Preliminary Investigation of Species Diversity of Rice Hopper Parasitoids in Southeast Asia

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study System and Sampling
2.2. Preliminary Morphological Identification
2.3. DNA Extraction and Amplification
2.4. Data Analysis
2.5. Sequence Data Deposition
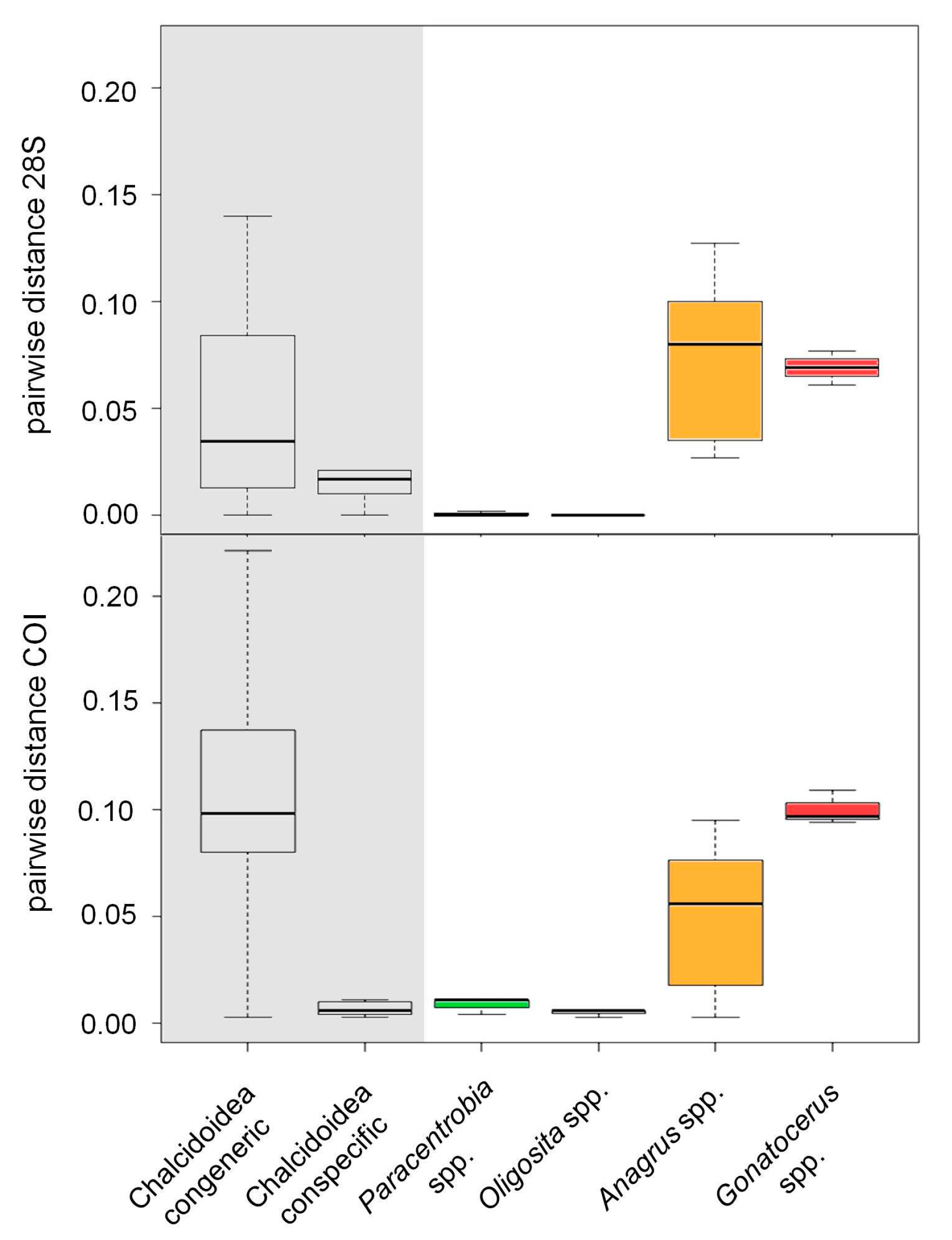
3. Results
