n-Alkane Distribution—A Paleovegetation Change Indicator during the Period from Late Glacial to Late Holocene on Russian Plain (Bryansk Region)

Abstract
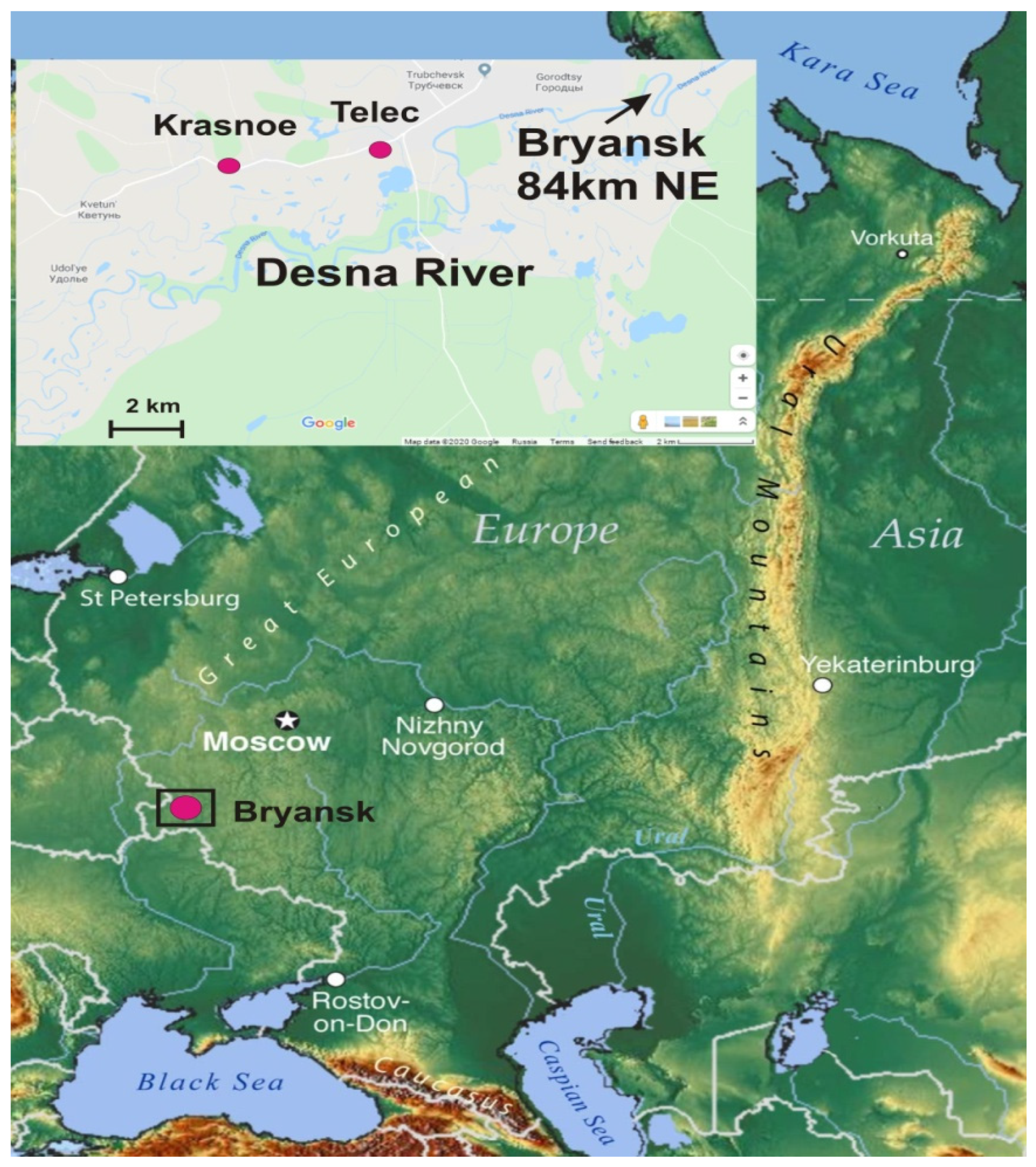
1. Introduction
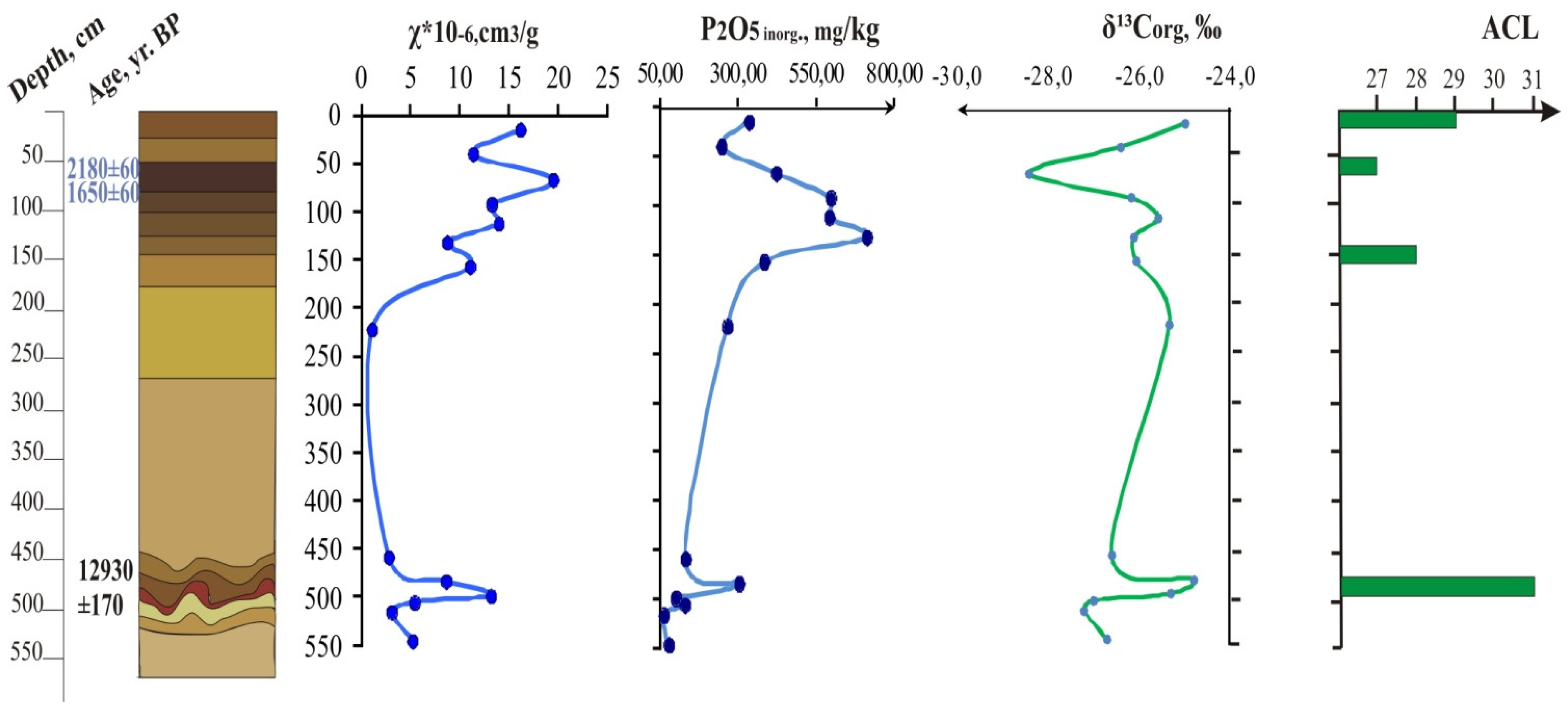
2. Materials and Methods
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Age, Years BP | Horizon, Depth, cm | Corg., % * | P2O5 org, mg/kg * | δ13Corg, ‰ * | δ13Ccarb, ‰ * | n-alkane, tot., μg/g d.w. ** |
|---|---|---|---|---|---|---|---|
| 1. Quarry, v. Telec, micro- elevation | Ap, 0−30 | 1.2 | 115.0 | −26.1 | non-carb. | 1.03 | |
| AE, 30−35 | 0.4 | 17.8 | −25.4 | non-carb. | 0.19 | ||
| EB, 35−46 | 0.2 | 18.4 | −24.7 | non-carb. | 0.41 | ||
| BC, 46−65 | 0.2 | 10.6 | −24.7 | non-carb. | - | ||
| C, 65−85 | 0.2 | 0.0 | −24.1 | non-carb. | - | ||
| 6690 ± 110 | Ab, 85−99 | 0.4 | 97.2 | −25.4 | −13.5 | - | |
| C1, 99−120 | 0.3 | 83.9 | −25.3 | −10.0 | - | ||
| C2, 120−.. | 0.2 | 38.9 | −25.7 | −8.3 | - | ||
| C3, 100−250 | 0.5 | 29.9 | −23.1 | non-carb. | |||
| 16,500 ± 230 | Bb, 250−275 | 0.3 | 0.0 | −26.5 | −22.3 | 0.00 | |
| BCb, 275−300 | 0.4 | 68.0 | −25.6 | −22.4 | 0.23 | ||
| ORTZ1, 350−420 | 0.3 | 44.5 | −26.5 | −22.0 | - | ||
| Eg, 770−970 | 0.1 | 0.0 | −26.6 | non-carb. | - | ||
| C4, 970−1000 | 0.1 | 0.0 | −26.7 | non-carb. | - | ||
| C5, 1000−1300 | 0.2 | 0.0 | −25.7 | non-carb. | - | ||
| 2. Quarry, v.Krasnoe, micro- depression | Ap, 0−30 | 1.8 | 178.7 | −25.0 | non-carb. | 0.73 | |
| AE, 30−50 | 1.5 | 311.9 | −26.4 | non-carb. | 0.10 | ||
| 2180 ± 60, 1650 ± 60 | *** Ab, 50−85 | 2.3 | 635.8 | −28.4 | non-carb. | 0.15 | |
| AEb, 85−100 | 1.3 | 389.8 | −26.2 | non-carb. | - | ||
| Eb, 100−125 | 0.4 | 125.4 | −25.6 | non-carb. | - | ||
| EBb, 124−140 | 0.6 | 0.0 | −26.1 | non-carb. | - | ||
| Bb, 140−175 | 0.3 | 0.0 | −26.1 | non-carb. | 1.02 | ||
| C1, 175−270 | 0.2 | 0.0 | −25.3 | non-carb. | - | ||
| C2,fe,1, 445–470 | 0.2 | 0.0 | −26.6 | non-carb. | - | ||
| 12,930 ± 170 | Bb2, 470–495 | 0.4 | 106.3 | −24.8 | −8.8 | 0.80 | |
| BCfe,b2, 495−500 | 0.1 | 0.0 | −25.3 | non-carb. | - | ||
| Eg, 500−510 | 0.1 | 0.0 | −27.0 | non-carb. | - | ||
| BCfe,b3 510−520 | 0.1 | 7.8 | −27.2 | non-carb. | - | ||
| C3 520−570 | 0.1 | 2.6 | −26.7 | non-carb. | - | ||
| 3. Modern soil, depression * | A, 10−45 | 3.4 | 271.2 | −32.7 | non-carb. | 1.14 | |
