
Effects of Force Modulation on Large Muscles during Human Cycling



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
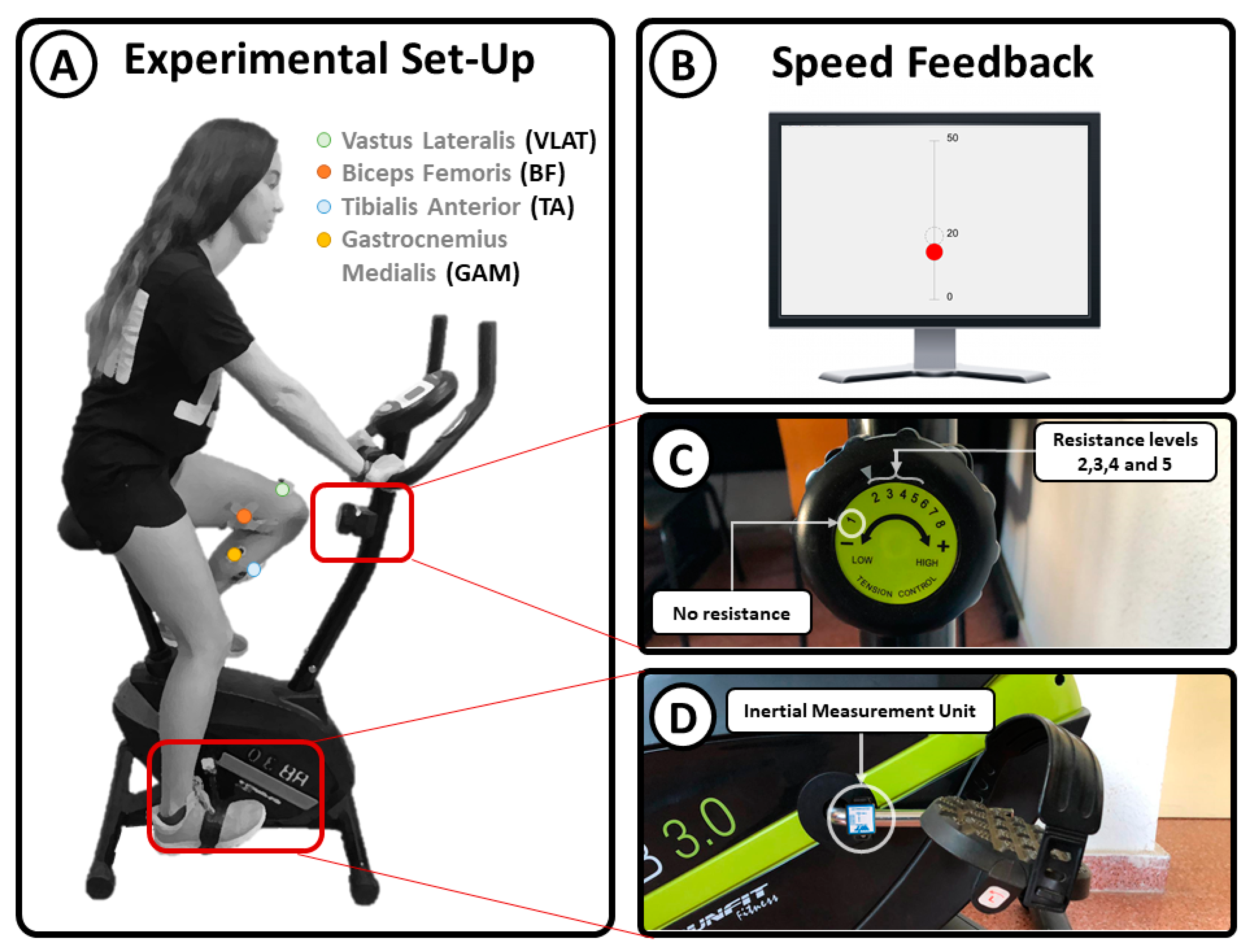
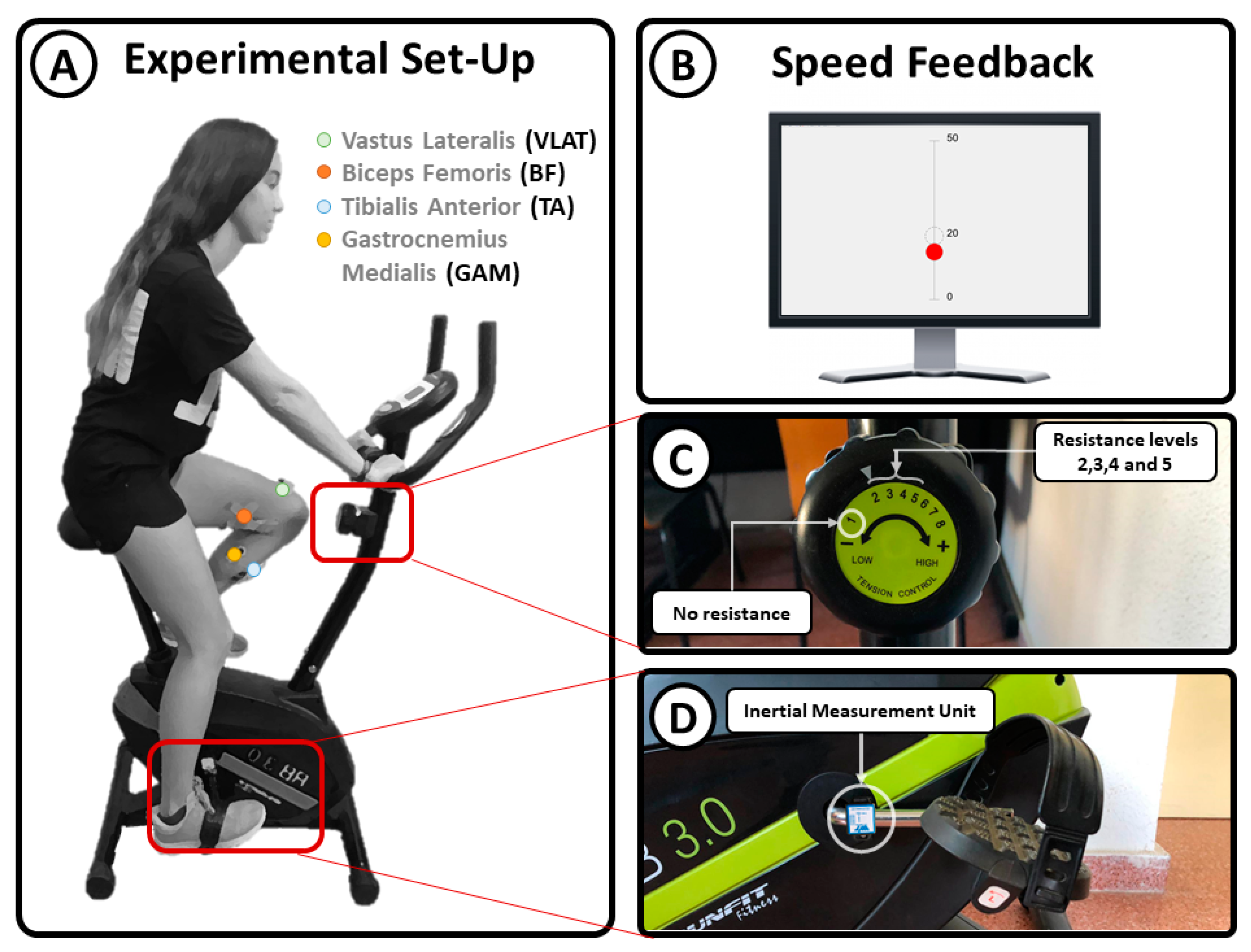
2.1. Experimental Setup
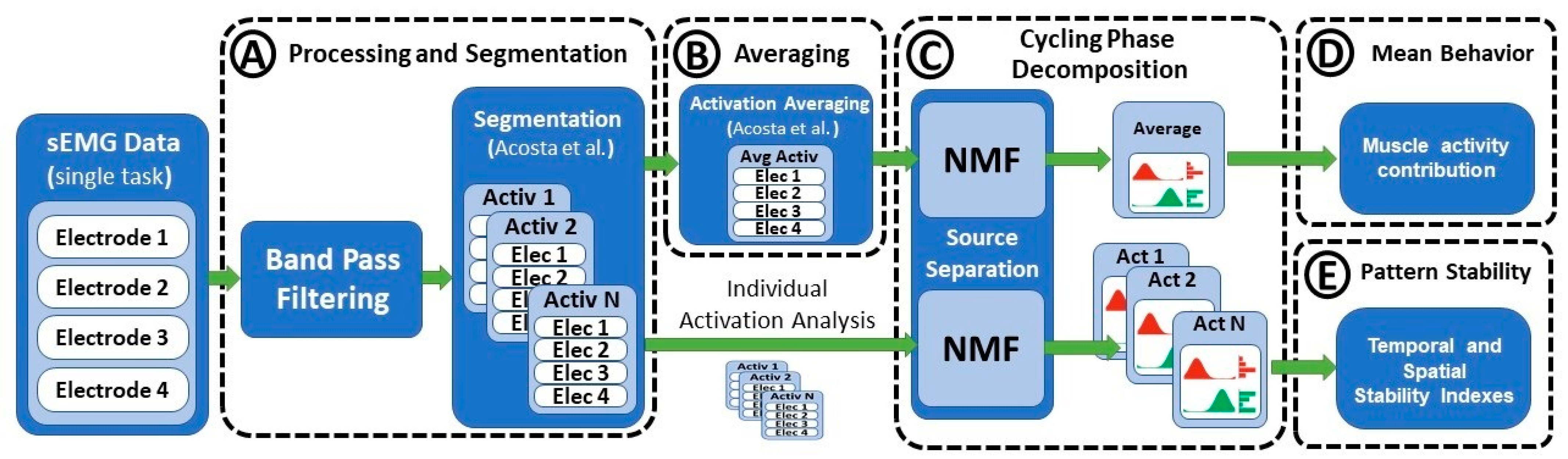
2.2. Signal Processing and Segmentation
2.3. Signal Averaging
2.4. Cycling Phase Decomposition
2.5. Evaluated Parameters
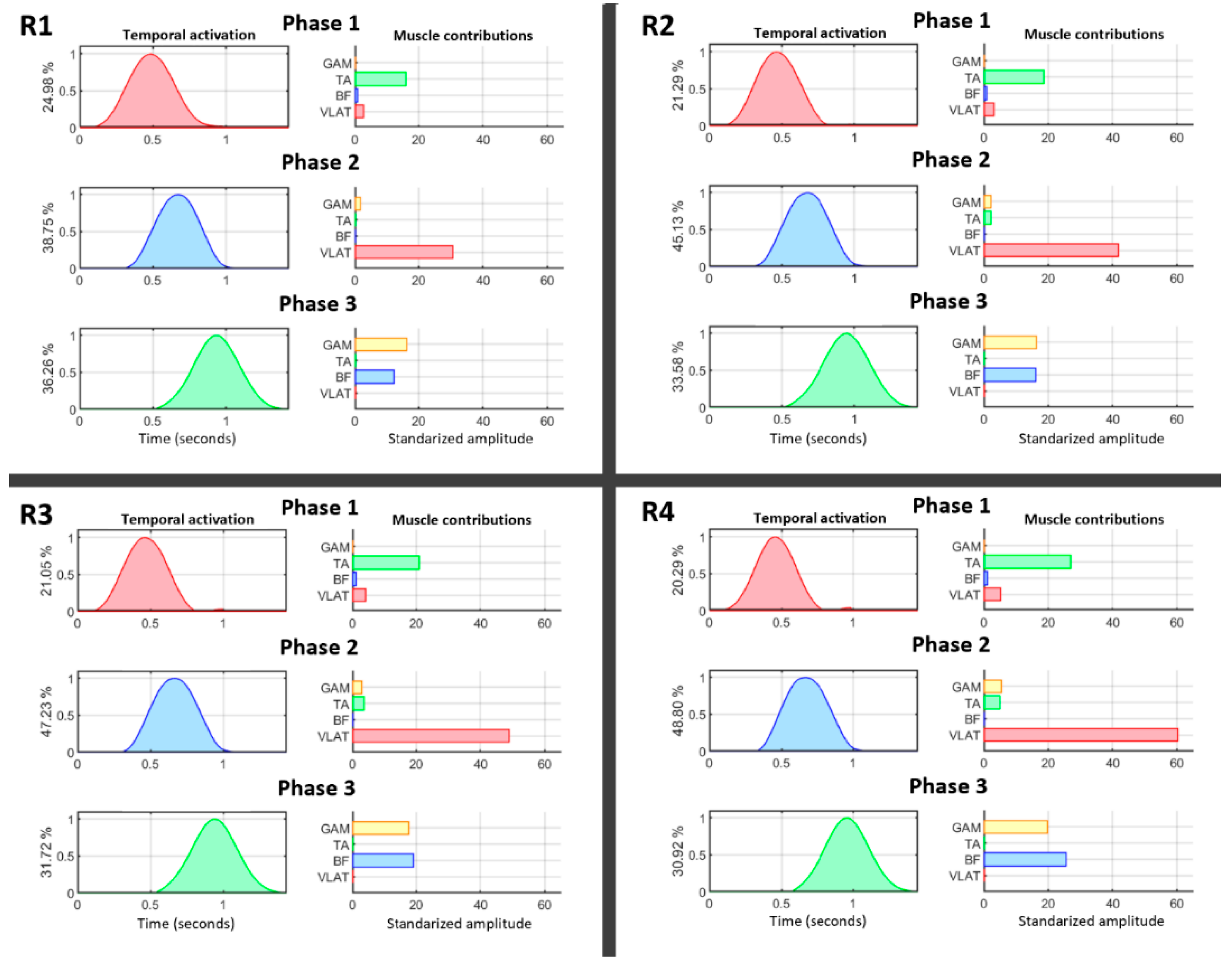
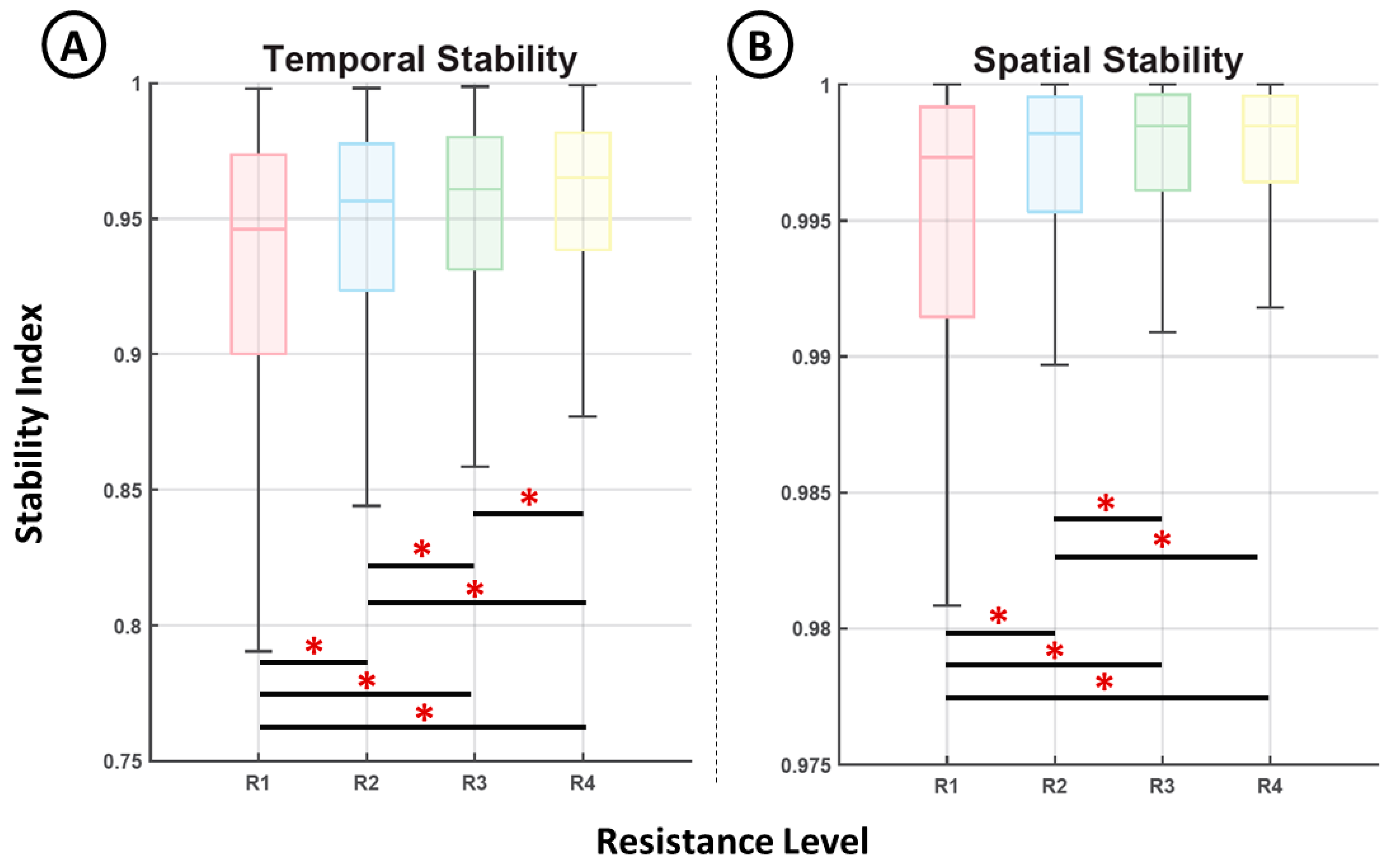
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ranganathan, V.K.; Siemionow, V.; Liu, J.Z.; Sahgal, V.; Yue, G.H. From mental power to muscle power—Gaining strength by using the mind. Neuropsychologia 2004, 42, 944–956. [Google Scholar] [CrossRef]
- Enoka, R.M. Muscle strength and its development. Sports Med. 1988, 6, 146–168. [Google Scholar] [CrossRef] [PubMed]
- Sale, D.G. Neural adaptation to resistance training. Med. Sci. Sports Exerc. 1988, 20 (Suppl. S5), S135–S145. [Google Scholar] [CrossRef] [PubMed]
- Cureton, K.J.; Collins, M.A.; Hill, D.W.; McElhannon, F.M., Jr. Muscle hypertrophy in men and women. Med. Sci. Sports Exerc. 