Predicting Geographic Distribution of Forensically Significant Blow Flies of Subfamily Chrysomyinae (Diptera: Calliphoridae) in Northern Thailand

 ,
,
Abstract
1. Introduction
2. Materials and Methods
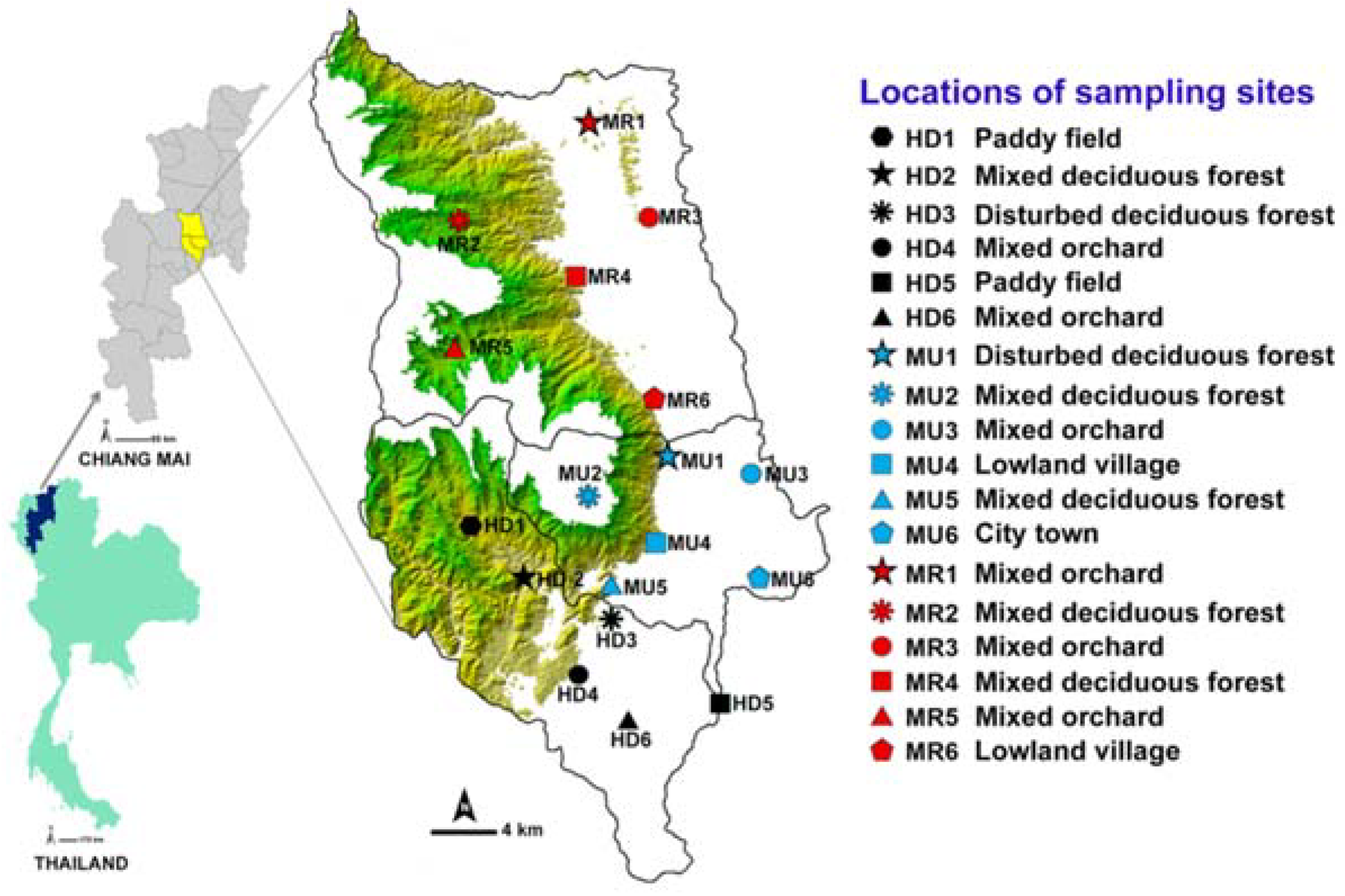
2.1. Study Areas
2.2. Fly Collection
2.3. Statistical Analysis
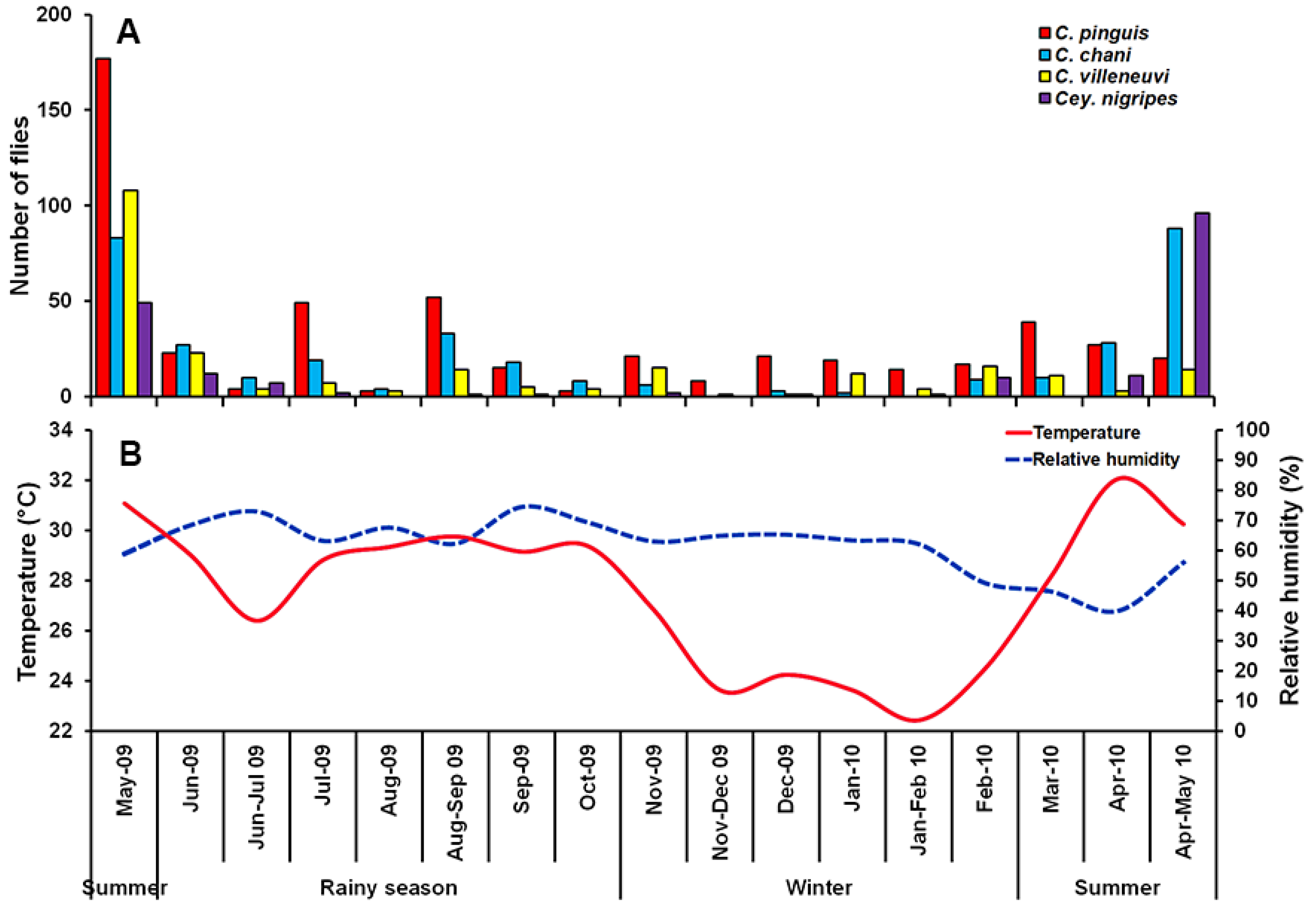
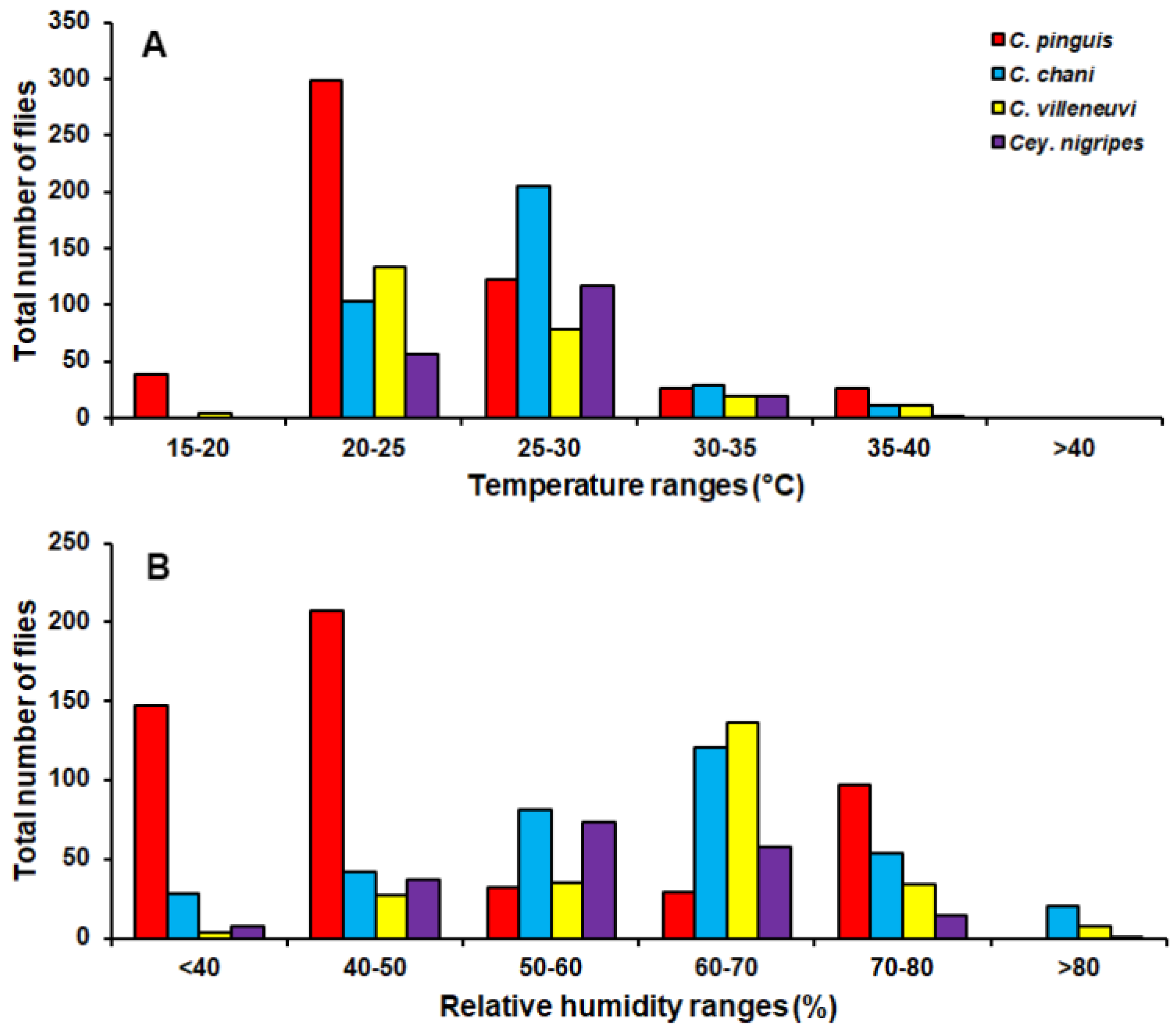
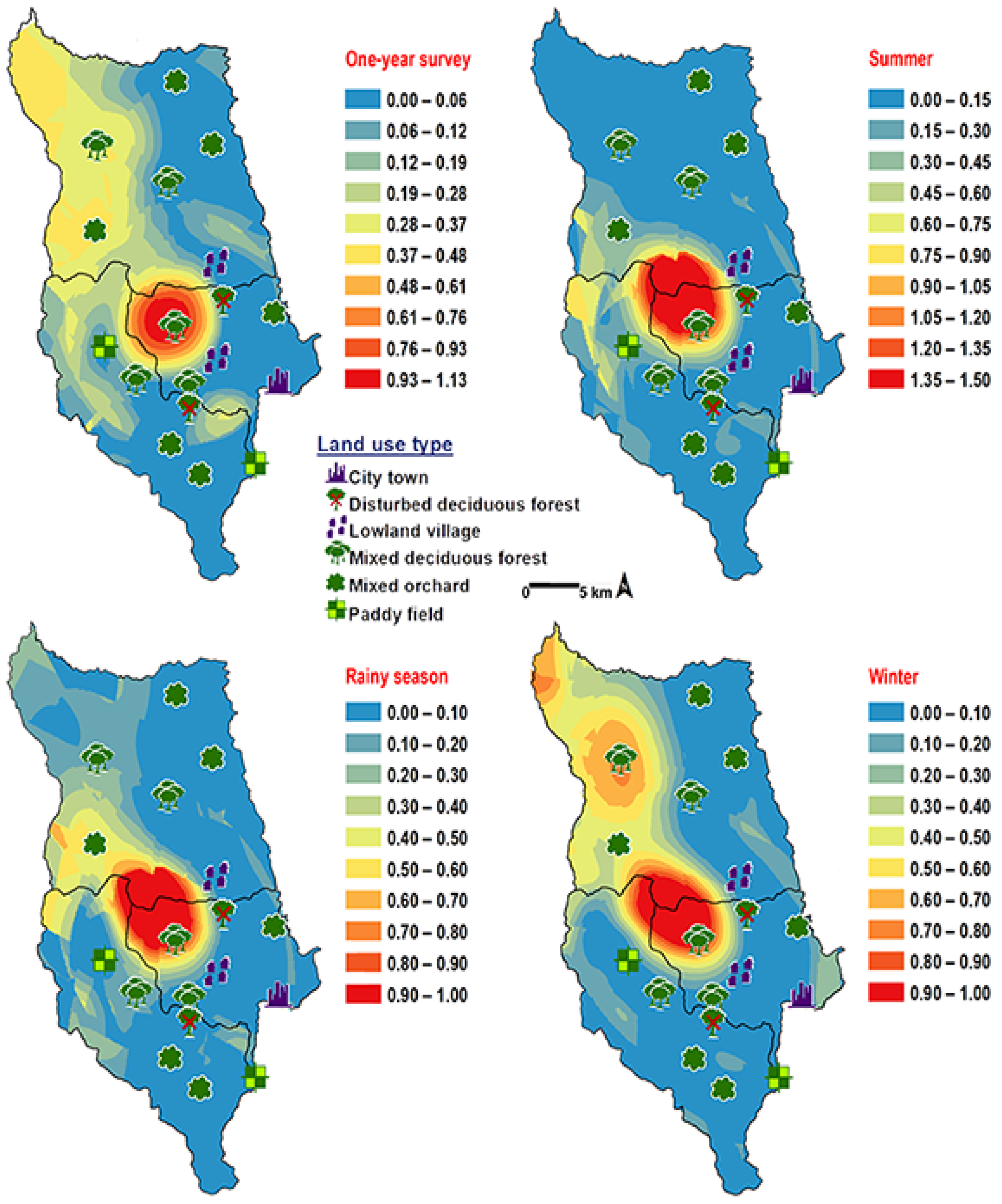
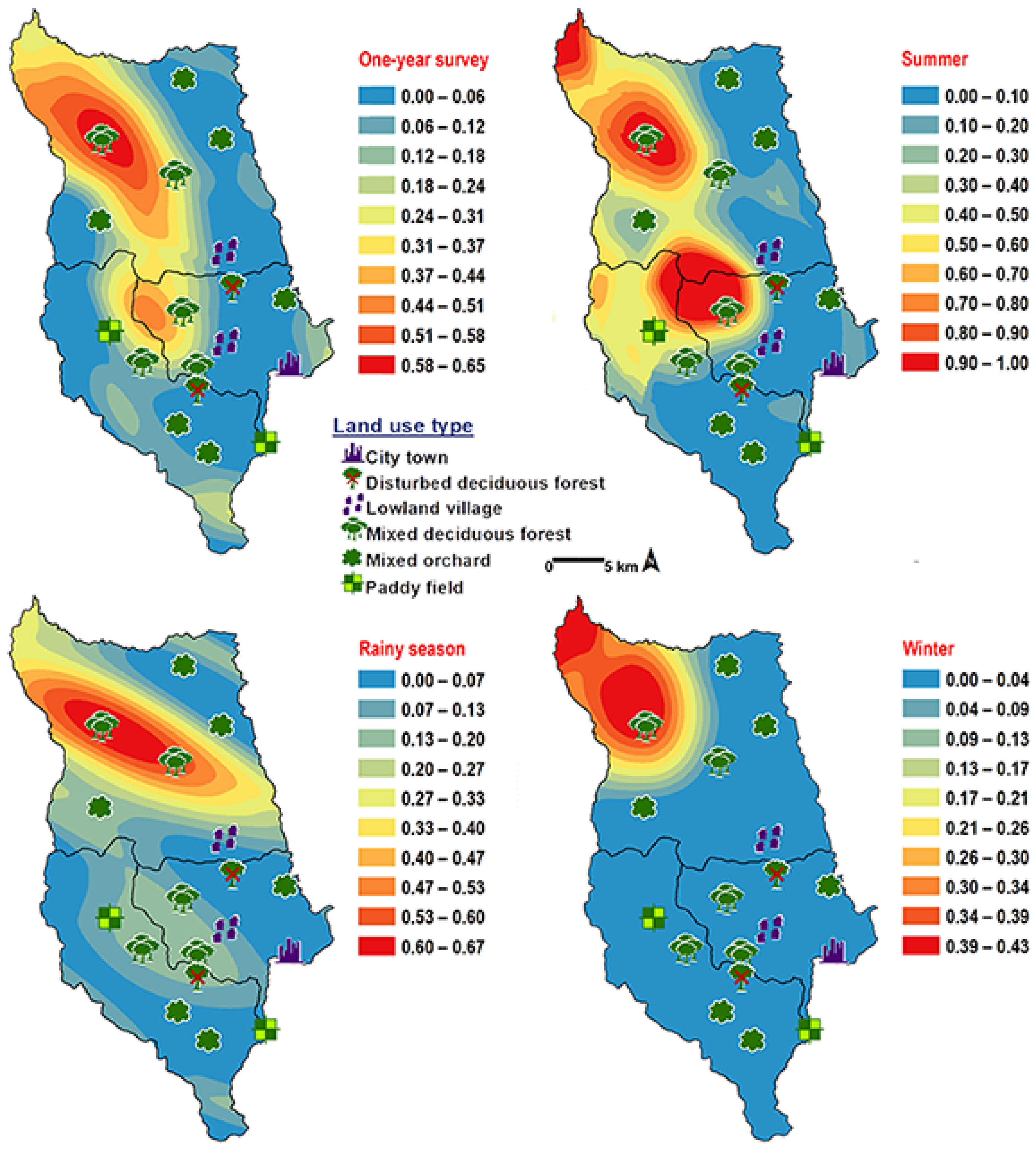
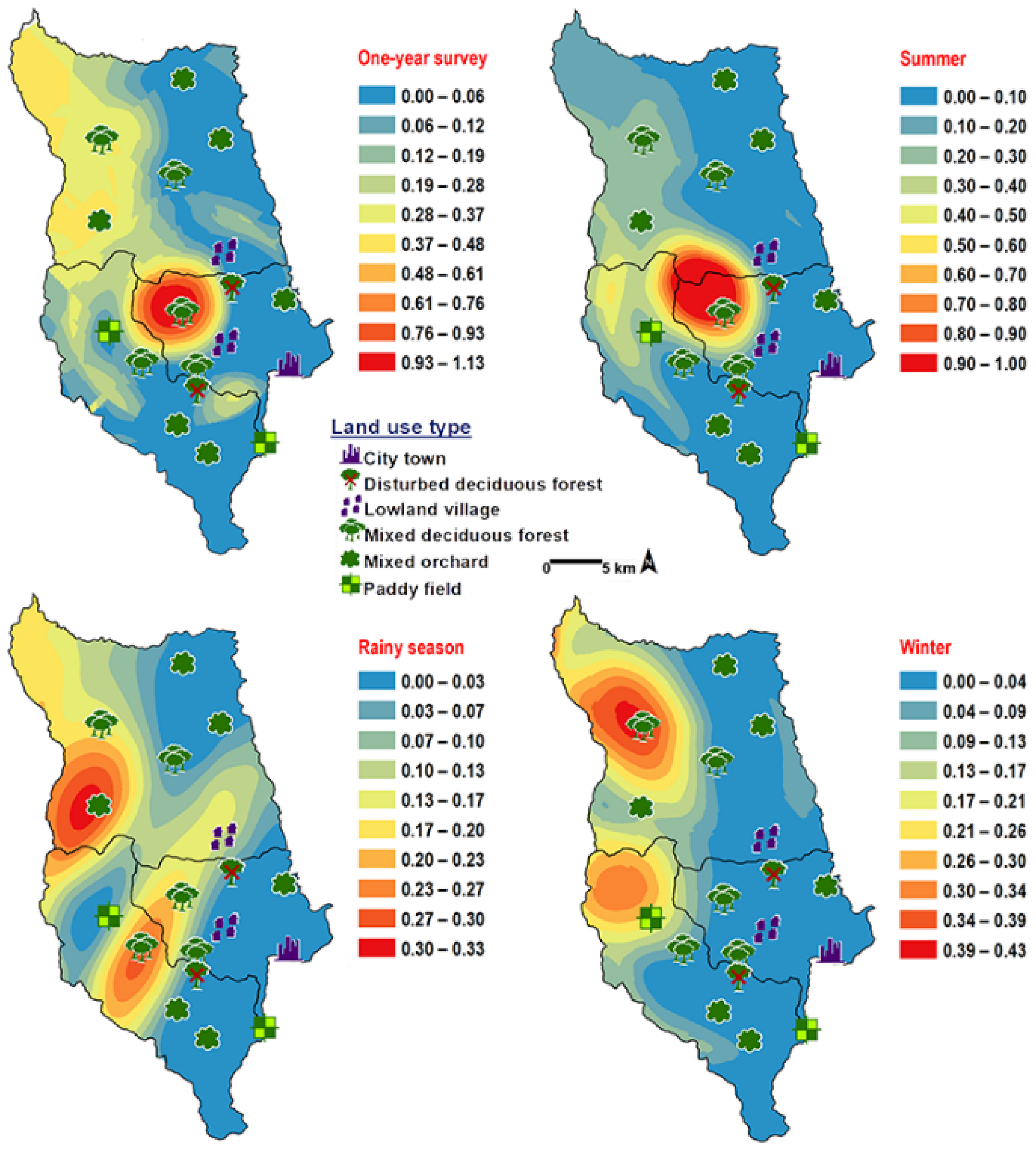
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tomberlin, J.K.; Crippen, T.L.; Tarone, A.M.; Chaudhury, M.F.B.; Singh, B.; Cammack, J.A.; Meisel, R.P. A review of bacterial interactions with blow flies (Diptera: Calliphoridae) of Medical, Veterinary, and Forensic importance. Ann. Entomol. Soc. Am. 2017, 110, 19–36. [Google Scholar] [CrossRef]
- Norris, K.R. The bionomics of blow flies. Annu. Rev. Entomol. 1965, 10, 47–68. [Google Scholar] [CrossRef]
- Brundage, A.; Bros, S.; Honda, J.Y. Seasonal and habitat abundance and distribution of some forensically important blow flies (Diptera: Calliphoridae) in Central California. Forensic Sci. Int. 2011, 212, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Amendt, J.; Campobasso, C.P.; Gaudry, E.; Reiter, C.; LeBlanc, H.N.; Hall, M.J. Best practice in Forensic Entomology—Standards and guidelines. Int. J. Legal Med. 2007, 121, 90–104. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.S.; Pepinelli, M.; de Almeida, E.C.; Ochoa-Quintero, J.M.; Roque, F.O. Blow flies from forest fragments embedded in different land uses: Implications for selecting indicators in Forensic Entomology. J. Forensic Sci. 2016, 61, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Sanit, S.; Sukontason, K.; Kurahashi, H.; Tomberlin, J.K.; Wannasan, A.; Kraisittipanit, R.; Sukontason, K.L. Morphology of immature stages of blow fly, Lucilia sinensis Aubertin (Diptera: Calliphoridae), a potential species of