| *** Ab, 40−65 | 4.0 | 170.0 | −29.5 | non-carb. | 2.67 | ||
| AEg, 60−80 | 0.8 | 50.8 | −27.5 | non-carb. | 2.15 | ||
| Eg, 80−110 | 0.3 | 0.0 | −27.0 | non-carb. | - |
| Sample, Depth, cm | Relief Position | Age, Years cal. BP | n-alkane tot.(C17-36), μg/kg d.w. | Cmax | OEP1 | ACL27-33 | CPI27-33 | (nC31 + nC33)/ (nC27 + nC29) | Paq |
|---|---|---|---|---|---|---|---|---|---|
| 1. Quarry Telec | |||||||||
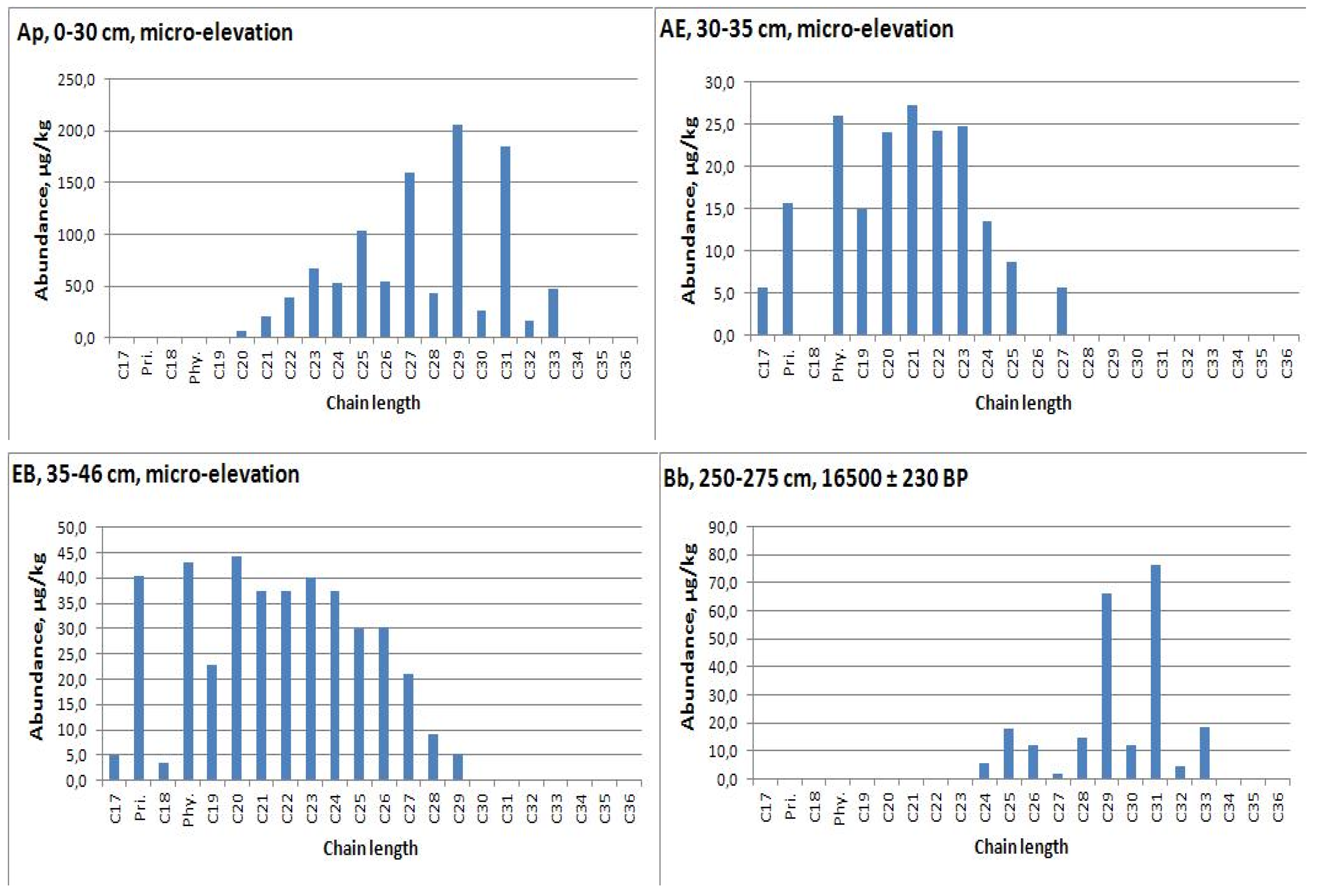
| Ap, 0−30 | micro-elevation | 1027.9 | C29 | 4.3 | 29 | 3.5 | 0.63 | 0.30 | |
| AE, 30−35 | 190.2 | C21 | - | 27 | - | 0 | 1.0 | ||
| EB, 35−46 | 406.2 | C20 | 0.7 | 27 | 1.4 | 0 | 0.93 | ||
| BCb, 275−300 | 16,500 ± 230 | 230.5 | C31 | 3.8 | 30 | 2.6 | 1.39 | 0.11 | |
| 2. Quarry Krasnoe | |||||||||
| Ap, 0−30 | micro-depression | 730.0 | C29 | 4.3 | 29 | 3.5 | 0.40 | 0.44 | |
| AE, 30−50 | 97.6 | C22 | 1.00 | ||||||
| Ab, 50−85 | 2180 ± 60, 1650 ± 60 | 151.3 | C21 | - | 27 | - | 0 | 1.00 | |
| Bb, 140−175 | 1020.3 | C21 | 0.8 | 28 | 1.4 | 0.08 | 0.81 | ||
| Bb2, 470−495 | 12,930 ± 170 | 804.3 | C31 | 5.7 | 31 | 4.0 | 1.95 | 0.10 | |
| 3. Modern soil, depression | |||||||||
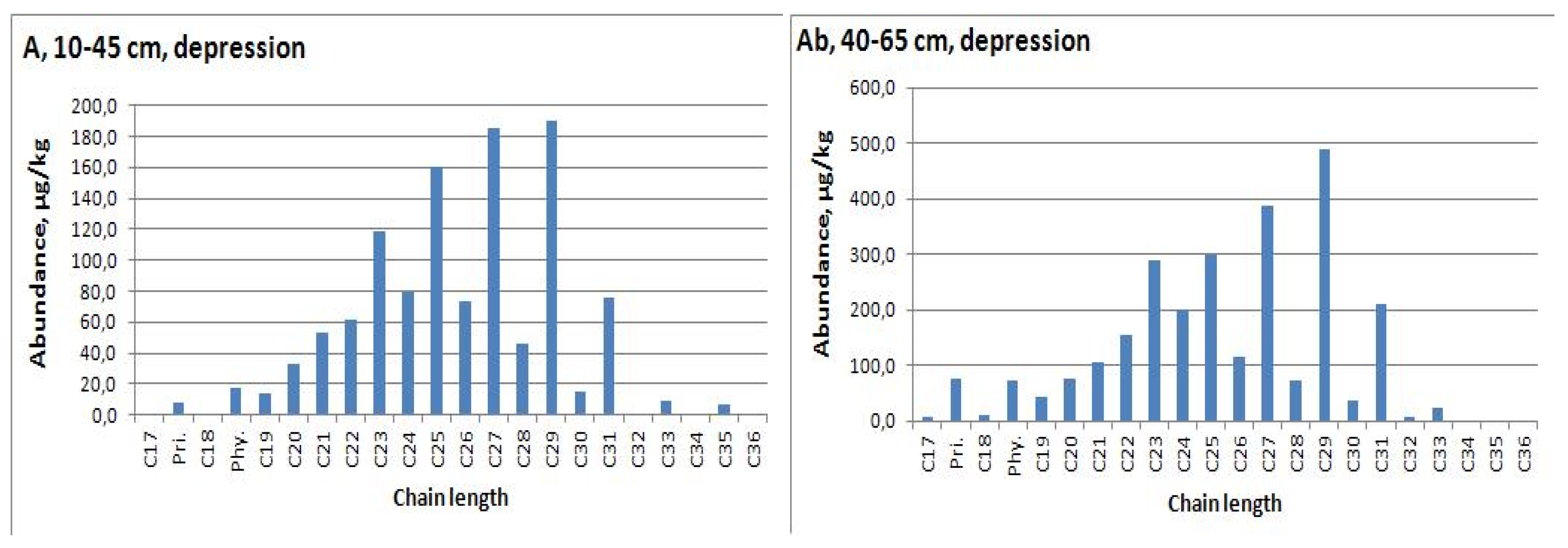
| A, 10−45 | depression | 1142.6 | C29 | 3.4 | 29 | 3.7 | 0.23 | 0.51 | |
| Ab, 40−65 | 2668.6 | C29 | 4.8 | 29 | 4.8 | 0.27 | 0.46 | ||
| AEg, 60−80 | 2153.9 | C25 | 0.9 | 28 | 1.0 | 0.14 | 0.71 | ||
References
- Bolihovskaya, N.S. Evolution of the Loess-Soil Formation of Northern Eurasia; MSU Press: Moscow, Russia, 1995. [Google Scholar]
- Velichko, A.A.; Morozova, T.D.; Nechaev, V.P.; Porozhnyakova, O.M. Paleocryogenesis, Soil Cover and Agriculture; Nauka: Moscow, Russia, 1996. [Google Scholar]
- Kovaleva, N.O.; Stolpnikova, E.M.; Kovalev, I.V. Paleoecological reconstruction for the Podesenie region on the Late Pleistocene-Holocene boundary (according to carbon isotopic composition of the soil). Povolzhskiy J. Eco. 2013, 4, 402–413. [Google Scholar]
- Morozova, T.D. Development of the Soil Cover of Europe in the Late Pleistocene; Nauka: Moscow, Russia, 1981. [Google Scholar]
- Gugalinskaya, L.A. Soil Formation and Cryogenesis in the Center of the East European Plain in late Pleistocene; ONTI NTsBI AN SSSR: Pushchino, Russia, 1982. [Google Scholar]