1988, 20, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, B.J. The mechanisms of muscle hypertrophy and their application to resistance training. J. Strength Cond. Res. 2010, 24, 2857–2872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farina, D.; Negro, F. Common synaptic input to motor neurons, motor unit synchronization, and force control. Exerc. Sport Sci. Rev. 2015, 43, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Jimbo, Y.; Tateno, T.; Robinson, H.P.C. Simultaneous induction of pathway-specific potentiation and depression in networks of cortical neurons. Biophys. J. 1999, 76, 670–678. [Google Scholar] [CrossRef] [Green Version]
- Shen, L. Neural integration by short term potentiation. Biol. Cybern. 1989, 61, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.E.; Iqbal, K.; White, G.; Hutchinson, T.E. A systematic review on muscle synergies: From building blocks to a neurorehabilitation tool. Appl. Bionics. Biomech. 2018, 2018, 3615368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanenko, Y.P.; Poppele, R.E.; Lacquaniti, F. Five muscle activation patterns account for muscle activity during human locomotion. J. Physiol. 2004, 556, 267–282. [Google Scholar] [CrossRef]
- Santuz, A.; Ekizos, A.; Eckardt, N.; Kibele, A.; Arampatzis, A. Challenging human locomotion: Stability and modular organisation in unsteady conditions. Sci. Rep. 2018, 8, 2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santuz, A.; Ekizos, A.; Jahnsen, L.; Mersmann, F.; Bohm, S.; Baltzopoulos, V.; Arampatzis, A. Modular control of human movement during running: An Open Access data set. Front. Physiol. 