Transcriptional Control of Honey Bee (Apis mellifera) Major Royal Jelly Proteins by 20-Hydroxyecdysone

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Honey Bee Samples
2.2. RNA Extraction and cDNA Synthesis
2.3. Quantitative Real-Time PCR (qPCR)
2.4. Statistics
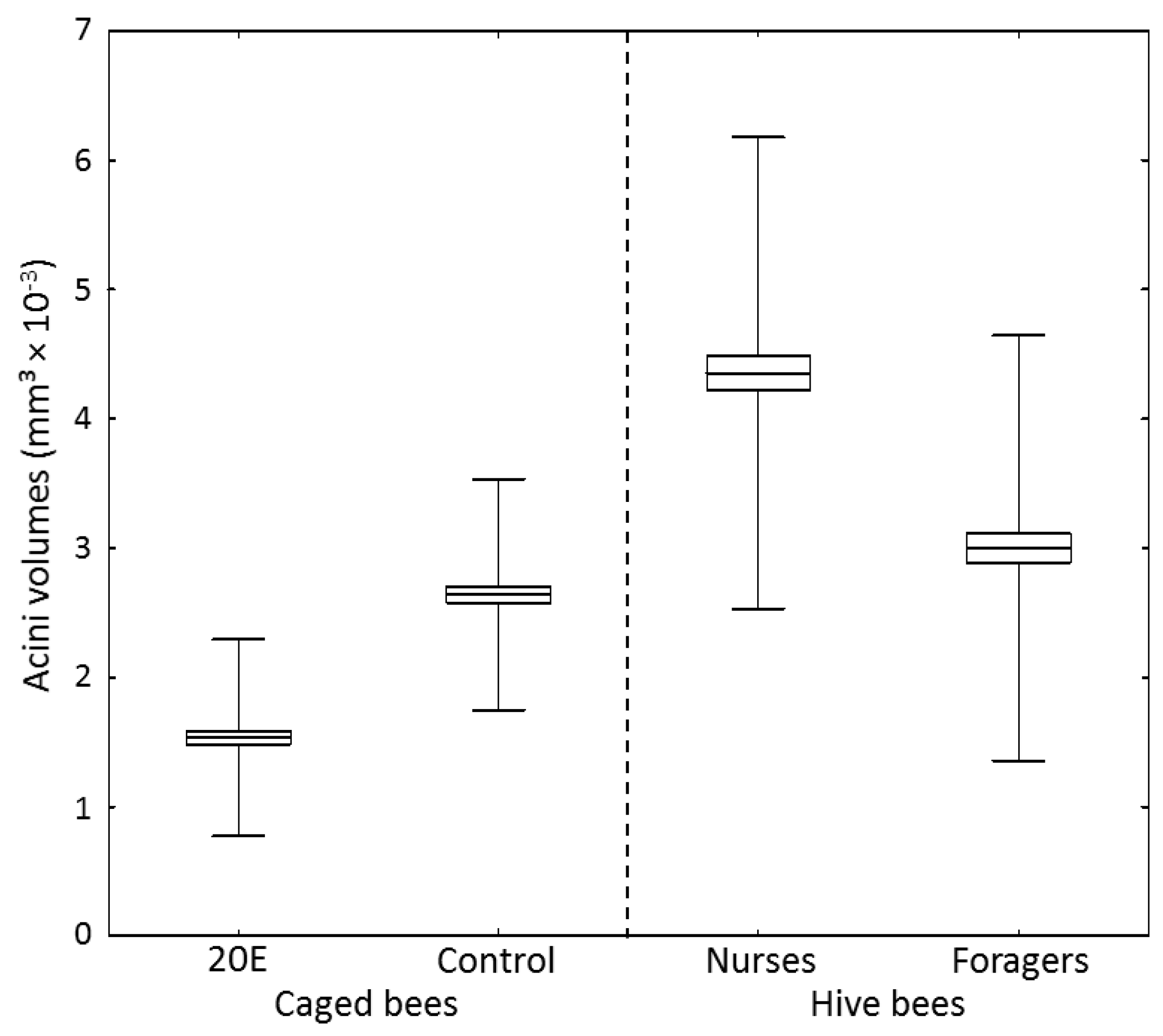
2.5. Acini Volumes
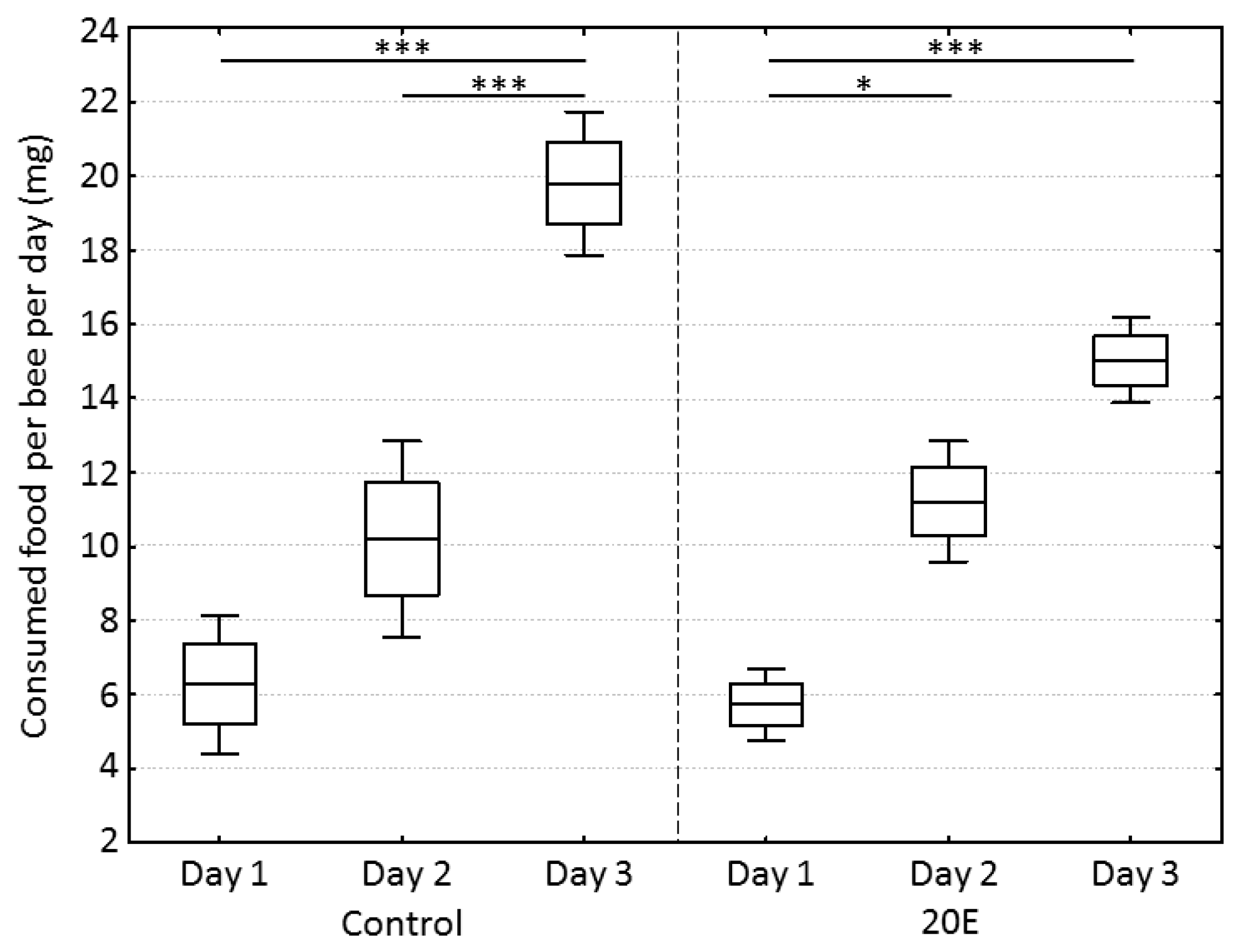
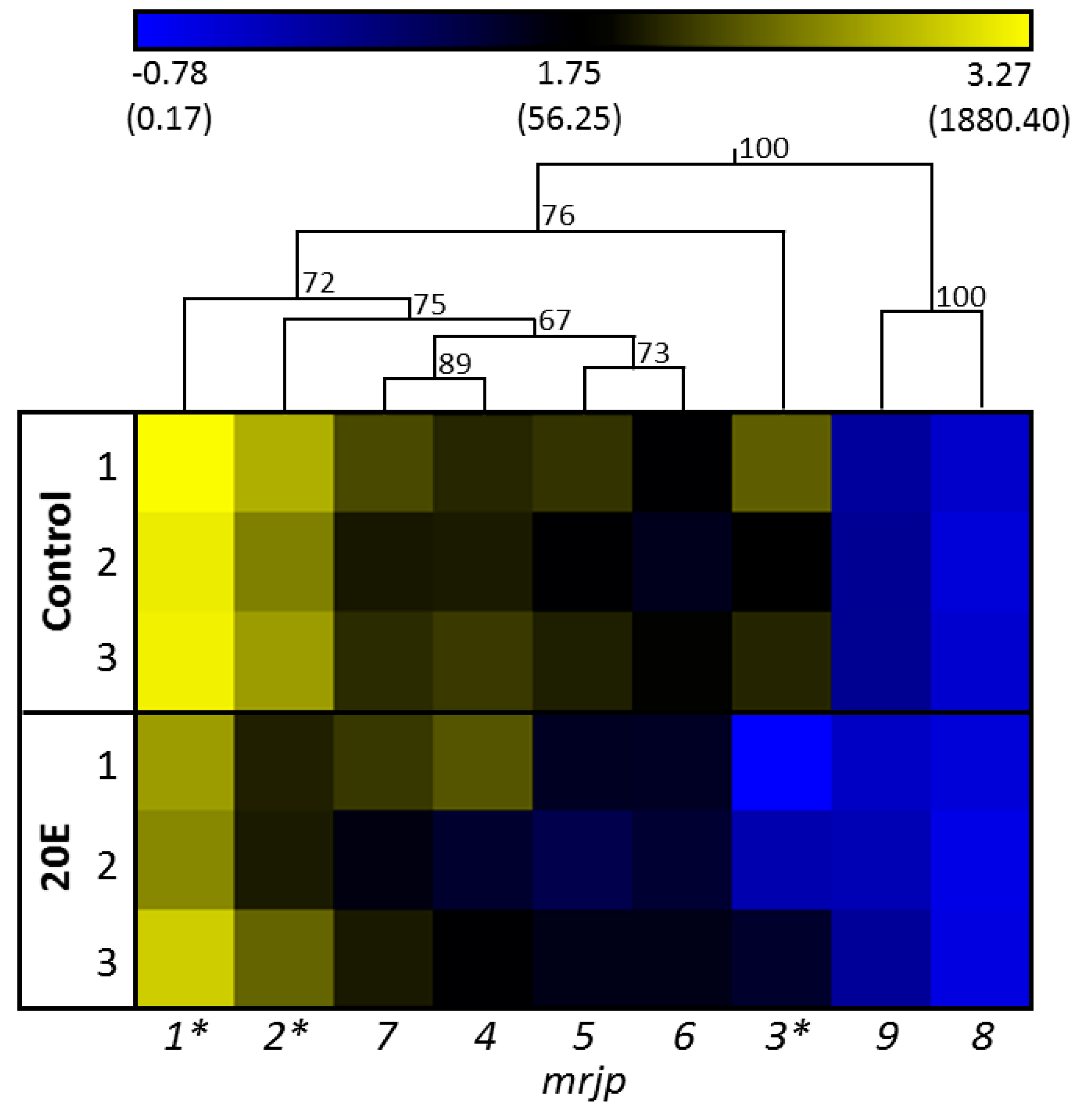
3. Results
4. Discussion
- (1)
- One way in which steroids act in target tissues is by binding to nuclear receptors that recognize thereupon specific DNA sequences, arranged as direct or inverted repeats (for review see Henrich & Brown [37]). Hitherto, ecdysone binds to a heterodimeric receptor subunit of the nuclear ecdysone receptor (EcR) and ultraspiracle (Usp) complex to mediate action [38,39]. Usp binding sites are present in mrjp promotor regions [40]. However, the EcR/Usp heterodimer usually activates gene expression in the presence of ecdysone, whereas the absence of ecdysone results in repression (for review see Henrich & Brown [37]). Therefore, a direct repression effect on mrjp gene expression, mediated by sole agonist activity at the nuclear ecdysone receptor complex, seems an unlikely explanation for the observed mrjp gene expression effects. However, the EcR/Usp dimer might also activate the expression of unknown target genes that in turn repress mrjps.
- (2)
- Besides the classical nuclear receptor action of ecdysone described above, a nongenomic action involving membrane receptors and second messengers (e.g., cyclic AMP) has repeatedly been suggested [41,42]. A first membrane ecdysone receptor was described in 2004 [42], subsequently leading to the identification of primarily G-protein-coupled receptors as being involved in steroid hormone signaling [43]. This membrane receptor-mediated ecdysone signaling can either repress or induce the expression of target genes, and thus might be an option as explanation for mrjp regulation.