forensic importance. Acta Trop. 2017, 176, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Sontigun, N.; Sanit, S.; Wannasan, A.; Sukontason, K.; Amendt, J.; Yasanga, T.; Sukontason, K.L. Ultrastructure of male genitalia of blow flies (Diptera: Calliphoridae) of forensic importance. Acta Trop. 2018, 179, 61–80. [Google Scholar] [CrossRef] [PubMed]
- Zajac, B.K.; Sontigun, N.; Wannasan, A.; Verhoff, M.A.; Sukontason, K.; Amendt, J.; Zehner, R. Application of DNA barcoding for identifying forensically relevant Diptera from northern Thailand. Parasitol. Res. 2016, 115, 2307–2320. [Google Scholar] [CrossRef] [PubMed]
- Bunchu, N.; Thaipakdee, C.; Vitta, A.; Sanit, S.; Sukontason, K.; Sukontason, K.L. Morphology and developmental rate of the blow fly, Hemipyrellia ligurriens (Diptera: Calliphoridae): Forensic Entomology applications. J. Parasitol. Res. 2012. [Google Scholar] [CrossRef] [PubMed]
- Sukontason, K.; Piangjai, S.; Siriwattanarungsee, S.; Sukontason, K.L. Morphology and developmental rate of blowflies Chrysomya megacephala and Chrysomya rufifacies in Thailand: Application in Forensic Entomology. Parasitol. Res. 2008, 102, 1207–1216. [Google Scholar] [CrossRef] [PubMed]
- Vitta, A.; Pumidonming, W.; Tangchaisuriya, U.; Poodendean, C.; Nateeworanart, S. A preliminary study on insects associated with pig (Sus scrofa) carcasses in Phitsanulok, northern Thailand. Trop. Biomed. 2007, 24, 1–5. [Google Scholar]
- Moophayak, K.; Sukontason, K.L.; Ruankham, W.; Tomberlin, J.K.; Bunchu, N. Variation in the time of colonization of broiler carcasses by carrion flies in Nakhonsawan Province, Thailand. J. Med. Entomol. 2017, 54, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Moophayak, K.; Klong-klaew, T.; Sukontason, K.; Kurahashi, H.; Tomberlin, J.K.; Sukontason, K.L. Species composition of carrion blow flies in northern Thailand: Altitude appraisal. Rev. Inst. Med. Trop. Sao Paulo 2014, 56, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Sukontason, K.; Sukontason, K.L.; Piangjai, S.; Tippanun, J.; Lertthamnongtham, S.; Vogtsberger, R.C.; Olson, J.K. Survey of forensically-relevant fly species in Chiang Mai, Northern Thailand. J. Vector Ecol. 2003, 28, 135–138. [Google Scholar] [PubMed]