2018, 9, 1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, A.S.; Gizzi, L.; Farina, D.; Kersting, U.G. Motor modules of human locomotion: Influence of EMG averaging, concatenation and number of step cycles. Front. Hum. Neurosci. 2014, 8, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hug, F.; Turpin, N.A.; Couturier, A.; Dorel, S. Consistency of muscle synergies during pedaling across different mechanical constraints. J. Neurophysiol. 2011, 106, 91–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marchis, C.; Castronovo, A.M.; Bibbo, D.; Schmid, M.; Conforto, S. Muscle synergies are consistent when pedaling under different biomechanical demands. In Proceedings of the 34th Annual International Conference of the IEEE EMBS, San Diego, CA, USA, 28 August–1 September 2012; pp. 3308–3311. [Google Scholar]
- Barroso, F.O.; Torricelli, D.; Moreno, J.C.; Taylor, J.; Gomez-Soriano, J.; Bravo-Esteban, E.; Piazza, S.; Santos, C.; Pons, J.L. Shared muscle synergies in human walking and cycling. J. Neurophysiol. 2014, 112, 1984–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torricelli, D.; De Marchis, C.; d’Avella, A.; Tobaruela, D.N.; Barroso, F.O.; Pons, J.L. Reorganization of muscle coordination underlying motor learning in cycling tasks. Front. Bioeng. Biotechnol. 2020, 8, 800. [Google Scholar] [CrossRef] [PubMed]
- Ivanenko, Y.; Gurfinkel, V.S. Human postural control. Front. Neurosci. 2018, 12, 171. [Google Scholar] [CrossRef] [PubMed]
- Costa-García, A.; Ozaki, K.; Yamasaki, H.; Itkonen, M.; Alnajjar, F.; Okajima, S.; Tanimoto, M.; Kondo, I.; Shimoda, S. Model for prompt and effective classification of motion recovery after stroke considering muscle strength and coordination factors. J. Neuroeng. Rehabil. 2019, 16, 130. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, C.A. Cycling biomechanics: A literature review. J. Orthop. Sports Phys. Ther. 1991, 14, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Guidetti, L.; Rivellini, G.; Figura, F. EMG patterns during running: Intra-and inter-individual variability. J. Electromyogr. Kinesiol. 1996, 6, 37–48. [Google Scholar] [CrossRef]
- Gabriel, D.A. Changes in kinematic and EMG variability while practicing a maximal performance task. J. Electromyogr. Kinesiol. 2002, 12, 407–412. [Google Scholar] [CrossRef]