- (3)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Butenandt, A.; Karlson, P. Über die Isolierung eines Metamorphose-Hormons der Insekten in kristallisierter Form. Z. Naturforsch. 1954, 9b, 389–391. [Google Scholar] [CrossRef]
- Karlson, P. Chemische Untersuchungen über die Metamorphosehormone der Insekten. Ann. Sci. Nat. Zool. Biol. Anim. 1956, 18, 125–137. [Google Scholar]
- Karlson, P.; Hoffmeister, H.; Hoppe, W.; Huber, R. Zur Chemie des Ecdysons. Justus Liebigs Ann. Chem. 1963, 662, 1–20. [Google Scholar] [CrossRef]
- Myers, E.M. The circadian control of eclosion. Chronobiol. Int. 2003, 20, 775–794. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, L.M.; Truman, J.W. Hormonal control of rates of metamorphic development in the tobacco hornworm Manduca sexta. Dev. Biol. 1983, 99, 103–114. [Google Scholar] [CrossRef]
- Amdam, G.V.; Page, R.E., Jr.; Fondrk, M.K.; Brent, C.S. Hormone response to bidirectional selection on social behavior. Evol. Dev. 2010, 12, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.Z.; Hartfelder, K.; Gentile Bitondi, M.M.; Simões, Z.L.P. Ecdysteroid titers in pupae of highly social bees relate to distinct modes of caste development. J. Insect Physiol. 2002, 48, 783–790. [Google Scholar] [CrossRef]
- Rachinsky, A.; Strambi, C.; Strambi, A.; Hartfelder, K. Caste and metamorphosis: Hemolymph titers of juvenile hormone and ecdysteroids in last instar honeybee larvae. Gen. Comp. Endocrinol. 1990, 79, 31–38. [Google Scholar] [CrossRef]
- Zufelato, M.S.; Bitondi, M.M.G.; Simões, Z.L.P.; Hartfelder, K. The juvenile hormone analog pyriproxyfen affects ecdysteroid-dependent cuticle melanization and shifts the pupal ecdysteroid peak in the honey bee (Apis mellifera). Arthropod Struct. Dev. 2000, 29, 111–119. [Google Scholar] [CrossRef]
- Hiruma, K.; Kaneko, Y. Hormonal regulation of insect metamorphosis with special reference to juvenile hormone biosynthesis. Curr. Top. Dev. Biol. 2013, 103, 73–100. [Google Scholar] [CrossRef] [PubMed]
- Barchuk, A.R.; Bitondi, M.M.G.; Simões, Z.L.P. Effects of juvenile hormone and ecdysone on the timing of vitellogenin appearance in hemolymph of queen and worker pupae of Apis mellifera. J. Insect Sci. 2002, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Engels, W. Occurrence and significance of vitellogenins in female castes of social hymenoptera. Am. Zool. 1974, 14, 1229–1237. [Google Scholar] [CrossRef]
- Bitondi, M.M.G.; Simoes, Z.L.P. The relationship between level of pollen in the diet, vitellogenin and juvenile hormone titres in Africanized Apis mellifera workers. J. Apic. Res. 1996, 35, 27–36. [Google Scholar] [CrossRef]
- Amdam, G.V.; Norberg, K.; Hagen, A.; Omholt, S.W. Social exploitation of vitellogenin. Proc. Natl. Acad. Sci. USA 2003, 100, 1799–1802. [Google Scholar] [CrossRef] [PubMed]
- Seehuus, S.-C.; Norberg, K.; Krekling, T.; Fondrk, K.; Amdam, G.V. Immunogold localization of vitellogenin in the ovaries, hypopharyngeal glands and head fat bodies of honeybee workers, Apis mellifera. J. Insect Sci. 2007, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Buttstedt, A.; Moritz, R.F.A.; Erler, S. Origin and function of the major royal jelly proteins of the honeybee (Apis mellifera) as members of the yellow gene family. Biol. Rev. 2014, 89, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Schmitzová, J.; Klaudiny, J.; Albert, S.; Schröder, W.; Schreckengost, W.; Hanes, J.; Júdová, J.; Simúth, J. A family of major royal jelly proteins of the honeybee Apis mellifera L. Cell. Mol. Life Sci. 1998, 54, 1020–1030. [Google Scholar] [CrossRef] [PubMed]
- Kratky, E. Morphologie und Physiologie der Drüsen in Kopf und Thorax der Honigbiene (Apis mellifica L.). Z. Wiss. Zool. 1931, 139, 119–200. [Google Scholar]