- Makeev, A.O. Pedogenic alteration of aeolian sediments in the upper loess mantles of the Russian Plain. Quat. Int. 2009, 209, 79–94. [Google Scholar] [CrossRef]
- Sedov, S.N.; Khokhlova, O.S.; Sinitsyn, A.A.; Korkka, M.A.; Rusakov, A.V.; Ortega, B.; Solleiro, E.; Rozanova, M.S.; Kuznetsova, A.M.; Kazdym, A.A. Late Pleistocene paleosol sequences as an instrument for the local paleographic reconstruction of the Kostenki 14 key section (Voronezh oblast) as an example. Eurasian Soil Sci. 2010, 43, 876–892. [Google Scholar] [CrossRef]
- Sycheva, S.; Khokhlova, O.; Pushkina, P.; Ukrainsky, P. Interrelations of the Bryansk paleosol (end of MIS 3) with the Holocene surface soils in micro-depressions of the central forest-steppe within the Russian Upland. Catena 2019, 172, 619–623. [Google Scholar] [CrossRef]
- Gennadiev, A.N.; Zavgorodnyaya, Y.A.; Pikovskii, Y.I.; Smirnova, M.A. Alkanes as components of soil hydrocarbon status: Behavior and indication significance. Eurasian Soil Sci. 2018, 51, 32–41. [Google Scholar] [CrossRef]
- Anokhina, N.A.; Demin, V.V.; Zavgorodnyaya, Y.A. Compositions of n-alkanes and n-methyl ketones in soils of the forest-park zone of Moscow. Eurasian Soil Sci. 2018, 51, 637–646. [Google Scholar] [CrossRef]
- Velichko, A.A. Natural Process in the Pleistocene; Nauka: Moscow, Russia, 1973. [Google Scholar]
- Alifanov, V.M.; Gugalinskaya, L.A.; Ovchinnikov, A.Y. Paleocriogenesis and a Variety of Soils on the Center of East European Plain; GEOS: Moscow, Russia, 2010. [Google Scholar]
- Zech, M.; Rass, S.; Buggle, B.; Loscher, M.; Zoller, L. Reconsruction of the Late Quaternary paleoenviroments of Nussloch loess paleosol sequence, Germany, using n-alkane biomarkers. Quat. Res. 2012, 78, 226–235. [Google Scholar] [CrossRef]
- Zech, M.; Buggle, B.; Leiber, K.; Marcovic, S.; Glaser, B.; Hambach, U.; Huwe, B.; Stevens, T.; Sumegi, P.; Wiesenberg, G.; et al. Reconstructing Quaternary vegetation in the Carpathian Basin, SE Europe, using n-alkane biomarkers as molecular fossils: Problems and possible solutions, potential and limitations. E&G Quat. Sci. J. 2009, 58, 148–155. [Google Scholar]
- Zech, R.; Zech, M.; Marković, S.; Hambach, U.; Huang, Y. Humid glacials, arid interglacials? Critical thoughts on pedogenesis and paleoclimate based on multi-proxy analyses of the loess-paleosol sequence Crvenka, Nothern Serbia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 387, 165–175. [Google Scholar] [CrossRef]
