3. Results
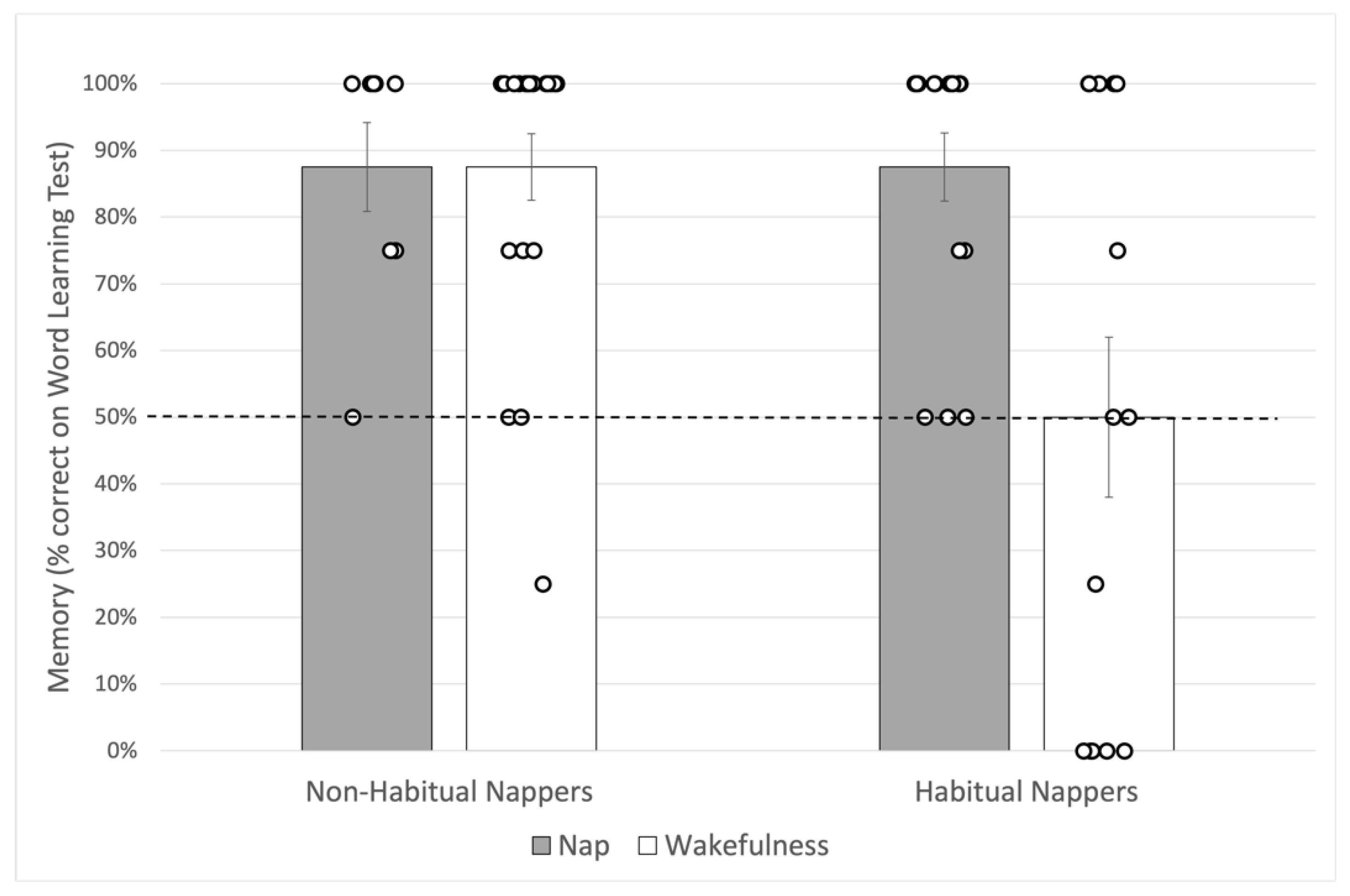
Averaged memory performance for each group in our experimental design is shown in
Figure 3.
