
- Kraus, K.S.; Canlon, B. Neuronal connectivity and interactions between the auditory and limbic systems. Effects of noise and tinnitus. Hear. Res. 2012, 288, 34–46. [Google Scholar] [CrossRef]
- Welch, D.; Fremaux, G. Understanding Why People Enjoy Loud Sound. Semin. Hear. 2017, 38, 348–358. [Google Scholar]
- Reybrouck, M.; Podlipniak, P.; Welch, D. Music and Noise: Same or Different? What Our Body Tells Us. Front. Psychol. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Ruud, E. Music in therapy: Increasing possibilities for action. MAiA 2008, 1, 46–60. [Google Scholar]
- Blood, A.; Zatorre, R. Intensely pleasurable responses to music correlate with activity in brain regions implicated in reward and emotion. Proc. Natl. Acad. Sci. USA 2001, 98, 11818–11823. [Google Scholar] [CrossRef]
- Grewe, O.; Nagel, F.; Kopiez, R.; Altenmüller, E. Listening to Music as a Re-Creative Process: Physiological, Psychological, and Psychoacoustical Correlates of Chills and Strong Emotions. Music Percept. 2007, 24, 297–314. [Google Scholar]
- Halpern, D.; Blake, R.; Hillenbrand, J. Psychoacoustics of a chilling sound. Percept. Psychophy. 1986, 39, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Panksepp, J. Affective Neuroscience: The Foundations of Human and Animal Emotions; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- Panksepp, J.; Bernatzky, G. Emotional sounds and the brain: The neuro-affective foundations of musical appreciation. Behav. Proc. 2002, 60, 133–155. [Google Scholar] [CrossRef]
- Silvia, P.; Nusbaum, E. On Personality and Piloerection: Individual Differences in Aesthetic Chills and Other Unusual Aesthetic Experiences. Psychol. Aesthet. Creat. Arts 2011, 5, 208–214. [Google Scholar] [CrossRef]
- Konečni, V.J. The aesthetic trinity: Awe, being moved, thrills. Bull. Psychol. Arts 2005, 5, 27–44. [Google Scholar]
- Konečni, V.; Wanic, R.; Brown, A. Emotional and Aesthetic Antecedents and Consequences of Music-Induced Thrills. Am. J. Psychol. 2007, 120, 619–643. [Google Scholar] [CrossRef] [PubMed]
- Bannister, S. Distinct varieties of aesthetic chills in response to multimedia. PLoS ONE 2019, 14, e0224974. [Google Scholar] [CrossRef]
- Reybrouck, M.; Podlipniak, P. Preconceptual spectral and temporal cues as a source of meaning in speech and music. Brain Sci. 2019, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Elliott, T.M.; Theunissen, F.E. The modulation transfer function for speech intelligibility. PLoS Comput. Biol. 2009, 5, e1000302. [Google Scholar] [CrossRef] [PubMed]
- Chi, T.; Gao, Y.; Guyton, M.C.; Ru, P.; Shamma, S. Spectrotemporal modulation transfer functions and speech intelligibility. J. Acoust. Soc. Am. 1999, 106, 2719–2732. [Google Scholar] [CrossRef]
- Theunissen, F.E.; Elie, J.E. Neural processing of natural sounds. Nat. Rev. Neurosci. 2014, 15, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Brattico, E.; Varankaité, U. Aesthetic empowerment through music. Music Sci. 2019, 23, 285–303. [Google Scholar] [CrossRef]
- Arnal, L.; Flinker, A.; Kleinsch midt, A.; Giraud, A.-L.; Poeppel, D. Human Screams Occupy a Privileged Niche in the Communication Soundscape. Curr. Biol. 2015, 25, 2051–2056. [Google Scholar] [CrossRef]