- Granata, K.P.; Padua, D.A.; Abel, M.F. Repeatability of surface EMG during gait in children. Gait Posture 2005, 22, 346–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoyama, T.; Kohno, Y. Temporal and quantitative variability in muscle electrical activity decreases as dexterous hand motor skills are learned. PLoS ONE 2020, 15, e0236254. [Google Scholar] [CrossRef] [PubMed]
- Rimini, D.; Agostini, V.; Knaflitz, M. Evaluation of muscle synergies stability in human locomotion: A comparison between normal and fast walking speed. In Proceedings of the 2017 IEEE International Instrumentation and Measurement Technology Conference (I2MTC), Turin, Italy, 22–25 May 2017; pp. 1–5. [Google Scholar]
- Cheung, V.C.; Piron, L.; Agostini, M.; Silvoni, S.; Turolla, A.; Bizzi, E. Stability of muscle synergies for voluntary actions after cortical stroke in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 19563–19568. [Google Scholar] [CrossRef] [Green Version]
- Hermens, H.J.; Freriks, B.; Merletti, R.; Stegeman, D.; Blok, J.; Rau, G.; Disselhorst-Klug, C.; Hägg, G. European recommendations for surface electromyography. Roessingh Res. Dev. 1999, 8, 13–54. [Google Scholar]
- Costa-García, Á.; Iáñez, E.; Sonoo, M.; Okajima, S.; Yamasaki, H.; Ueda, S.; Shimoda, S. Segmentation and averaging of sEMG muscle activations prior to synergy extraction. IEEE Robot Autom. Lett. 2020, 5, 3106–3112. [Google Scholar] [CrossRef]