- Stolpnikova, E.M.; Kovaleva, N.O.; Kovalev, I.V. The Carbon Isotope Composition of Organic Matter and the Age od Paleosols from Wurm Glaciation Interstadials to Holocene (Bryansk region, Russia). IOP Conf. Series EES 2018, 107, 1–6. [Google Scholar] [CrossRef]
- Kogel-Knaber, I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol. Biochem. 2002, 34, 139–162. [Google Scholar] [CrossRef]
- Avato, P.; Bianchi, G.; Mariani, G. Epicuticular waxes of Sorghum and some compositional changes with plant age. Phytochemistry 1984, 23, 2843–2846. [Google Scholar] [CrossRef]
- Bull, I.D.; van Bergen, P.F.; Nott, C.J.; Poulton, P.R.; Evershed, R.P. Organic geochemical studies of soils from the Rothamsted classical experiments. The fate of lipids in different long-term experiments. Org. Geochem. 2000, 31, 389–408. [Google Scholar] [CrossRef]
- Jetter, R.; Kunst, L.; Samuels, A.L. Composition of plant cuticular waxes//Biology of plant cuticle; Wiley-Blackwell: Hoboken, NJ, USA, 2006; Volume 23, pp. 145–181. [Google Scholar]
- Luo, P.; Peng, P.; Lu, H.Y.; Zheng, Z.; Wang, X. Latitudinal variations of CPI values of long-chain n-alkanes in surface soil: Evidence for CPI as a proxy of aridity. Sci. China, Earth Sci. 2012, 55, 1134–1146. [Google Scholar] [CrossRef]
- Bush, R.T.; McInerney, F.A. Leaf wax n-alkane distributions in and across modern plants: Implications to paleoecology and chemotaxonomy. Geochim. Cosmochim. Acta 2013, 117, 161–179. [Google Scholar] [CrossRef]
- Zech, M.; Pedentchouk, N.; Markovic, S.B.; Glaser, B. Effect of leaf litter degradation and seasonality on D/H isotope ratios of n-alkane biomarkers. Geochim. Cosmochim. Acta 2011, 75, 4917–4928. [Google Scholar] [CrossRef]
- Peters, K.E.; Walter, C.C.; Moldowan, J.M. The n-alkane concentrations in buds and leaves of browsed broad leaf trees. J. Agric. Sci. 2000, 135, 311–320. [Google Scholar] [CrossRef]
- Loez-Dias, V.; Blanco, C.D.; Bechtel, A. Different source of n-alkanes and n-alkan-2 ones in a 6000 cal. yr. BP Sphagnum-rich temperate peat bog. Org. Geochem. 2013, 57, 7–10. [Google Scholar]
- Van Beilen, J.B.; Neuenschwander, M.; Smits, T.H.M.; Roth, C.; Balada, S.B.; Witholt, B. Rubredoxins involved in alkane oxidation. J. Bacteriol. 2003, 184, 427–440. [Google Scholar] [CrossRef]