3.1. Morphological Analysis
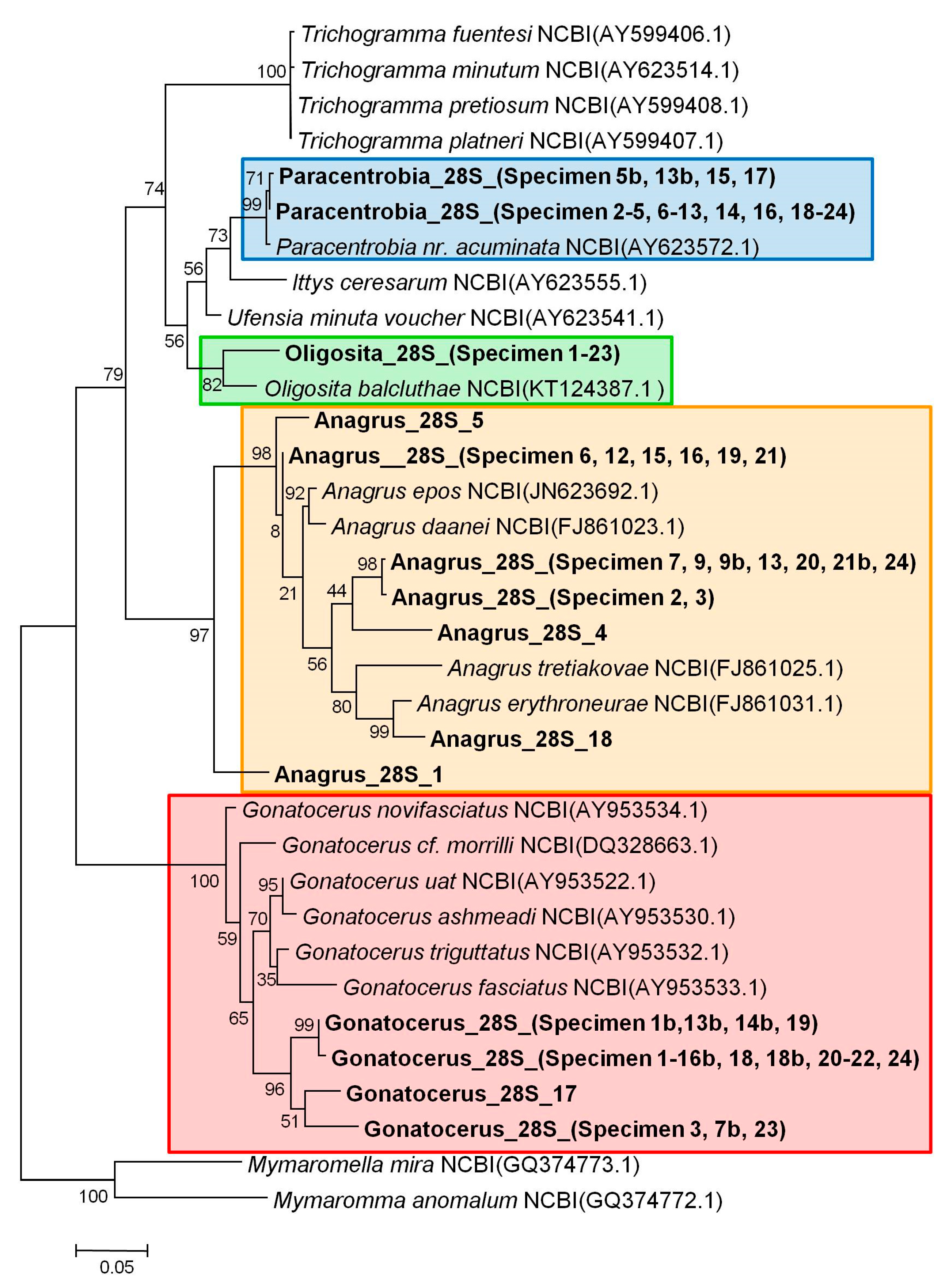
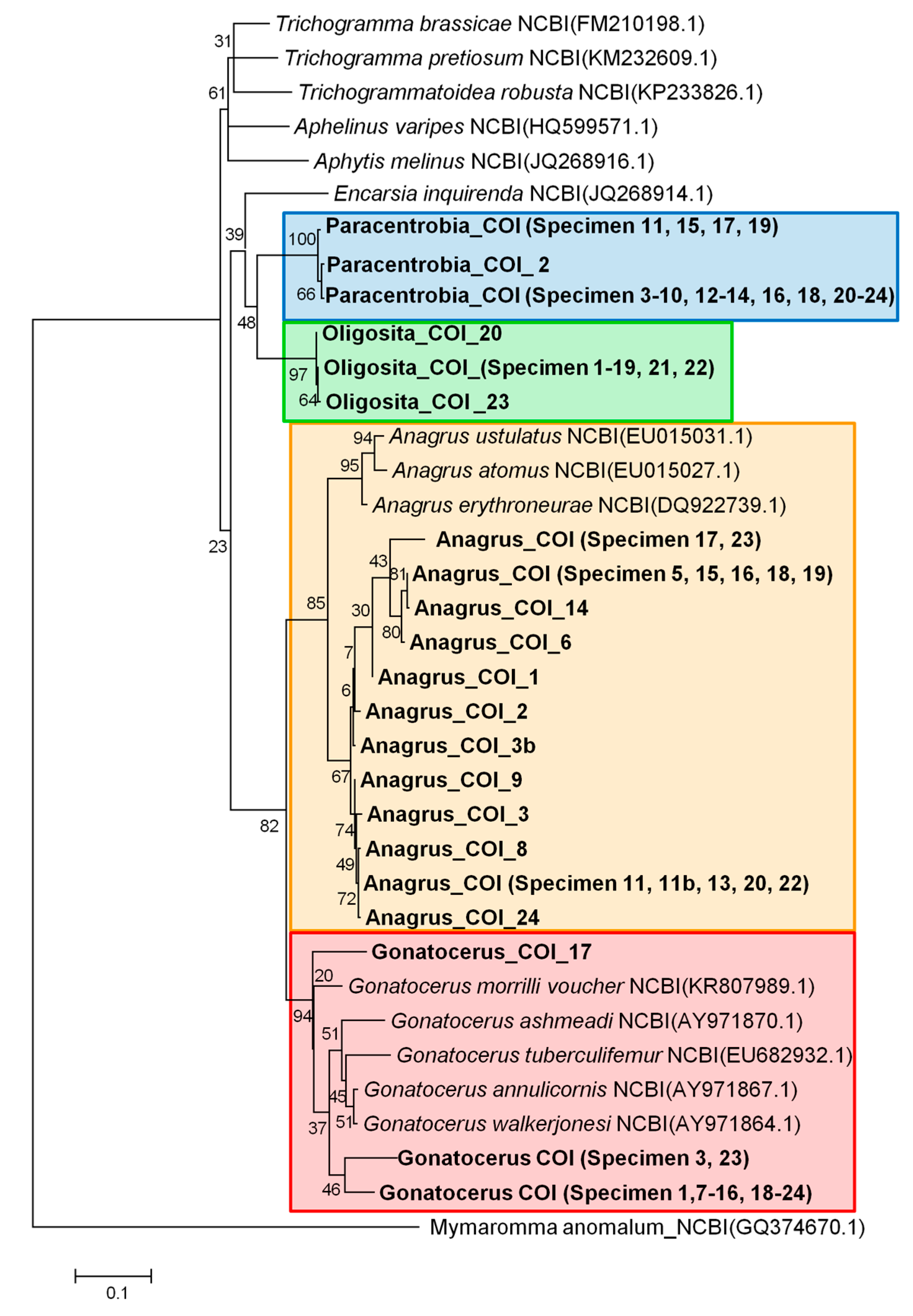
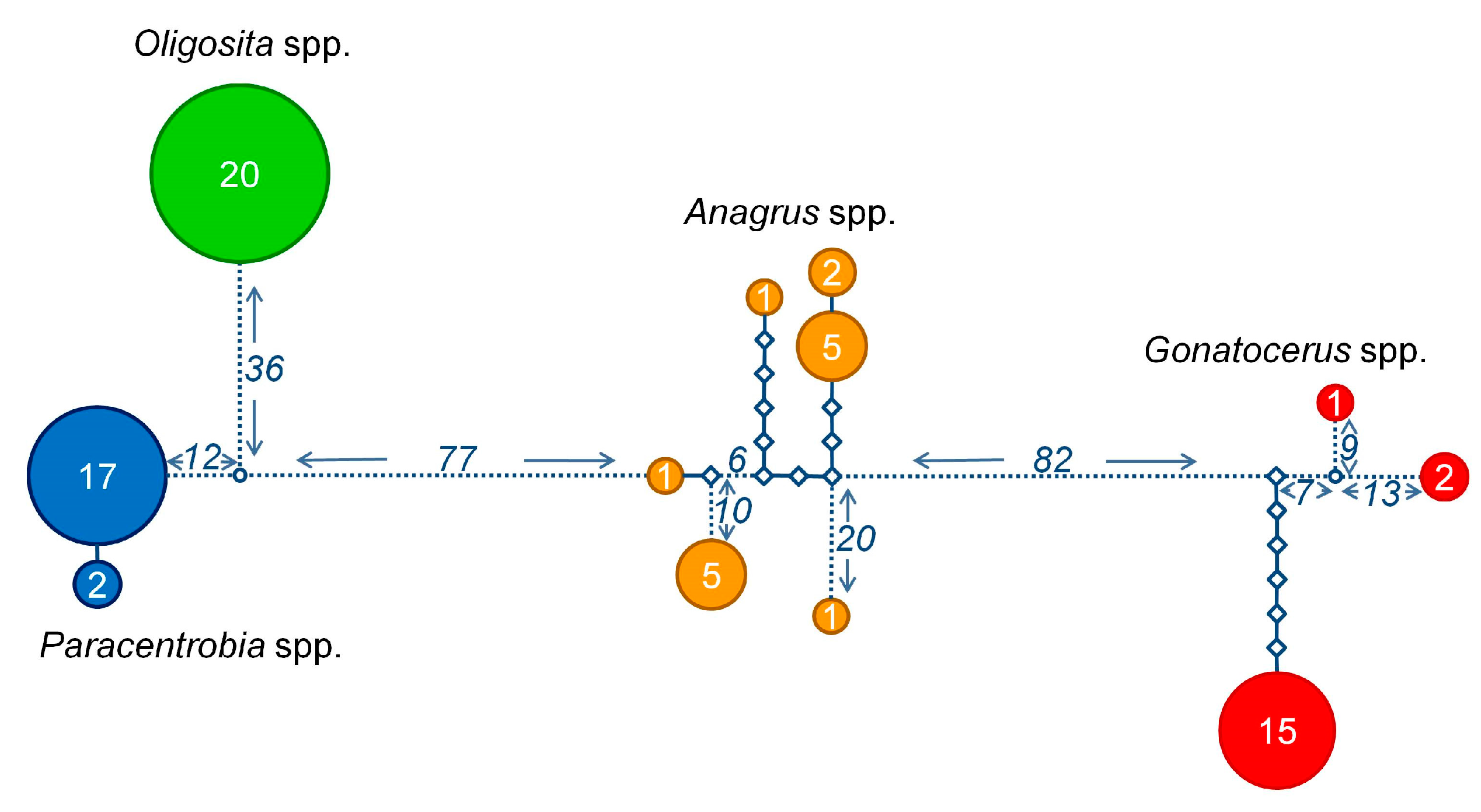
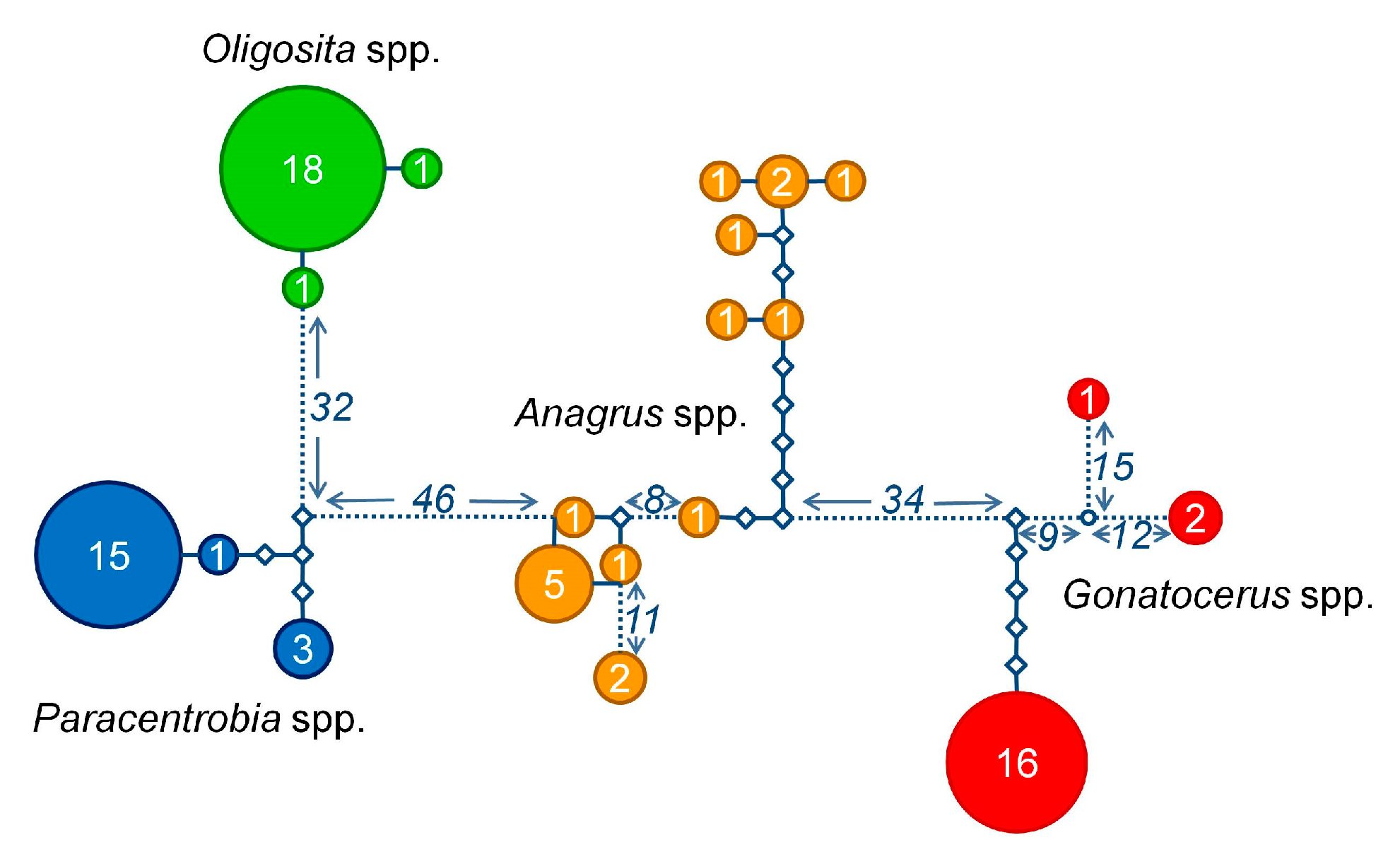
3.2. Molecular Analysis
3.3. Comparison between the Morphological and Molecular Approach
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAO. Available online: http://www.fao.org/faostat/en/#home (accessed on 30 March 2017).
- IRRI—Trends in Global Rice Consumption. Available online: http://irri.org/rice-today/trends-in-global-rice-consumption (accessed on 1 June 2017).