- Klong-klaew, T.; Sukontason, K.; Ngoen-klan, R.; Moophayak, K.; Irvine, K.N.; Kurahashi, H.; Prangkio, C.; Sanit, S.; Sukontason, K.L. Impact of abiotic factor changes in blowfly, Achoetandrus rufifacies (Diptera: Calliphoridae), in northern Thailand. Parasitol. Res. 2014, 113, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Ngoen-klan, R.; Moophayak, K.; Klong-klaew, T.; Irvine, K.N.; Sukontason, K.L.; Prangkio, C.; Somboon, P.; Sukontason, K. Do climatic and physical factors affect populations of the blow fly Chrysomya megacephala and house fly Musca domestica? Parasitol. Res. 2011, 109, 1279–1292. [Google Scholar] [CrossRef] [PubMed]
- Toukairin, Y.; Arai, T.; Hoshi, T.; Oliva Trejo, J.A.; Nogami, M. The geographical distribution of fly larvae on corpses in Saitama Prefecture in Japan during the summer season. Leg. Med. 2017, 24, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, R.; Nazni, W.A.; Tan, T.C.; Lee, H.L.; Azirun, M.S. Review of forensically important entomological specimens collected from human cadavers in Malaysia (2005–2010). J. Forensic Leg. Med. 2013, 20, 480–482. [Google Scholar] [CrossRef] [PubMed]
- Sukontason, K.; Narongchai, P.; Kanchai, C.; Vichairat, K.; Sribanditmongkol, P.; Bhoopat, T.; Kurahashi, H.; Chockjamsai, M.; Piangjai, S.; Bunchu, N.; et al. Forensic Entomology cases in Thailand: A review of cases from 2000 to 2006. Parasitol. Res. 2007, 101, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Syamsa, R.A.; Omar, B.; Ahmad, F.M.; Hidayatulfathi, O.; Shahrom, A.W. Comparative fly species composition on indoor and outdoor forensic cases in Malaysia. J. Forensic Leg. Med. 2017, 45, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Richards, C.S.; Price, B.W.; Villet, M.H. Thermal ecophysiology of seven carrion-feeding blowflies in Southern Africa. Entomol. Exp. Appl. 2009, 131, 11–19. [Google Scholar] [CrossRef]