- Filippi, P.; Congdon, J.; Hoang, J.; Bowling, D.; Reber, S.; Pašukonis, A.; Hoeschele, M.; Ocklenburg, S.; de Boer, B.; Sturdy, C.; et al. Humans recognize emotional arousal in vocalizations across all classes of terrestrial vertebrates: Evidence for acoustic universals. Proc. R. Soc. B. 2017, 284, 20170990. [Google Scholar] [CrossRef] [PubMed]
- Briefer, E. Vocal expression of emotions in mammals: Mechanisms of production and evidence. J. Zool. 2012, 288, 1–20. [Google Scholar] [CrossRef]
- Taylor, A.; Reby, D. The contribution of source–filter theory to mammal vocal communication research. J. Zool. 2010, 280, 221–236. [Google Scholar] [CrossRef]
- Zimmermann, E.; Leliveld, L.; Schehka, S. Toward the Evolutionary Roots of Affective Prosody in Human Acoustic Communication: A Comparative Approach to Mammalian Voices. In Evolution of Emotional Communication: From Sounds in Nonhuman Mammals to Speech and Music in Man; Zimmermann, E.; Leliveld, L.; Schehka, S. Oxford University Press: Oxford, UK, 2013; pp. 116–132. [Google Scholar]
- Persico, G.; Antolini, L.; Vergani, P.; Costantini, W.; Nardi, M.T.; Bellotti, L. Maternal singing of lullabies during pregnancy and after birth: Effects on mother–infant bonding and on newborns’ behaviour. Concurrent Cohort Study. Women Birth 2017, 30, e214–e220. [Google Scholar] [CrossRef]
- Chanda, M.; Levitin, D. The neurochemistry of music. Trends Cogn. Sci. 2013, 17, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Gerra, G.; Zaimovic, A.; Franchini, D.; Palladino, M.; Giucastro, G.; Reali, N.; Maestri, D.; Caccavari, R.; Delsignore, R.; Brambilla, F. Neuroendocrine responses of healthy volunteers to ‘techno-music’: Relationships with personality traits and emotional state. Int. J. Psychophysiol. 1998, 28, 99–111. [Google Scholar] [CrossRef]
- Račić, L. On the aesthetics of rock music. Int. Rev. Aesthet. Sociol. 1981, 12, 199–202. [Google Scholar] [CrossRef]
- von Appen, R. On the aesthetics of popular music. Mus. Today 2007, 8, 5–25. [Google Scholar]
- Atienda, M.; Cantero, J.L.; Dominguez-Marin, E. Mismatch negativity (MMN): An objective measure of sensory memory and long-lasting memories during sleep. Int. J. Psychophysiol. 2002, 46, 215–225. [Google Scholar] [CrossRef]
- Brattico, E.; Bogert, B.; Jacobsen, T. Toward a neural chronometry for the aesthetic experience of music. Front. Psychol. 2013, 4, 1–21. [Google Scholar] [CrossRef]
- Reybrouck, M. Musical universals and the axiom of psychobiological equivalence. In Topicality of Musical Universals/Actualité des Universaux Musicaux; Leroy, J.-L., Ed.; Editions des Archives Contemporaines: Paris, France, 2013; pp. 31–44. [Google Scholar]
- Hargreaves, D.J.; North, A.C. Experimental aesthetics and liking for music. In Series in Affective Science. Handbook of Music and Emotion: Theory Research Applications; Juslin, P.N., Sloboda, J.A., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 515–546. [Google Scholar]
- Digman, J. Personality structure: Emergence of the five-factor model. Annu. Rev. Psychol. 1990, 41, 417–440. [Google Scholar] [CrossRef]
- Chamorro-Premuzic, T.; Furnham, A. Personality and music: Can traits explain how people use music in everyday life? Br. J. Psychol. 2007, 98, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Rentfrow, P.J.; Goldberg, L.R.; Levitin, D.J. The structure of musical preferences: A five-factor model. J. Pers. Soc. Psychol. 2011, 100, 1139–1157. [Google Scholar] [CrossRef] [PubMed]