3.1. Interaction between Napping Status and Sleep versus Wakefulness during Delay
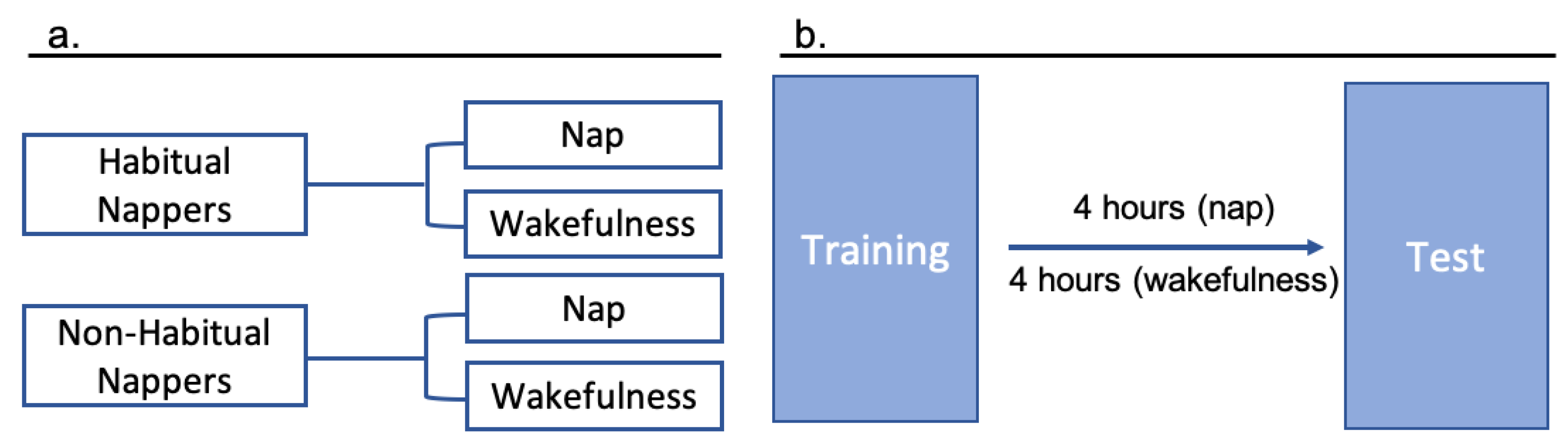
In order to determine whether typical napping behavior plays a role in the relationship between memory and sleep after a delay including a nap or wakefulness, we conducted a mixed effects regression [
29] predicting the log odds of correctly choosing the target object at test as a function of delay type (Nap or Wakefulness) and typical napping status (Habitual Napper or Non-Habitual Napper). We allowed delay type and nap status to interact in order to determine whether memory performance after a period of sleep or wakefulness would differ between habitual and non-habitual nappers. The outcome variable was the score (correct or incorrect) for each of four test trials.
Table 2 displays the results of this regression. First, napping status was a significant predictor of memory performance after a delay such that habitual napping was associated with lower odds of correctly selecting the target object at test. The interaction between delay type and typical napping behavior did not significantly predict memory performance after a delay,
p = 0.057, though trended towards significance. Importantly, as discussed above, we were not able to achieve equal group sizes due to a halt in data collection related to the coronavirus pandemic. Thus, with equal sample sizes, significance may have been achieved.
3.2. Differences in Outcomes Based on Nap Habituality
As previous studies have suggested developmental differences in memory networks between habitually napping children and non-habitually napping children [
14,
24], we divided our sample based on typical napping status and analyzed each group separately to determine any differences in memory performance in response to a period of sleep or wakefulness after learning.
We conducted a second mixed effects regression for habitually napping children and used delay type (Nap or Wakefulness) as a predictor. The results of this analysis are displayed in
Table 3. Here, we found that delay type did predict memory performance after a 4-h delay such that the odds of selecting the correct target object at test for habitually napping children who nap after learning are 59.5 times that of habitually napping children who remain awake after learning.
Finally, we conducted a third mixed effects regression for non-habitually napping children and used delay type (Nap or Wakefulness) as a predictor. The results of this analysis are displayed in
Table 4. Here, delay type did not significantly predict the log odds of selecting the target object at test, suggesting a period of sleep or wakefulness after learning does not impact memory performance for non-habitually napping children.
In order to determine whether children remembered the novel objects significantly better than would be expected by chance, trial scores were averaged for each child and compared to chance performance (50%) by experimental group, using one-tailed Wilcoxon signed rank tests. Only habitually napping children who remained awake after learning failed to perform significantly better than chance, p > 0.05. All other groups exceeded chance performance: Habitual Nappers/Nap, p < 0.001, r = 0.88; Non-Habitual Nappers/Wakefulness, p < 0.001, r = 0.86; and, despite the low group size (n = 8), Non-habitual Nappers/Nap, p = 0.009, r = 0.89 with power = 78%. As the effect size for the comparison to 50% chance in Habitual Nappers/wakefulness was exactly zero, an estimated effect size of 0.001 with the desired power of 80% would require a sample size of 6,182,559 children to detect an effect, a sample size we would not achieve, pandemic or not.