- Gurr, G.M.; Lu, Z.; Zheng, X.; Xu, H.; Zhu, P.; Chen, G.; Yao, X.; Cheng, J.; Zhu, Z.; Catindig, J.L.; et al. Multi-country evidence that crop diversification promotes ecological intensification of agriculture. Nat. Plants 2016, 2, 16014. [Google Scholar] [CrossRef] [PubMed]
- Heong, K.L.; Aquino, G.-B.; Barrion, A.T. Population dynamics of plant- and leafhoppers and their natural enemies in rice ecosystems in the Philippines. Crop Prot. 1992, 11, 371–379. [Google Scholar] [CrossRef]
- Ling, K.C. Rice Virus Diseases; International Rice Research Institute: Los Banos, Philippines, 1972. [Google Scholar]
- Cabauatan, P.Q.; Cabunagan, R.C.; Choi, I.-R. Rice viruses transmitted by the brown planthopper Nilaparvata lugens Stal. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Banos, Philippines, 2009; pp. 357–368. [Google Scholar]
- Catindig, J.L.; Arida, G.S.; Baehaki, S.E.; Bentur, J.S.; Cuong, L.Q.; Norowi, M.; Rattanakarn, W.; Sriratanasak, W.; Xia, J.; Lu, Z. Situations of planthoppers in Asia. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Banos, Philippines, 2009; pp. 191–220. [Google Scholar]
- Seo, B.Y.; Jung, J.K.; Choi, B.-R.; Park, H.M.; Lee, B.H. Resistance-breaking ability and feeding behavior of the brown planthopper Nilaparvata lugens, recently collected in Korea. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; International Rice Research Institute: Los Banos, Philippines, 2009; pp. 303–314. [Google Scholar]
- Heong, K.L.; Schoenly, K.G. Impact of insecticides on herbivore-natural enemy communities in tropical rice ecosystems. In Ecotoxicology; Springer: Boston, MA, USA, 1998; pp. 381–403. ISBN 978-1-4613-7653-8. [Google Scholar]
- Heong, K.L. Are planthopper problems caused by a breakdown in ecosystem services? In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Banos, Philippines, 2009; pp. 221–232. ISBN 978-971-22-0251-3. [Google Scholar]
- Gurr, G.M.; Liu, J.; Read, D.M.Y.; Catindig, J.L.A.; Cheng, J.A.; Lan, L.P.; Heong, K.L. Parasitoids of Asian rice planthopper (Hemiptera: Delphacidae) pests and prospects for enhancing biological control by ecological engineering. Ann. Appl. Biol. 2011, 158, 149–176. [Google Scholar] [CrossRef]
- Antolin, M.F.; Strong, D.R. Long-distance dispersal by a parasitoid (Anagrus delicatus, Mymaridae) and its host. Oecologia 1987, 73, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Fowler, S.V.; Claridge, M.F.; Morgan, J.C.; Peries, I.D.R.; Nugaliyadde, L. Egg mortality of the brown planthopper, Nilaparvata lugens (Homoptera: Delphacidae) and green leafhoppers, Nephotettix spp. (Homoptera: Cicadellidae), on rice in Sri Lanka. Bull. Entomol. Res. 1991, 81, 161–167. [Google Scholar] [CrossRef]
- Watanabe, T.; Wada, T.; Salleh, N.M.N. Parasitic Activities of Egg Parasitoids on the rice planthoppers, Nilaparvata lugens (STAL) and Sogatella furcifera (HORVATH) (Homoptera: Delphacidae), in the Muda Area, Peninsular Malaysia. Appl. Entomol. Zool. 1992, 27, 205–211. [Google Scholar] [CrossRef]
- Drechsler, M.; Settele, J. Predator–prey interactions in rice ecosystems: Effects of guild composition, trophic relationships, and land use changes—A model study exemplified for Philippine rice terraces. Ecol. Model. 2001, 137, 135–159. [Google Scholar] [CrossRef]
- Nishida, T.; Wongsiri, T.; Wongsiri, N. Species composition, population trends and egg parasitism of planthopper and leafhopper rice pests of Thailand. Plant Prot. Bull. FAO 1976, 24, 22–26. [Google Scholar]
- Chesters, D.; Wang, Y.; Yu, F.; Bai, M.; Zhang, T.-X.; Hu, H.-Y.; Zhu, C.-D.; Li, C.-D.; Zhang, Y.-Z. The integrative taxonomic approach reveals host specific species in an encyrtid parasitoid species Complex. PLoS ONE 2012, 7, e37655. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the Neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Rodriguez, J.J.; Whitfield, J.B.; Deans, A.R.; Janzen, D.H.; Hallwachs, W.; Hebert, P.D.N. Extreme diversity of tropical parasitoid wasps exposed by iterative integration of natural history, DNA barcoding, morphology, and collections. Proc. Natl. Acad. Sci. USA 2008, 105, 12359–12364. [Google Scholar] [CrossRef] [PubMed]