- Walker, J.D.; Cooney, J.J. Aliphatic hydrocarbons of Cladosporium resinae cultured on glucose, glutamic acid, and hydrocarbons. Appl. Microbiol. 1973, 26, 705–708. [Google Scholar] [CrossRef]
- Bagaeva, T.V.; Zinurova, E.E. Comparative characterization of extracellular and intracellular hydrocarbons of Clostridium pasteurianum. Biochemistry (Moscow) 2004, 69, 427–428. [Google Scholar] [CrossRef] [PubMed]
- Bagaeva, T.V.; Zolotukhina, L.M. The formation of hydrocarbons by sulfate reducing bacteria grown under chemolithoheterotrophic conditions. Microbiology (Moscow) 1994, 63, 993–995. [Google Scholar]
- Bagaeva, T.V. The ability of sulfate-reducing bacteria of various taxonomic groups to synthesize extracellular hydrocarbons. Microbiology (Moscow) 1997, 66, 796–799. [Google Scholar]
- Bagaeva, T.V. Effect of the composition of the gaseous phase on the formation of hydrocarbons in Desulfovibrio desulfuricans. Appl. Biochem. Microbiol. (Moscow) 2000, 36, 195–198. [Google Scholar]
- Koser, J.; Volkman, J.K.; Rullkotter, J.; Scholzbottcher, B.M.; Rethmeier, J.; Fischer, U. Monomethyl-branched, dimethyl-branched, and trimethyl-branched alkanes in cultures of the filamentous cyanobacterium Calothrix scopulorum. Org. Geochem. 1999, 30, 1367–1379. [Google Scholar] [CrossRef]
- Fisher, D.J.; Holloway, P.J.; Richmond, D.V. Fatty acid and hydrocarbon constituents of the surface and wall lipids of some fungals pores. Microbiology 1972, 72, 71–78. [Google Scholar]
- Joes, J.G. Studies on lipids of soil microorganisms with particular reference to hydrocarbons. Microbiology 1969, 59, 145–152. [Google Scholar]
- Eglington, G.; Logan, G.A. Molecular preservation. Phill. Trans. Roy. Soc. London 1991, 333, 315–328. [Google Scholar]
- Wentzel, A.; Ellingsen, T.E.; Kotlar, H.K.; Zotchev, S.B.; Throne-Holst, M. Bacterial metabolism of long-chain n-alkanes. Appl. Microbiol. Biotechnol. 2007, 76, 1209–1221. [Google Scholar] [CrossRef]
- Handley, L.; Pearson, P.N.; McMillan, I.K.; Pancost, R.D. Large terrestrial and marine carbon and hydrocarbon isotope excursions in a new Paleocene/Eocene boundary section from Tanzania. Earth Planet. Sci. Lett. 2008, 275, 17–25. [Google Scholar] [CrossRef]