- Rogers, D.J.; Williams, B.G. Monitoring trypanosomiasis in space and time. Parasitology 1993, 106 (Suppl. S1), S77–S92. [Google Scholar] [CrossRef]
- Bunchu, N.; Sukontason, K.L.; Olson, J.K.; Kurahashi, H.; Sukontason, K. Behavioral responses of Chrysomya megacephala to natural products. Parasitol. Res. 2008, 102, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Tumrasvin, W.; Kurahashi, H.; Kano, R. Studies on medically important flies in Thailand VII. Report on 42 species of Calliphorid flies, including the taxonomic keys (Diptera: Calliphoridae). Bull. Tokyo Med. Dent. Univ. 1979, 26, 243–272. [Google Scholar] [PubMed]
- Zabala, J.; Diaz, B.; Salona-Bordas, M.I. Seasonal blowfly distribution and abundance in fragmented landscapes. Is it useful in forensic inference about where a corpse has been decaying? PLoS ONE 2014, 9, e99668. [Google Scholar] [CrossRef] [PubMed]
- Monum, T.; Sukontason, K.L.; Sribanditmongkol, P.; Sukontason, K.; Samerjai, C.; Limsopatham, K.; Suwannayod, S.; Klong-klaew, T.; Wannasan, A. Forensically important blow flies Chrysomya pinguis, C. villeneuvi, and Lucilia porphyrina (Diptera: Calliphoridae) in a case of human remains in Thailand. Korean J. Parasitol. 2017, 55, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Kavazos, C.R.J.; Wallman, J.F. Community composition of carrion-breeding blowflies (Diptera: Calliphoridae) along an urban gradient in south-eastern Australia. Landsc. Urban Plan. 2012, 106, 183–190. [Google Scholar] [CrossRef]
- Baumgartner, D.L.; Greenberg, B. Distribution and medical ecology of the blow flies (Diptera: Calliphoridae) of Peru. Ann. Entomol. Soc. Am. 1985, 78, 565–587. [Google Scholar] [CrossRef]
- Grant, R.H.; Heisler, G.M.; Gao, W. Estimation of pedestrian level UV exposure under trees. Photochem. Photobiol. 2002, 75, 369–376. [Google Scholar] [CrossRef]