- Patel, N.G.; Haydak, M.H.; Gochnauer, T.A. Electrophoretic components of the proteins in honeybee larval food. Nature 1960, 186, 633–634. [Google Scholar] [CrossRef] [PubMed]
- Bíliková, K.; Mirgorodskaya, E.; Bukovská, G.; Gobom, J.; Lehrach, H.; Šimúth, J. Towards functional proteomics of minority component of honeybee royal jelly: The effect of post-translational modifications on the antimicrobial activity of apalbumin2. Proteomics 2009, 9, 2131–2138. [Google Scholar] [CrossRef] [PubMed]
- Vezeteu, T.V.; Bobiş, O.; Moritz, R.F.A.; Buttstedt, A. Food to some, poison to others-honeybee royal jelly and its growth inhibiting effect on European Foulbrood bacteria. MicrobiologyOpen 2017, 6, e00397. [Google Scholar] [CrossRef] [PubMed]
- Buttstedt, A.; Moritz, R.F.A.; Erler, S. More than royal food-Major royal jelly protein genes in sexuals and workers of the honeybee Apis mellifera. Front. Zool. 2013, 10, 72. [Google Scholar] [CrossRef] [PubMed]
- Buttstedt, A.; Mureşan, C.I.; Lilie, H.; Hause, G.; Ihling, C.H.; Schulze, S.H.; Pietzsch, M.; Moritz, R.F.A. How honeybees defy gravity with royal jelly to raise queens. Curr. Biol. 2018, 28, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Drapeau, M.D.; Albert, S.; Kucharski, R.; Prusko, C.; Maleszka, R. Evolution of the yellow/major royal jelly protein family and the emergence of social behavior in honey bees. Genome Res. 2006, 16, 1385–1394. [Google Scholar] [CrossRef] [PubMed]
- Helbing, S.; Lattorff, H.M.G.; Moritz, R.F.A.; Buttstedt, A. Comparative analyses of the major royal jelly protein gene cluster in three Apis species with long amplicon sequencing. DNA Res. 2017, 24, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Chan, Q.W.; Chan, M.Y.; Logan, M.; Fang, Y.; Higo, H.; Foster, L.J. Honey bee protein atlas at organ level resolution. Genome Res. 2013, 23, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- Ueno, T.; Nakaoka, T.; Takeuchi, H.; Kubo, T. Changes in the Gene Expression Profiles of the Hypopharyngeal Gland of Worker Honeybees in Association with Worker Behavior and Hormonal Factors. PLoS ONE 2015, 10, e0130206. [Google Scholar] [CrossRef] [PubMed]
- Mello, T.R.P.; Aleixo, A.C.; Pinheiro, D.G.; Nunes, F.M.F.; Bitondi, M.M.G.; Hartfelder, K.; Barchuk, A.R.; Simões, Z.L.P. Developmental regulation of ecdysone receptor (EcR) and EcR-controlled gene expression during pharate-adult development of honey bees (Apis mellifera). Front. Genet. 2014, 5, 445. [Google Scholar] [CrossRef] [PubMed]
- Hartfelder, K.; Bitondi, M.M.G.; Santana, W.C.; Simões, Z.L.P. Ecdysteroid titer and reproduction in queens and workers of the honey bee and of a stingless bee: Loss of ecdysteroid function at increasing levels of sociality? Insect Biochem. Mol. Biol. 2002, 32, 211–216. [Google Scholar] [CrossRef]
- Rharrabe, K.; Bouayad, N.; Sayah, F. Effects of ingested 20-hydroxyecdysone on development and midgut epithelial cells of Plodia interpunctella (Lepidoptera, Pyralidae). Pestic. Biochem. Physiol. 2009, 93, 112–119. [Google Scholar] [CrossRef]
- Boncristiani, H.; Li, J.; Evans, J.D.; Pettis, J.; Chen, Y. Scientific note on PCR inhibitors in the compound eyes of honey bees, Apis mellifera. Apidologie 2011, 42, 457–460. [Google Scholar] [CrossRef]
- Chromzynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Howe, E.; Holton, K.; Nair, S.; Schlauch, D.; Sinha, R.; Quackenbush, J. Mev: Multiexperiment viewer. In Biomedical Informatics for Cancer Research; Ochs, M.F., Casagrande, J.T., Davuluri, R.V., Eds.; Springer: New York, NY, USA, 2010; pp. 267–277. ISBN 978-1-4419-5712-2. [Google Scholar]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Omar, E.; Abd-Ella, A.A.; Khodairy, M.M.; Moosbeckhofer, R.; Crailsheim, K.; Brodschneider, R. Influence of different pollen diets on the development of hypopharyngeal glands and size of acid gland sacs in caged honey bees (Apis mellifera). Apidologie 2017, 48, 425–436. [Google Scholar] [CrossRef]