- Singh, S.N. Microbial Degradation of Xenobiotics; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Quenea, K.; Derenne, S.; Largeau, C.; Rumpel, C.; Mariotti, A. Variation in lipid relative abundance and composition among different particle size fractions of a forest soil. Org. Geochem. 2004, 35, 1355–1370. [Google Scholar] [CrossRef]
- Jansen, B.; Hausmann, N.S.; Tonneijck, F.H.; Verstraten, J.M.; de Vooght, P. Characteristic straight-chain lipid ratios as a quick method to assess past forest—paramo transitions in the Ecuadorian Andes. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 262, 129–139. [Google Scholar] [CrossRef]
- Duchko, M.A.; Gulaya, E.V.; Serebrenikova, O.V.; Strelnikova, E.B.; Preis, Y.I. Distribution of n-alkanes, steroids and triterpenoids in peat and plants of swamp “Tyomnoe”. Proc. Tomsk Polytech. Univ. 2013, 323, 40–44. [Google Scholar]
- Huang, J.; Lockheart, M.J.; Collister, J.W.; Eglington, G. Molecular and isotopic biogeochemistry of the Miocene Clarkia Formation: Hydrocarbons and alcohols. Org. Geochem. 1995, 23, 785–801. [Google Scholar] [CrossRef]
- Otto, A.; Simoneit, B.R.T.; Rember, W.C. Conifer and angiosperm biomarkers in clay sediments and fossil plants from the Miocene Clarkia Formation, Idaho, USA. Org. Geochem. 2005, 36, 907–922. [Google Scholar] [CrossRef]
- Aichner, B.; Ott, F.; Slowinski, M.; Noryskievicz, A.M.; Brauer, A.; Sashce, D. Leaf wax n-alkane distributions record ecological changes during Younger Dryas at Trzechowskie paleolake (northern Poland) without temporal delay. Clim. Past 2018, 14, 1607–1624. [Google Scholar] [CrossRef]
- Li, G.; Li, L.; Tarozo, R.; Longo, W.M.; Wang, K.J.; Dong, H.; Huang, Y. Microbial production of long-chain n-alkanes: Implication for interpreting sedimentary leaf wax signals. Org. Geochem. 2018, 115, 24–31. [Google Scholar] [CrossRef]
- McKirdly, D.M.; Thorpe, C.S.; Haynes, D.E.; Grice, K.; Krull, E.S.; Halverson, G.P.; Webster, L.J. The biogeochemical evolution of the Coorong during the mid- to late Holocene: An elemental, isotopic and biomarker perspective. Org. Geochem. 2010, 41, 96–110. [Google Scholar] [CrossRef]
- Vogts, A.; Badewien, T.; Rullkötter, J.; Schefuß, E. Near-constant apparent hydrogen isotope fractionation between leaf wax n-alkanes and precipitation in tropical regions: Evidence from a marine sediment transect off SW Africa. Org. Geochem. 2016, 96, 18–27. [Google Scholar] [CrossRef]