- Hurlemann, R.; Patin, A.; Onur, O.; Cohen, M.; Baumgartner, T.; Metzler, S.; Dziobek, I.; Gallinat, J.; Wagner, M.; Maier, W.; et al. Oxytocin enhances amygdala-dependent, socially reinforced learning and emotional empathy in humans. J. Neurosci. 2010, 30, 4999–5007. [Google Scholar] [CrossRef]
- Huron, D.; Vuoskoski, J. On the Enjoyment of Sad Music: Pleasurable Compassion Theory and the Role of Trait Empathy. Front. Psychol. 2020, 11, 1060. [Google Scholar] [CrossRef]
- Lackner, C.L.; Bowman, L.C.; Sabbagh, M.A. Dopaminergic functioning and preschoolers’ theory of mind. Neuropsychologia 2010, 48, 1767–1774. [Google Scholar] [CrossRef]
- Shamay-Tsoory, S.G.; Aharon-Peretz, J. Dissociable prefrontal networks for cognitive and affective theory of mind: A lesion study. Neuropsychologia 2007, 45, 3054–3067. [Google Scholar] [CrossRef] [PubMed]
- Shamay-Tsoory, S.G.; Aharon-Peretz, J.; Perry, D. Two systems for empathy: A double dissociation between emotional and cognitive empathy in inferior frontal gyrus versus ventromedial prefrontal lesions. Brain 2009, 132, 617–627. [Google Scholar] [CrossRef]
- Shamay-Tsoory, S.G. The neural bases for empathy. Neuroscientist 2011, 17, 18–24. [Google Scholar] [CrossRef]
- Eerola, T.; Vuoskoski, J.; Kautiainen, H. Being moved by unfamiliar sad music is associated with high empathy. Front. Psychol. 2016, 7, 1176. [Google Scholar] [CrossRef] [PubMed]
- Vuoskoski, J.; Eerola, T. The Pleasure Evoked by Sad Music is Mediated by Feelings of Being Moved. Front. Psychol. 2017, 8, 439. [Google Scholar] [CrossRef]
- Liang, C.; Earl, B.; Thompson, I.; Whitaker, K.; Cahn, S.; Xiang, J.; Fu, Q.-J.; Zhang, F. Musicians Are Better than Non-musicians in Frequency Change Detection: Behavioral and Electrophysiological Evidence. Front. Neurosci. 2016, 10, 464. [Google Scholar] [CrossRef]
- Criscuolo, A.; Pando-Naude, V.; Bonetti, L.; Vuust, P.; Brattico, E. Rediscovering the musician’s brain: A systematic review and meta-analysis. bioRxiv 2021. Preprint. [Google Scholar] [CrossRef]
- Perlovsky, L. Musical emotions: Functions, origins, evolution. Phys. Life Rev. 2010, 7, 2–27. [Google Scholar] [CrossRef] [PubMed]
- Brattico, E. From pleasure to liking and back: Bottom-up and top-down neural routes to the aesthetic enjoyment. In Art, Aesthetics, and the Brain; Huston, J., Nadal, M., Mora, F., Agnati, L., Cela Conde., C.J., Eds.; Oxford University Press: Oxford, UK, 2015; pp. 303–318. [Google Scholar]
- Altenmüller, E. How Many Music Centres Are in the Brain? In The Cognitive Neuroscience of Music; Peretz, I., Zatorre, R., Eds.; Oxford University Press: Oxford NY, USA, 2003; pp. 346–353. [Google Scholar]
- Lent, R. Toward a Unifying Theoretical and Practical Perspective on Well-Being and Psychosocial Adjustment. J. Couns. Psychol. 2004, 51, 482–509. [Google Scholar] [CrossRef]
- Ryff, C. Psychological well-being in adult life. Curr. Dir. Psychol. Sci. 1995, 4, 99–104. [Google Scholar] [CrossRef]
- Waterman, A. Two Conceptions of Happiness: Contrasts of Personal Expressiveness (Eudaimonia) and Hedonic Enjoyment. J. Pers. Soc. Psychol. 1993, 64, 678–691. [Google Scholar] [CrossRef]