- Henrich, V.C.; Brown, N.E. Insect nuclear receptors: A developmental and coparative perspective. Insect Biochem. Mol. Biol. 1995, 25, 881–897. [Google Scholar] [CrossRef]
- Barchuk, A.R.; Maleszka, R.; Simões, Z.L.P. Apis mellifera ultraspiracle: CDNA sequence and rapid upregulation by juvenile hormone. Insect Mol. Biol. 2004, 13, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.P.; Segraves, W.A.; Oro, A.E.; McKeown, M.; Evans, R.M. Drosophila ultraspiracle modulates ecdysone receptor function via heterodimer formation. Cell 1992, 2, 63–72. [Google Scholar] [CrossRef]
- Malecová, B.; Ramser, J.; O’Brien, J.K.; Janitz, M.; Júdová, J.; Lehrach, H.; Simúth, J. Honeybee (Apis mellifera L.) mrjp gene family: Computational analysis of putative promoters and genomic structure of mrjp1, the gene coding for the most abundant protein of larval food. Gene 2003, 16, 165–175. [Google Scholar] [CrossRef]
- Applebaum, S.W.; Gilbert, L.I. Stimulation of adenyl cyclase in pupal wing epidermis by β-ecdysone. Dev. Biol. 1972, 27, 165–175. [Google Scholar] [CrossRef]
- Elmogy, M.; Iwami, M.; Sakurai, S. Presence of membrane ecdysone receptor in the anterior silk gland of the silkworm Bombyx mori. Eur. J. Biochem. 2004, 271, 3171–3179. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhao, W.-L.; Cai, M.-J.; Wang, J.-X.; Zhao, X.-F. G-protein-coupled receptor controls steroid hormone signaling in cell membrane. Sci. Rep. 2015, 5, 8675. [Google Scholar] [CrossRef] [PubMed]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of pollen nutrition on honey bee health: Do pollen quality and diversity matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef] [PubMed]
- Hrassnigg, N.; Crailsheim, K. Adaptation of hypopharyngeal gland development to the brood status of honeybee (Apis mellifera L.) colonies. J. Insect Physiol. 1998, 44, 929–939. [Google Scholar] [CrossRef]
- Knecht, D.; Kaatz, H.H. Patterns of larval food production by hypopharyngeal glands in adult worker honey bees. Apidologie 1990, 21, 457–468. [Google Scholar] [CrossRef]
- Huang, Z.-Y.; Otis, G.W.; Teal, P.E.A. Nature of brood signal activating the protein synthesis of hypopharyngeal gland in honey bees, Apis mellifera (Apidae: Hymenoptera). Apidologie 1989, 20, 455–464. [Google Scholar] [CrossRef]
- Hrassnigg, N.; Crailsheim, K. The influence of brood on the pollen consumption of worker bees (Apis mellifera L.). J. Insect Physiol. 1998, 44, 393–404. [Google Scholar] [CrossRef]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winkler, P.; Sieg, F.; Buttstedt, A. Transcriptional Control of Honey Bee (Apis mellifera) Major Royal Jelly Proteins by 20-Hydroxyecdysone. Insects 2018, 9, 122. https://doi.org/10.3390/insects9030122
Winkler P, Sieg F, Buttstedt A. Transcriptional Control of Honey Bee (Apis mellifera) Major Royal Jelly Proteins by 20-Hydroxyecdysone. Insects. 2018; 9(3):122. https://doi.org/10.3390/insects9030122
Chicago/Turabian StyleWinkler, Paul, Frank Sieg, and Anja Buttstedt. 2018. "Transcriptional Control of Honey Bee (Apis mellifera) Major Royal Jelly Proteins by 20-Hydroxyecdysone" Insects 9, no. 3: 122. https://doi.org/10.3390/insects9030122
APA StyleWinkler, P., Sieg, F., & Buttstedt, A. (2018). Transcriptional Control of Honey Bee (Apis mellifera) Major Royal Jelly Proteins by 20-Hydroxyecdysone. Insects, 9(3), 122. https://doi.org/10.3390/insects9030122