- Bliedtner, M.; Zech, R.; Kühn, P.; Schneider, B.; Zielhofer, C.; von Suchodoletz, H. The potential of leaf wax biomarkers from fluvial soil-sediment sequences for paleovegetation reconstructions—Apper Alazani River, central southern Greater Caucasus (Geogia). Quat. Sci. Rev. 2018, 196, 62–79. [Google Scholar] [CrossRef]
- Obreht, I.; Zeeden, C.; Hambach, U.; Veres, D.; Marković, S.B.; Lehmkuhl, F. A critical revaluation of paleoclimate proxy records from loess in the Carpatian Basin. Earth-Sci. Rev. 2019, 190, 498–520. [Google Scholar] [CrossRef]
- Tyuryukanov, A.N.; Bystritskaya, T.L. Opolya of Central Russia and Soils; Nauka: Moscow, Russia, 1971. [Google Scholar]
- Skripkin, V.; Kovalyukh, N. Recent developments in the procedures used at the SSCER laboratory for the preparation of Lithium Carbide. Radiocarbon 1998, 40, 211–214. [Google Scholar] [CrossRef]
- Ramsey, C.B. Bayesian analysis of radiocarbon dates. Radiocarbon 2009, 51, 337–360. [Google Scholar] [CrossRef]
- Reimer, P.J.; Bard, E.; Bayliss, A.; Beck, J.W.; Blackwell, P.G.; Bronk Ramsey, C.; Grootes, P.M.; Guilderson, T.P.; Haflidason, H.; Hajdas, I.; et al. IntCal13 and Marine13 Radiocarbon Age Calibration Curves 0–50,000 Years cal BP. Radiocarbon 2013, 55, 1869–1887. [Google Scholar] [CrossRef]
- Svendsen, J.I.; Alexanderson, H.; Astakhov, V.I.; Demidov, I.; Dowdeswell, J.A.; Funder, S.; Gataullin, V.; Henriksen, M.; Hjort, C.; Houmark-Nielsen, M.; et al. Late Quaternary ice sheet history of northern Eurasia. Quat. Sci. Rev. 2004, 23, 1229–1271. [Google Scholar] [CrossRef]
- Mol, J. Difinition of time slices. Landscape and climate change during the Last Glaciation in Europe; A review. In Evolution of the European Ecosystems during Pleistocene—Holocene transition (24–8 kyr BP); Markova, van Kolfschoten, T., Eds.; KMK Scientific Press Ltd.: Moscow, Russia, 2008; pp. 73–91. [Google Scholar]
- Kovaleva, N. Northern Tien-Shan paleosol sedimentary sequences as a record of major climatic events in the last 30,000 years. Revista Mexicana de Ciencias Geologicas 2004, 21-1, 71–78. [Google Scholar]
- Khokhlova, O.S.; Khokhlov, A.A.; Kuznetsova, A.M.; Stolpnikova, E.M.; Kovaleva, N.O.; Lyubin, V.P.; Belyaeva, E.V. Carbonate features in the uppermost layers of Quaternary deposits, Northern Armenia, and their significance for paleoenvironmental reconstruction. Quat. Int. 2016, 418, 94–104. [Google Scholar] [CrossRef]