3.3. Control Variables, Standardized Measures, and Patterns of Sleep
To explore the possibility that any memory differences between habitual nappers and non-habitual nappers may stem from differences in vocabulary, we conducted an independent sample
t test on children’s standard PPVT scores (see
Table 5). This revealed no differences between habitual nappers (
M = 114.5) and non-habitual nappers (
M = 111.8);
t(48) = 0.78,
p > 0.05,
d = 0.22. Additionally, we compared habitual and non-habitual nappers on the 19 subscales of the Conners Early Childhood Questionnaire and found no differences between children based on habitual napping behavior (all
ps > 0.05). Further, we confirmed no differences in age between habitual nappers (
M = 47.0) and non-habitual nappers (
M = 47.8),
t(56) = −0.78,
p > 0.05,
d = −0.20.
Data from the 1-week sleep diary were also analyzed to confirm that the number of naps per week as reported in the sleep diary was consistent with the initial condition assignment. Complete sleep diary data was obtained from 36 children (Habitual Nappers/Nap: 10, Habitual Nappers/Wakefulness: 8, Non-Habitual Nappers/Nap: 7, and Non-Habitual Nappers/Wakefulness: 11). Though a few of the sleep diary reports were off by 2 or fewer days compared to the initial parent reported napping days, all were consistent with the condition assignment of Habitual Napper (4–7 days per week) and Non-Habitual Napper (0–3 days per week).
As training and test times were scheduled in 4 h intervals around children’s typical nap time (nap groups) or a time when the child would typically be awake (wakefulness groups), we expected to see differences in time of training and time of test between the nap and wakefulness groups. Indeed, among Habitual Nappers, children in the wakefulness group were trained and tested at a significantly earlier time of day (MTrain = 9:29 a.m. and MTest = 1:31 p.m.) compared to children in the sleep group (MTrain = 12:30 p.m. and MTest = 4:30 p.m.), t(14) = 4.03, p < 0.001, d = 1.56 for time of training and t(16) = 4.02, p < 0.001, d = 1.55 for time of test. Similarly, among Non-Habitual Nappers, children in the wakefulness group were trained at a significantly earlier time of day (MTrain = 9:58 a.m. and MTest = 1:58 p.m.) compared to children in the sleep group (MTrain = 12:22 p.m. and MTest = 4:26 p.m.), t(25) = 5.96, p < 0.001, d = 1.72 for time of training and t(25) = 6.17, p < 0.001, d = 1.77 for time of test. Importantly though, there was not a significant difference between children in the Habitual Nap/Wakefulness and Non-Habitual Nap/Wakefulness groups in time of training, t(20) = −0.61, p = 0.57, d = −0.23, or test, t(20) = −0.61, p = 0.55, d = −0.22, suggesting that the time of day training and test occurred is not related to the differences in retention we see here.
We also examined the wakefulness interval (i.e., the amount of time between last sleep and training) and found a significant difference between children in the Nap conditions (M = 5.6 h) and children in the Wakefulness conditions (M = 2.3 h), t(54) = 8.79, p < 0.001, d = 2.15.
For non-habitually napping children, parents selected the reasons their child transitioned out of naps from the following: preschool/daycare schedule, child’s desire to transition out of napping, activities that interfere with naptime, or decision from caregivers. Parents could select multiple options. The vast majority of parents indicated that at least one of the reasons for nap cessation was their child’s desire to stop napping (83%). In addition, 24% of parents reported that activities interfered with their child’s nap time, 14% of parents indicated that attending preschool or daycare prompted the transition out of naps, and 17% of parents indicated that it was a decision by the caregivers.