- Lee, D.D.; Seung, H.S. Learning the parts of objects by nonnegative matrix factorization. Nature 1999, 401, 788–791. [Google Scholar] [CrossRef]
- Colombo, R. Performance Measures in Robot Assisted Assessment of Sensorimotor Functions (Chapter 8) in Rehabilitation Robotics; Academic Press: Cambridge, MA, USA, 2018; pp. 101–115. [Google Scholar]
- Alnajjar, F.; Ozaki, K.-I.; Itkonen, M.; Yamasaki, H.; Tanimoto, M.; Ueda, I.; Kamiya, M.; Tournier, M.; Nagai, C.; Costa-García, Á.; et al. Self-support biofeedback training for recovery from motor impairment after stroke. IEEE Access 2020, 8, 72138–72157. [Google Scholar] [CrossRef]
- Campanini, I.; Disselhorst-Klug, W.; Rymer, W.Z.; Merletti, R. Surface EMG in clinical assessment and neurorehabilitation: Barriers limiting its use. Front. Neurol. 2020, 11, 934. [Google Scholar] [CrossRef]
- Safavynia, S.A.; Torres-Oviedo, G.; Ting, L.H. Muscle synergies: Implications for clinical evaluation and rehabilitation of movement. Top. Spinal Cord Inj. Rehabil. 2011, 17, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Sun, Y.; Xie, B.; Huang, Z.; Wu, J.; Hou, J.; Liu, Y.; Huang, Z.; Zhang, Z. Alterations of muscle synergies during voluntary arm reaching movement in subacute stroke survivors at different levels of impairment. Front. Comput. Neurosci. 2018, 12, 69. [Google Scholar] [CrossRef] [PubMed]