- Silahuddin, S.A.; Latif, B.; Kurahashi, H.; Heo, C.C. The importance of habitat in the ecology of decomposition on rabbit carcasses in Malaysia: Implications in Forensic Entomology. J. Med. Entomol. 2015, 52, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Weidner, L.M.; Jennings, D.E.; Tomberlin, J.K.; Hamilton, G.C. Seasonal and geographic variation in biodiversity of forensically important blow flies (Diptera: Calliphoridae) in New Jersey, USA. J. Med. Entomol. 2015, 52, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.; Turner, B.D. Spatial and temporal variability of necrophagous Diptera from urban to rural areas. Med. Vet. Entomol. 2005, 19, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, Y.; Wang, J.; Ma, M.; Lai, Y. Tempeature-dependent development and the significance for estimating postmortem interval of Chrysomya nigripes Aubertin, a new forensically important species in China. Int. J. Legal Med. 2016, 130, 1363–1370. [Google Scholar] [CrossRef] [PubMed]
- Owings, C.G.; Spiegelman, C.; Tarone, A.M.; Tomberlin, J.K. Developmental variation among Cochliomyia macellaria Fabricius (Diptera: Calliphoridae) populations from three ecoregions of Texas, USA. Int. J. Legal Med. 2014, 128, 709–717. [Google Scholar] [CrossRef] [PubMed]
- O’Flynn, M.A. Notes on the biology of Chrysomya nigripes Aubertin (Diptera: Calliphoridae). Aust. Entomol. 1983, 22, 341–342. [Google Scholar] [CrossRef]
- Ahmad, N.W.; Lim, L.H.; Dhang, C.C.; Chin, H.C.; Abdullah, A.G.; Mustaffa, W.-N.W.; Kian, C.W.; Jeffery, J.; Hashim, R.; Azirun, S.M. Comparative insect fauna succession on indoor and outdoor monkey carrions in a semi-forested area in Malaysia. Asian Pac. J. Trop. Biomed. 2011, 1 (Suppl. S2), S232–S238. [Google Scholar] [CrossRef]
- Yang, S.T.; Shiao, S.F. Temperature adaptation in Chrysomya megacephala and Chrysomya pinguis, two blow fly species of forensic significance. Entomol. Exp. Appl. 2014, 152, 100–107. [Google Scholar] [CrossRef]