- Markova, A.K.; Van Kolfschoten, T.; Bohncke, S.; Kosintsev, P.A.; Mol, J.; Puzachenko, A.Y. Evolution of the European Ecosystems during Pleistocene—Holocene transition (24–8 kyr BP); Scientific Press Ltd.: Moscow, Russia, 2008. [Google Scholar]
- Simakova, A.N.; Puzachenko, A.Y. Paleovegetation of Europe during the Bölling-Alleröd interstadial complex warming (12.4–10.9 ka BP). Polish Geol. Inst. Spec. Pap. 2005, 16, 116–122. [Google Scholar]








| Sample | Depth (cm) | Lab. Number | Material | Radiocarbon Age (yr) | Calendar Age (yr) |
|---|---|---|---|---|---|
| Ab, micro-depression | 50−85 | Ki-18775 | Humic acids | 1650 ± 60 | 1σ 1 334−530 BC 2σ 257−539 BC |
| Ab, micro-depression | 50−85 | Ki-17415 | Humic acids | 2180 ± 60 | 1σ 360−270 BC 2σ 390−90 BC |
| Ab, micro-elevation | 85−99 | Ki-18776 | Carbonates | 6690 ± 110 | 1σ 5706−5527 BC 2 5837−5392 BC |
| Bb2, Krasnoe | 470−495 | Ki-17413 | Humic acids | 12930 ± 170 | 1σ 14000−13200 BC 2σ 14200−12400 BC |
| Bb, Telec | 250−275 | Ki-17414 | Humic acids | 16500 ± 230 | 1σ 18150−17300 BC 2σ 18600−16900 BC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stolpnikova, E.; Kovaleva, N.; Kovalev, I. n-Alkane Distribution—A Paleovegetation Change Indicator during the Period from Late Glacial to Late Holocene on Russian Plain (Bryansk Region). Geosciences 2020, 10, 86. https://doi.org/10.3390/geosciences10030086
Stolpnikova E, Kovaleva N, Kovalev I. n-Alkane Distribution—A Paleovegetation Change Indicator during the Period from Late Glacial to Late Holocene on Russian Plain (Bryansk Region). Geosciences. 2020; 10(3):86. https://doi.org/10.3390/geosciences10030086
Chicago/Turabian StyleStolpnikova, Ekaterina, Natalia Kovaleva, and Ivan Kovalev. 2020. "n-Alkane Distribution—A Paleovegetation Change Indicator during the Period from Late Glacial to Late Holocene on Russian Plain (Bryansk Region)" Geosciences 10, no. 3: 86. https://doi.org/10.3390/geosciences10030086
APA StyleStolpnikova, E., Kovaleva, N., & Kovalev, I. (2020). n-Alkane Distribution—A Paleovegetation Change Indicator during the Period from Late Glacial to Late Holocene on Russian Plain (Bryansk Region). Geosciences, 10(3), 86. https://doi.org/10.3390/geosciences10030086