4. Discussion
Daytime sleep has previously been shown to impact retention of new word learning in early childhood [
8,
9,
10,
13,
15,
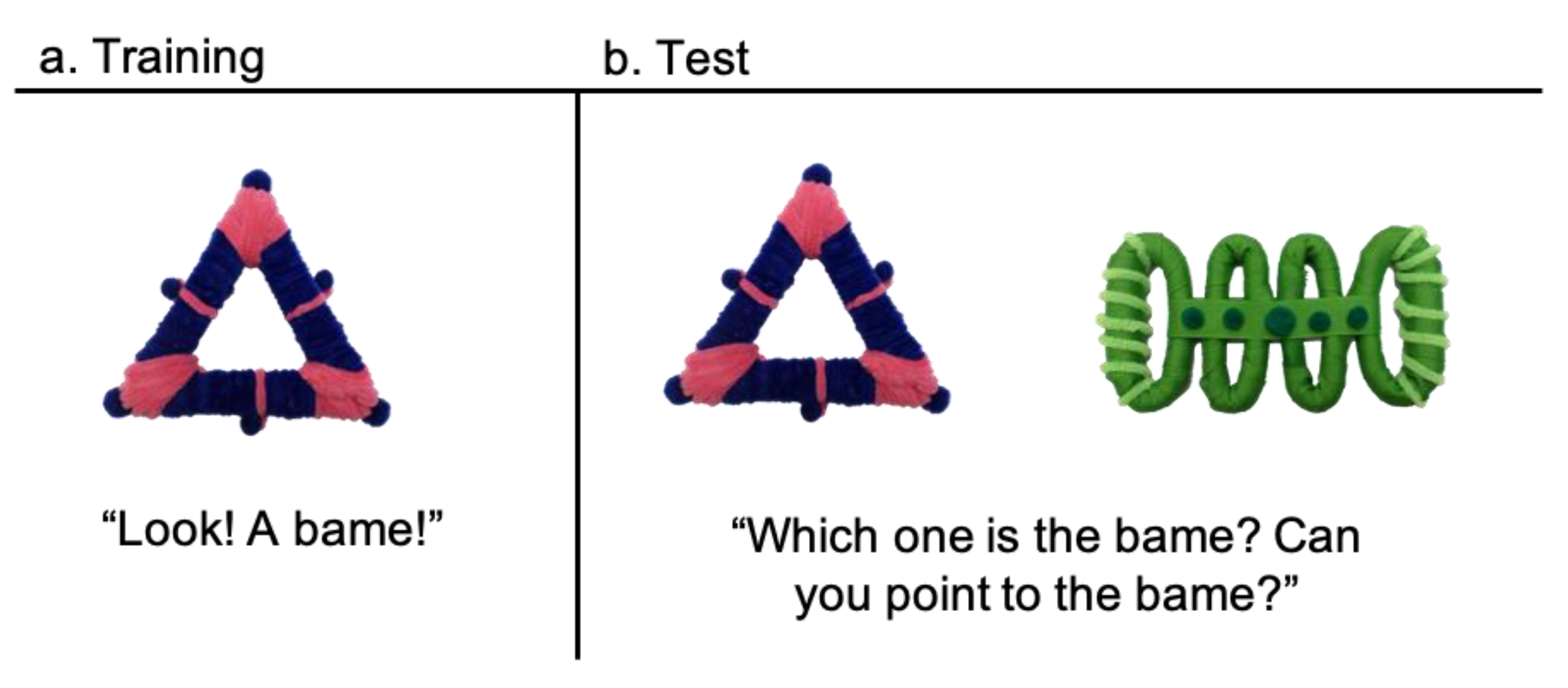
17], yet children transition out of regular napping, so the question of how this transition impacts sleep and memory in children remains unclear. Here, we examine the impact of typical napping patterns on retention of new words after a delay including either sleep or wakefulness. Children were given a single exposure to two novel words, then either took their typical nap or stayed awake during the four hours after training. Children were then tested on the object–label pairings in a two-alternative forced choice task. Here, we found a different pattern of retention based on napping status. Non-habitually napping children (those who have mostly or fully transitioned out of regular napping) remember the object–label pairings after a 4-h delay regardless of whether they nap after learning or not. Children who still nap regularly show much more variability in their responses. On average such children must take a nap after learning in order to remember the object-label pairings 4 h later. Importantly, this study also demonstrates that children are able to retain novel object–label pairings after only a brief exposure (one labeling of a novel word mapping).
These findings may suggest that as children transition out of naps, they are better able to retain novel information across longer periods of wakefulness. Interestingly, our behavioral findings map onto imaging results comparing regularly napping children with children who had transitioned out of naps. Children who rarely nap have smaller hippocampal subfield volumes [
30], consistent with findings that indicate a reduction in hippocampal subfield volume with maturation [
31]. Children who rarely napped also showed better memory on an ordered recall task than children of the same age who were still napping occasionally or regularly, leading the authors to propose that children who no longer nap regularly have more developed memory networks [
30].