- Azevedo, R.R.; Krüger, R.F. The influence of temperature and humidity on abundance and richness of Calliphoridae (Diptera). Iheringia Sér. Zool. 2013, 103, 145–152. [Google Scholar] [CrossRef]
- Mohr, R.M.; Tomberlin, J.K. Environmental factors affecting early carcass attendance by four species of blow flies (Diptera: Calliphoridae) in Texas. J. Med. Entomol. 2014, 51, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Charabidze, D.; Gosselin, M.; Hedouin, V. Use of necrophagous insects as evidence of cadaver relocation: Myth or reality? PeerJ 2017, 5, e3506. [Google Scholar] [CrossRef] [PubMed]
- Matuszewski, S.; Szafalowicz, M.; Jarmusz, M. Insects colonising carcasses in open and forest habitats of Central Europe: Search for indicators of corpse relocation. Forensic Sci. Int. 2013, 231, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Sukontason, K.L.; Vogtsberger, R.C.; Boonchu, N.; Chaiwong, T.; Sripakdee, D.; Ngern-Klun, R.; Piangjai, S.; Sukontason, K. Larval morphology of Chrysomya nigripes (Diptera: Calliphoridae), a fly species of forensic importance. J. Med. Entomol. 2005, 42, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Sukontason, K.L.; Bhoopat, T.; Wannasan, A.; Sontigun, N.; Sanit, S.; Amendt, J.; Samerjai, C.; Sukontason, K. Chrysomya chani Kurahashi (Diptera: Calliphoridae), a blow fly species of forensic importance: Morphological characters of the third larval instar and a case report from Thailand. Forensic Sci. Res. 2018, 3, 83–93. [Google Scholar] [CrossRef]
- Sukontason, K.; Sukontason, K.L.; Piangjai, S.; Narongchai, P.; Samai, W.; Boonchu, N.; Sripakdee, D.; Ngern-Klun, R.; Siriwattanarungsee, S. Morphology of second and third instars of Chrysomya villeneuvi Patton (Diptera: Calliphoridae), a fly species of forensic importance. Forensic Sci. Int. 2005, 154, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, R.; Nazni, W.A.; Tan, T.C.; Lee, H.L.; Isa, M.N.; Azirun, M.S. Molecular identification of blow flies recovered from human cadavers during crime scene investigations in Malaysia. Malays. J. Pathol. 2012, 34, 127–132. [Google Scholar] [PubMed]