- Schwarzfeld, M.D.; Broad, G.R.; Sperling, F.A.H. Molecular phylogeny of the diverse parasitoid wasp genus Ophion Fabricius (Hymenoptera: Ichneumonidae: Ophioninae): Phylogeny of Ophion. Syst. Entomol. 2016, 41, 191–206. [Google Scholar] [CrossRef]
- Munro, J.B.; Heraty, J.M.; Burks, R.A.; Hawks, D.; Mottern, J.; Cruaud, A.; Rasplus, J.-Y.; Jansta, P. A molecular phylogeny of the Chalcidoidea (Hymenoptera). PLoS ONE 2011, 6, e27023. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Woodley, N.E.; Janzen, D.H.; Hallwachs, W.; Hebert, P.D.N. DNA barcodes reveal cryptic host-specificity within the presumed polyphagous members of a genus of parasitoid flies (Diptera: Tachinidae). Proc. Natl. Acad. Sci. USA 2006, 103, 3657–3662. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.-S.; Xi, Y.-Q.; Yu, F.; Zhang, X.; Li, X.-J.; Liu, C.-L.; Niu, Z.-Q.; Zhu, C.-D.; Qiao, G.-X.; Zhang, Y.-Z. Application of DNA barcoding to the identification of Hymenoptera parasitoids from the soybean aphid (Aphis glycines) in China: DNA barcoding to identify parasitoids from Aphis glycines in China. Insect Sci. 2014, 21, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Wood, D.M.; Janzen, D.H.; Hallwachs, W.; Hebert, P.D.N. DNA barcodes affirm that 16 species of apparently generalist tropical parasitoid flies (Diptera, Tachinidae) are not all generalists. Proc. Natl. Acad. Sci. USA 2007, 104, 4967–4972. [Google Scholar] [CrossRef] [PubMed]
- Padial, J.M.; Miralles, A.; De la Riva, I.; Vences, M. The integrative future of taxonomy. Front. Zool. 2010, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Sann, C.; Theodorou, P.; Heong, K.L.; Villareal, S.; Settele, J.; Vidal, S.; Westphal, C. Hopper parasitoids do not significantly benefit from non-crop habitats in rice production landscapes. Agric. Ecosyst. Environ. 2018, 254, 224–232. [Google Scholar] [CrossRef]
- Settele, J.; Spangenberg, J.H.; Heong, K.L.; Burkhard, B.; Bustamante, J.V.; Cabbigat, J.; Van Chien, H.; Escalada, M.; Grescho, V.; Hai, L.H.; et al. Agricultural landscapes and ecosystem services in South-East Asia—The LEGATO-Project. Basic Appl. Ecol. 2015, 16, 661–664. [Google Scholar] [CrossRef]
- Heinrichs, E.A.; Medrano, F.G.; Rapusas, H.R. Genetic Evaluation for Insect in Rice; International Rice Research Institute: Los Banos, Philippines, 1985; ISBN 971-10-4110-3. [Google Scholar]
- Shepard, B.M.; Barrion, A.T.; Listinger, J.A. Friends of the Rice Farmer. Helpful Insects, Spiders and Pathogens; International Rice Research Institute: Los Banos, Philippines, 1987; ISBN 971-10-4162-6. [Google Scholar]
- Dupo, A.L.; Barrion, A.T. Taxonomy and general biology of delphacid planthoppers in rice agroecosystems. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Banos, Philippines, 2009; pp. 3–156. ISBN 978-971-22-0251-3. [Google Scholar]
- Heinrichs, E.A. Biology and Management of Rice Insects; Wiley Eastern Limited: New Delhi, India, 1994; ISBN 81-224-0581-2. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Campbell, B.C.; Steffen-Campbell, J.D.; Werren, J.H. Phylogeny of the Nasonia species complex (Hymenoptera: Pteromalidae) inferred from an internal transcribed spacer (ITS2) and 28S rDNA sequences. Insect Mol. Biol. 1993, 2, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.C.; Heraty, J.M.; Rasplus, J.Y.; Chan, K.; Steffen-Campbell, J.D.; Babcock, C. Molecular systematic of the Chalcidoidea using 28S-D2 rDNA. In Hymenoptera: Evolution, Biodiversity and Biological Control; Heong, K.L., Hardy, B., Eds.; CSIRO Publishing: Collingwood, Australia, 2000; pp. 59–73. ISBN 0-643-06610-1. [Google Scholar]
- Bonfield, J.K.; Whitwham, A. Gap5—Editing the billion fragment sequence assembly. Bioinformatics 2010, 26, 1699–1703. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Dunn, O.J. Multiple comparisons using rank sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- R Development Core Team. The R Foundation for Statistical Computing, Ver. 3.0.2; Vienna University of Technology: Vienna, Austria, 2013. Available online: http://www.r-project.org/ (accessed on 3 May 2017).