- Nadal, M.; Skov, M. Introduction to the Special Issue: Toward an Interdisciplinary Neuroaesthetics. Psychol. Aesthet. Creat. Arts 2013, 7, 1–12. [Google Scholar] [CrossRef]
- Zatorre, R.J.; Salimpoor, V.N. From perception to pleasure: Music and its neural substrates. Proc. Natl. Acad. Sci. USA 2013, 110, 10430–10437. [Google Scholar] [CrossRef]
- Belfi, A.; Loui, P. Musical anhedonia and rewards of music listening: Current advances and a proposed model. Ann. N. Y. Acad. Sci. 2020, 1464, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Kringelbach, M.; Berridge, K. Towards a functional neuroanatomy of pleasure and happiness. Trends Cogn. Sci. 2009, 13, 479–487. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, R.; Kreutz, G.; Mitchell, L. (Eds.) Music Health and Wellbeing; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Quarto, T.; Blasi, G.; Pallesen, K.J.; Bertolino, A.; Brattico, E. Implicit Processing of Visual Emotions Is Affected by Sound-Induced Affective States and Individual Affective Traits. PLoS ONE 2014, 9, e103278. [Google Scholar] [CrossRef] [PubMed]
- Panksepp, J. The Emotional Sources of “Chills” Induced by Music. Music Percept. 1995, 13, 171–207. [Google Scholar] [CrossRef]
- Goldstein, A. Thrills in response to music and other stimuli. Physiol. Psychol. 1980, 8, 126–129. [Google Scholar] [CrossRef]
- Garza Villarreal, E.A.; Brattico, E.; Vase, L.; Østergaard, L.; Vuust, P. Superior Analgesic Effect of an Active Distraction versus Pleasant Unfamiliar Sounds and Music: The Influence of Emotion and Cognitive Style. PLoS ONE 2012, 7, e29397. [Google Scholar]
- Brattico, E.; Jacobsen, T.; De Baene, W.; Glerean, E.; Tervaniemi, M. Cognitive vs. affective listening modes and judgements of music–An ERP study. Biol. Psychol. 2010, 85, 393–409. [Google Scholar] [CrossRef]
- Ellison, D.; Moisseinen, N.; Fachner, J.; Brattico, E. Affective versus cognitive responses to musical chords: An ERP and behavioral study. Psychomusicology 2015, 25, 423. [Google Scholar] [CrossRef]
- Bogert, B.; Numminen-Kontti, T.; Gold, B.; Sams, M.; Numminen, J.; Burunat, I.; Lampinen, J.; Brattico, E. Hidden sources of joy, fear, and sadness: Explicit versus implicit neural processing of musical emotions. Neuropsychologia 2016, 8, 393–402. [Google Scholar] [CrossRef]
- Hodges, D.A. The neuroaesthetics of music. In The Oxford Handbook of Music Psychology; Hallam, S., Cross, I., Thaut, M., Eds.; Oxford University Press: Oxford, UK, 2016; pp. 247–262. [Google Scholar]
- Brattico, E. The Neuroaesthetics of Music: A Research Agenda Coming of Age. In The Oxford Handbook of Music and the Brain; Thaut, M., Hodges, D., Eds.; Oxford University Press: Oxford, UK, 2020; pp. 364–390. [Google Scholar]
- Brown, S.; Dissanayake, E. The arts are more than aesthetics: Neuroaesthetics as narrow aesthetics. In Neuroaesthetics; Skov, M., Vartanian, O., Eds.; Baywood: Amityville, NY, USA, 2009; pp. 43–57. [Google Scholar]
- Chatterjee, A. Neuroaesthetics: A Coming of Age Story. J. Cogn. Neurosci. 2010, 23, 53–62. [Google Scholar] [CrossRef]
- Leder, H. Next Steps in Neuroaesthetics: Which Processes and Processing Stages to Study? Psychol. Aesthet. Creat. Arts 2013, 7, 27–37. [Google Scholar] [CrossRef]
- Pearce, M.T.; Zaidel, D.W.; Vartanian, O.; Skov, M.; Leder, H.; Chatterjee, A.; Nadal, M. Neuroaesthetics: The Cognitive Neuroscience of Aesthetic Experience. Perspect. Psychol. Sci. 2016, 11, 265–279. [Google Scholar] [CrossRef]