- Bharti, M.; Singh, B. DNA-based identification of forensically important blow flies (Diptera: Calliphoridae) from India. J. Med. Entomol. 2017, 54, 1151–1156. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land Uses | Climatic Factors Recorded * | Number of Chrysomyinae Flies Collected | ||||||
|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | Relative Humidity (%) | Light Intensity (Lux) | C. pinguis | C. chani | C. villeneuvi | Cey. nigripes | Total | |
| mixed deciduous forest | 26.5 (16.7–39.8) | 67.0 (35–87) | 26,550 (193–107,500) | 417 | 282 | 163 | 119 | 981 |
| disturbed mixed deciduous forest | 28.4 (21.0–39.3) | 61.5 (35–80) | 59,403 (2320–359,600) | 0 | 2 | 2 | 5 | 9 |
| mixed orchard | 28.4 (19.7–45.2) | 59.0 (25–89) | 47,200 (1000–118,500) | 83 | 18 | 27 | 21 | 149 |
| paddy field | 27.5 (19.6–40.8) | 63.5 (39–83) | 36,300 (6700–112,000) | 11 | 44 | 44 | 42 | 141 |
| lowland village | 28.3 (18.7–35.8) | 65.0 (36–83) | 38,550 (2900–95,700) | 1 | 2 | 9 | 6 | 18 |
| city town | 25.8 (18.8–30.7) | 64.0 (40–78) | 22,000 (442–94,000) | 0 | 0 | 0 | 0 | 0 |
| Land Uses | n | Fly Species | |||
|---|---|---|---|---|---|
| C. pinguis | C. chani | C. villeneuvi | Cey. nigripes | ||
| mixed deciduous forest | 84 | 4.96 ± 1.88 a | 3.36 ± 0.91 a | 1.94 ± 0.04 a | 1.42 ± 0.71 a |
| disturbed mixed deciduous forest | 34 | 0 | 0.06 ± 0.04 b | 0.06 ± 0.04 b | 0.15 ± 0.10 a |
| mixed orchard | 101 | 0.82 ± 0.35 b | 0.18 ± 0.07 b | 0.27 ± 0.12 b | 0.21 ± 0.12 a |
| paddy field | 34 | 0.32 ± 0.19 b | 1.29 ± 1.20 b | 1.29 ± 0.90 a,b | 1.24 ± 1.14 a |
| lowland village | 34 | 0.03 ± 0.03 b | 0.06 ± 0.06 b | 0.26 ± 0.16 b | 0.18 ± 0.08 a |
| city town | 15 | 0 | 0 | 0 | 0 |
| Climatic Factors | Fly Species | ||||
|---|---|---|---|---|---|