Our behavioral results discussed here also complement neurophysiological data showing documented changes in slow-wave activity across early childhood. Slow-wave activity (SWA), a type of slow oscillatory electroencephalography (EEG) activity, has long been considered a marker of sleep need: SWA increases across wakefulness and decreases across sleep. Kurth et al. [
32] measured children’s naps at three different ages using electroencephalography (EEG). At each age, children’s naps were measured at three times across the day—in the morning, afternoon, and evening. Among the developmental changes in nap physiology, a notable finding is a decrease in slow-wave activity (SWA) in children’s naps as they mature. SWA has long been considered a physiological marker of sleep pressure [
33], so these findings suggest that as children age, they accumulate sleep pressure at a slower rate, allowing them to remain awake for longer periods of time, and eventually phase out napping. As such, were we to add polysomnography to our design, we might expect to see higher SWA in the naps of habitually napping children that should correlate with retention compared to those who no longer nap habitually.
Importantly, we observe a different pattern of results than two previous studies that also examined children’s napping status and word learning. Sandoval et al. [
15] demonstrated that, regardless of napping status, young 3-year-olds generalized novel verb learning when they had a nap soon after learning—but not when they remained awake after learning. Similarly, Williams and Horst [
17] showed that non-habitually napping 3.5-year-olds who stayed awake after learning novel words did not retain as much of their learning as habitually napping children who had napped after learning. In the current study, we purposely recruited children from a slightly older age range (42–53 months,
M = 47 months, or just under 4-years-old). Thus, is it possible that children who transition out of napping earlier in life may show a different response to periods of wakefulness after learning than children who transition out of naps slightly later.
In the current study, a notable finding is that children in the Wakefulness groups showed a striking difference in memory performance across a 4-h period of wakefulness based on their typical napping status. Though the same age on average, habitually napping children who stayed awake after learning performed much worse on average (no better than chance) on the memory task compared to non-habitually napping children who stayed awake after learning. Importantly, these children were not deprived of a nap—they were simply scheduled during a time they would already be awake. This finding may suggest that, as children transition out of naps, they become less susceptible to interference across periods of wakefulness (see Riggins and Spencer [
24] for a similar argument). Indeed, evidence from both human and animal studies show that memory representations are susceptible to interference during waking and are resistant to interference once sleep is initiated [
34]. More mature memory networks [
24] may make non-habitual nappers more resistant to interference during wakefulness compared to habitual nappers.
A recent theory on memory formation and retention, the contextual binding (CB) theory, proposes that sleep benefits memory, not through active consolidation, but through a reduction in contextual interference [
23]. In this theory, the hippocampus binds together contextual elements of an experience (for example, spatial, temporal, cognitive aspects, etc.) and according to CB, gradual changes in context result in memories that share similar context, which causes interference and forgetting. In this view, memories encoded soon before sleep are subject to less contextual interference, as sleep reduces the possibility of encoding new information that would share a similar context as the learning experience. The results of the current study can be interpreted within this framework as we see that, regardless of typical napping behavior, children who sleep soon after learning retain the object–label associations. However, this model would also predict that children who remain awake after learning would be subject to contextual interference as representations for experiences they encounter soon after learning would share some similar context. Importantly, we only see this pattern in children who nap habitually—non-habitually napping children who remain awake after learning retain the object–label associations, even after a period of wakefulness. Considered within the framework of CB, perhaps children who have transitioned out of naps are less susceptible to contextual interference than typical nappers. If children who are still napping regularly have less mature memory networks, as suggested by Riggins and Spencer [
24], perhaps they have less distinct contextual representations that lead to greater contextual overlap between memories and greater forgetting.
Another possibility is that non-habitual nappers may be more cognitively mature and therefore any differences in retention between non-habitual nappers and habitual nappers may be attributed to cognitive differences between the two. However, unpublished work in our laboratory using this paradigm showed equal performance for habitual and non-habitual nappers on an immediate test (habitual nappers,
M = 0.69,
SD = 0.37; non-habitual nappers,
M = 0.73,
SD = 0.35,
t(46)
= −0.45,
p > 0.05,
d = −0.13), suggesting equal encoding regardless of napping status. Additionally, in the current study, we found no evidence for vocabulary differences between habitual and non-habitual nappers. This finding is consistent with a previous study that found no difference in PPVT scores between non-habitually napping and habitually napping 3-year-olds [
15]. These findings are inconsistent with Lam et al. [
35] who showed that smaller vocabularies were associated with more napping during the week. However, as discussed in Sandoval et al. [
15], differences in nighttime sleep duration may play help explain this difference. Children in the Lam et al. [
35] study obtained a lower amount of nighttime sleep than is recommended for that age group [
36]. Therefore, if children are obtaining too little nighttime sleep, compensatory napping during the day may interfere with their nighttime sleep in either duration or quality, which may in turn impact cognitive functioning.