- Zaidel, D.W. Brain and art: Neuro-clues from intersection of disciplines. In Neuroaesthetics; Skov, M., Vartanian, O., Eds.; Baywood: Amityville, NY, USA, 2009; pp. 153–170. [Google Scholar]
- Zaidel, D.; Nadal, M. Brain intersections of aesthetics and morals: Perspectives from biology, neuroscience, and evolution. Perspect. Biol. Med. 2011, 54, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Reybrouck, M. Musical Sense-Making. Enaction Experience and Computation; Routledge: Abingdon, NY, USA, 2021. [Google Scholar]
- Lindquist, K.; Wager, T.; Kober, H.; Bliss-Moreau, E.; Feldman Barrett, L. The brain basis of emotion: E meta-analytic review. Behav. Brain Sci. 2012, 35, 121–143. [Google Scholar] [CrossRef]
- Peciña, S.; Smith, K.; Berridge, K. Hedonic hot spots in the brain. Neuroscientist 2006, 12, 500–511. [Google Scholar] [CrossRef]
- Ferreri, L.; Mas-Herrero, E.; Zatorre, R.; Ripollés, P.; Gomez-Andresa, A.; Alicarta, H.; Olivé, G.; Marco-Pallarés, J.; Antonijoan, R.; Vallei, M.; et al. Dopamine modulates the reward experiences elicited by music. Proc. Natl. Acad. Sci. USA 2019, 116, 3793–3798. [Google Scholar] [CrossRef]
- Kringelbach, M.L. The Pleasure Center. Trust your Animal Instincts; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Pando-Naude, V.; Patyczek, A.; Bonetti, L.; Vuust, P. An ALE meta-analytic review of top-down and bottom-up processing of music in the brain. Sci. Rep. 2020, 11, 20813. [Google Scholar] [CrossRef]
- Jacobsen, T.; Schubotz, R.I.; Hofel, L.; Cramon, D.Y. Brain correlates of aesthetic judgment of beauty. Neuroimage 2006, 276–285. [Google Scholar] [CrossRef]
- LeDoux, J.E. Emotion, memory and the brain. Sci. Am. 1994, 270, 50–57. [Google Scholar] [CrossRef]
- Power, M.; Dalgleish, T. Two routes to emotion: Some implications of multilevel theories of emotion for therapeutic practice. Behav. Cogn. Psychother. 1999, 27, 129–141. [Google Scholar] [CrossRef]
- Salimpoor, V.N.; Benovoy, M.; Larcher, K.; Dagher, A.; Zatorre, R. Anatomically distinct dopamine release during anticipation and experience of peak emotion to music. Nat. Neurosci. 2011, 14, 257–262. [Google Scholar] [CrossRef]
- Berridge, K.C. Motivation concepts in behavioral neuroscience. Physiol. Behav. 2004, 81, 179–209. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C. The debate over dopamine’s role in reward: The case for incentive salience. Psychopharmacology 2007, 191, 391–431. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.; Kringelbach, M. Affective neuroscience of pleasure: Reward in humans and animals. Psychopharmacology 2008, 199, 457–480. [Google Scholar] [CrossRef] [PubMed]
- Wise, R. Dopamine, learning and motivation. Nat. Rev. Neurosci. 2004, 5, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Miranda, D.; Claes, M. Music listening, coping, peer affiliation and depression in adolescence. Psychol. Music 2009, 37, 215–233. [Google Scholar] [CrossRef]
- Selye, H. The Stress of Life; McGraw-Hill: New York, NY, USA, 1956. [Google Scholar]
- Selye, H. Stress without Distress; Springer Science and Business Media LLC: Berlin, Germany, 1976. [Google Scholar]
- Ryff, C.; Singer, B. The Contours of Positive Health. Psychol. Inq. 1988, 9, 1–28. [Google Scholar] [CrossRef]