| C. pinguis | C. chani | C. villeneuvi | Cey. nigripes | ||
| temperature | r | −0.232 ** | −0.003 | −0.83 | 0.017 |
| p | 0.000 | 0.961 | 0.148 | 0.768 | |
| relative humidity | r | 0.051 | 0.064 | 0.086 | −0.042 |
| p | 0.374 | 0.266 | 0.138 | 0.462 | |
| light intensity | r | −0.319 ** | −0.102 | −0.180 ** | 0.016 |
| p | 0.000 | 0.078 | 0.002 | 0.779 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klong-klaew, T.; Ngoen-klan, R.; Moophayak, K.; Sukontason, K.; Irvine, K.N.; Tomberlin, J.K.; Somboon, P.; Chareonviriyaphap, T.; Kurahashi, H.; Sukontason, K.L. Predicting Geographic Distribution of Forensically Significant Blow Flies of Subfamily Chrysomyinae (Diptera: Calliphoridae) in Northern Thailand. Insects 2018, 9, 106. https://doi.org/10.3390/insects9030106
Klong-klaew T, Ngoen-klan R, Moophayak K, Sukontason K, Irvine KN, Tomberlin JK, Somboon P, Chareonviriyaphap T, Kurahashi H, Sukontason KL. Predicting Geographic Distribution of Forensically Significant Blow Flies of Subfamily Chrysomyinae (Diptera: Calliphoridae) in Northern Thailand. Insects. 2018; 9(3):106. https://doi.org/10.3390/insects9030106
Chicago/Turabian StyleKlong-klaew, Tunwadee, Ratchadawan Ngoen-klan, Kittikhun Moophayak, Kom Sukontason, Kim N. Irvine, Jeffery K. Tomberlin, Pradya Somboon, Theeraphap Chareonviriyaphap, Hiromu Kurahashi, and Kabkaew L. Sukontason. 2018. "Predicting Geographic Distribution of Forensically Significant Blow Flies of Subfamily Chrysomyinae (Diptera: Calliphoridae) in Northern Thailand" Insects 9, no. 3: 106. https://doi.org/10.3390/insects9030106
APA StyleKlong-klaew, T., Ngoen-klan, R., Moophayak, K., Sukontason, K., Irvine, K. N., Tomberlin, J. K., Somboon, P., Chareonviriyaphap, T., Kurahashi, H., & Sukontason, K. L. (2018). Predicting Geographic Distribution of Forensically Significant Blow Flies of Subfamily Chrysomyinae (Diptera: Calliphoridae) in Northern Thailand. Insects, 9(3), 106. https://doi.org/10.3390/insects9030106