- Ambrosini, E.; Parati, M.; Peri, E.; De Marchis, C.; Nava, C.; Pedrocchi, A.; Ferrero, G.; Ferrante, S. Changes in leg cycling muscle synergies after training augmented by functional electrical stimulation in subacute stroke survivors: A pilot study. J. Neuroeng. Rehabil. 2020, 17, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa-García, Á.; Úbeda, A.; Shimoda, S. Effects of Force Modulation on Large Muscles during Human Cycling. Brain Sci. 2021, 11, 1537. https://doi.org/10.3390/brainsci11111537
Costa-García Á, Úbeda A, Shimoda S. Effects of Force Modulation on Large Muscles during Human Cycling. Brain Sciences. 2021; 11(11):1537. https://doi.org/10.3390/brainsci11111537
Chicago/Turabian StyleCosta-García, Álvaro, Andrés Úbeda, and Shingo Shimoda. 2021. "Effects of Force Modulation on Large Muscles during Human Cycling" Brain Sciences 11, no. 11: 1537. https://doi.org/10.3390/brainsci11111537
APA StyleCosta-García, Á., Úbeda, A., & Shimoda, S. (2021). Effects of Force Modulation on Large Muscles during Human Cycling. Brain Sciences, 11(11), 1537. https://doi.org/10.3390/brainsci11111537