- Sterling, P.; Eyer, J. Allostasis: A new paradigm to explain arousal pathology. In Handbook of Life Stress Cognition and Health; Fisher, J., Reason, J., Eds.; Wiley: New York, NY, USA, 1988; pp. 629–649. [Google Scholar]
- McEwen, B.S. Stress, adaptation, and disease. Allostasis and allostatic load. Ann. N. Y. Acad. Sci. 1998, 840, 33–44. [Google Scholar] [CrossRef]
- Vago, D.; Silbersweig, D. Self-awareness, self-regulation, and self-transcendence(S-ART): A framework for understanding the neurobiological mechanisms of mindfulness. Front. Hum. Neurosci. 2012, 6, 296. [Google Scholar] [CrossRef] [PubMed]
- Panksepp, J. Brain opioids-A neurochemical substrate for narcotic and social dependence. In Theory in Psychopharmacology; Cooper, S., Ed.; Academic: New York, NY, USA, 1981; pp. 149–175. [Google Scholar]
- Panksepp, J. Neurochemical control of moods and emotions: Amino acids to neuropeptides. In Handbook of Emotions; Lewis, M., Haviland, J.M., Eds.; Guilford: New York, NY, USA, 1993; pp. 87–106. [Google Scholar]
- Sapolsky, R.M. Stress in the wild. Sci. Am. 1990, 262, 116. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B. Central effects of stress hormones in health and disease: Understanding the protective and damaging effects of stress and stress mediators. Eur. J. Pharmacol. 2008, 583, 174–185. [Google Scholar] [CrossRef]
- Chapados, C.; Levitin, D.J. Cross-modal interactions in the experience of musical performances: Physiological correlates. Cognition 2008, 108, 639–651. [Google Scholar] [CrossRef]
- Saarikallio, S.; Erkkilä, J. The role of music in adolescents’ mood regulation. Psychol. Music 2007, 35, 88–109. [Google Scholar] [CrossRef]
- Van den Tol, A.; Edwards, J.; Heflick, N. Sad music as a means for acceptance-based coping. Mus. Sci. 2016, 20, 68–83. [Google Scholar] [CrossRef]
- Selye, H. The stress syndrome. AJN 1965, 65, 97–99. [Google Scholar]
- Perrez, M. Eustress. In The Oxford Companian to Emotion and the Affective Sciences; Sander, D., Scherer, K., Eds.; Oxford University Press: Oxford, NY, USA, 2009; p. 158. [Google Scholar]
- Keltner, D.; Haidt, J. Approaching awe, a moral, spiritual, and aesthetic emotion. Cognit. Emot. 2003, 17, 297–314. [Google Scholar] [CrossRef]
- Williams, P.; Johnson, K.; Curtis, B.; King, J.; Anderson, J. Individual differences in aesthetic engagement are reflected in resting-state fMRI connectivity: Implications for stress resilience. Neuroimage 2018, 179, 156–165. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reybrouck, M.; Vuust, P.; Brattico, E. Neural Correlates of Music Listening: Does the Music Matter? Brain Sci. 2021, 11, 1553. https://doi.org/10.3390/brainsci11121553
Reybrouck M, Vuust P, Brattico E. Neural Correlates of Music Listening: Does the Music Matter? Brain Sciences. 2021; 11(12):1553. https://doi.org/10.3390/brainsci11121553
Chicago/Turabian StyleReybrouck, Mark, Peter Vuust, and Elvira Brattico. 2021. "Neural Correlates of Music Listening: Does the Music Matter?" Brain Sciences 11, no. 12: 1553. https://doi.org/10.3390/brainsci11121553
APA StyleReybrouck, M., Vuust, P., & Brattico, E. (2021). Neural Correlates of Music Listening: Does the Music Matter? Brain Sciences, 11(12), 1553. https://doi.org/10.3390/brainsci11121553