We did find a significant difference in the wakefulness interval between when children last slept and training such that children in the Nap groups, overall, experienced a longer waking interval before training than did children in the Wakefulness groups. This finding is not surprising, given the study design and scheduling restrictions based on children’s typical sleep and wakefulness patterns. However, one concern could be that a shorter wakefulness interval for the wakefulness group might result in better encoding and hence better retention than for the nap group. If this were the case, we would expect to see the best performance in the wakefulness groups (Non-Habitual Nappers/Wakefulness and Habitual Nappers/Wakefulness) and the worst performance in the nap groups (Non-Habitual Nappers/Nap and Habitual Nappers/Nap), but this is not what we observe. In fact, we see excellent performance in both Nap groups, and we see the worst performance in the Habitual-Napper/Wakefulness group. Furthermore, if we compare wakefulness intervals across napping status there are no differences (Non-Habitual Nappers, M = 3.63 h and Habitual Nappers, M = 3.69 h). If we look just within the Wakefulness groups, Non-Habitual Nappers averaged 2.86 h of wakefulness before training and Habitual Nappers averaged 1.56 h of wakefulness before training, and yet this latter group showed the worst performance. Thus, it does not seem that a short wakefulness interval before training aids performance.
Additionally, as expected, we did find that children in the wakefulness groups were trained and tested earlier than children in the nap groups. This is not surprising, as children’s naps tend to occur mid-day, and the wakefulness groups’ training and test were scheduled around nap times. Importantly, however, there were no differences in training and test times between Habitual Nappers and Non-Habitual Nappers who remained awake after training, which is where we observed a difference in retention. This suggests that circadian differences during training and test cannot explain differences in retention between these two groups.
The current study has a few limitations. First, most parents of non-habitual nappers in this study indicated that their child transitioned out of regular napping of their own desire. This may indicate that children in the current study transitioned out of naps as they were ready, rather than due to external pressures. It is possible that the pattern we found here may reflect the relationship between sleep and memory across a natural nap transition, and children who transition out of naps due to external factors may show a different pattern of response to a period of wakefulness.
Second, as indicated above, we scheduled children at times of day that accorded with their natural sleep and wakefulness schedules. Thus, we trained and tested children in sleep and wakefulness groups at different times of day. However, circadian effects did not contribute to our main finding as habitual and nonhabitual nappers in the wakefulness groups trained and tested at the same times of day.
Additionally, the current study cannot discern whether the pattern of results found could be attributed to active processes during sleep or the nap offering protection from interference. One previous study has shown that spindle activity in children’s naps correlates positively with their change in memory performance on an object–location memory task, suggesting sleep played an active role in consolidation [
14]. However, we did not employ polysomnography in the current study. Future studies could measure nap physiology after novel word learning in children to address this. Furthermore, future studies should specifically measure the impact of interference during a period of wakefulness in non-habitual nappers compared to habitual nappers. If non-habitual nappers are less impacted by an interference task after novel learning compared to habitual nappers, as would be predicted from the findings in our current study, it would be evidence that, as children transition out of naps, they become less susceptible to interference allowing them to retain novel information across longer periods of wakefulness.
Finally, due to the coronavirus pandemic, data collection was halted, so complete samples could not be obtained. With a complete sample, we may have been able to better detect an interaction between habitual napping status and delay type. Furthermore, the size of our Non-Habitual Nappers/Nap group is relatively small (n = 8). We anticipated this problem, given that once children have transitioned out of regular napping, it may be difficult for them to nap during the day—a fact that is reflected in the number of children who were originally assigned to this group but were unable to nap during the delay (n = 5). However, a power analysis was conducted for this group showing that, despite the low group size, sufficient power (78%) was achieved. Additionally, our best estimate of effect size for our Habitual Nappers/Wakefulness group requires a sample size of 6,182,599 children to obtain significance, underscoring the fact that our target N would not result in significance for this group.



{kind=link}
{kind=link}
{kind=link}