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.; Fernández-Triana, J.L.; Eveleigh, E.; Gómez, J.; Guclu, C.; Hallwachs, W.; Hebert, P.D.N.; Hrcek, J.; Huber, J.T.; Janzen, D.; et al. DNA barcoding and the taxonomy of Microgastrinae wasps (Hymenoptera, Braconidae): Impacts after 8 years and nearly 20000 sequences. Mol. Ecol. Resour. 2013, 13, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, S.G.; Buerki, S.; Hansson, C.; Alvarez, N.; Benrey, B. Uncovering cryptic parasitoid diversity in Horismenus (Chalcidoidea, Eulophidae). PLoS ONE 2015, 10, e0136063. [Google Scholar] [CrossRef] [PubMed]
- Mottern, J.L.; Heraty, J.M. Revision of the Cales noacki species complex (Hymenoptera, Chalcidoidea, Aphelinidae). Syst. Entomol. 2014, 39, 354–379. [Google Scholar] [CrossRef]
- Brower, A.V.Z. Problems with DNA barcodes for species delimitation: “Ten species” of Astraptes fulgerator reassessed (Lepidoptera: Hesperiidae). Syst. Biodivers. 2006, 4, 127–132. [Google Scholar] [CrossRef]
- Collins, R.A.; Cruickshank, R.H. The seven deadly sins of DNA barcoding. Mol. Ecol. Resour. 2013, 13, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Arellano, D.; Gutiérrez-Arellano, C.; Zaldívar-Riverón, A. DNA Barcoding of the parasitoid wasp subfamily Doryctinae (Hymenoptera: Braconidae) from Chamela, Mexico. Biodivers. Data J. 2015, 3, e5109. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Babcock, C.S.; Heraty, J.M.; De Barro, P.J.; Driver, F.; Schmidt, S. Preliminary phylogeny of Encarsia Förster (Hymenoptera: Aphelinidae) based on morphology and 28S rDNA. Mol. Phylogenet. Evol. 2001, 18, 306–323. [Google Scholar] [CrossRef] [PubMed]
- Manzari, S.; Polaszek, A.; Belshaw, R.; Quicke, D.L.J. Morphometric and molecular analysis of the Encarsia inaron species-group (Hymenoptera: Aphelinidae), parasitoids of whiteflies (Hemiptera: Aleyrodidae). Bull. Entomol. Res. 2002, 92, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Triapitsyn, S.V.; Vickerman, D.B.; Heraty, J.M.; Logarzo, G.A. A new species of Gonatocerus (Hymenoptera: Mymaridae) parasitic On Proconiine Sharpshooters (Hemiptera: Cicadellidae) in the New World. Zootaxa 2006, 1158, 55–67. [Google Scholar]
- Gillespie, J.J. A secondary structural model of the 28S rRNA expansion segments D2 and D3 for chalcidoid wasps (Hymenoptera: Chalcidoidea). Mol. Biol. Evol. 2005, 22, 1593–1608. [Google Scholar] [CrossRef] [PubMed]
- Greenstone, M.H. Molecular methods for assessing insect parasitism. Bull. Entomol. Res. 2006, 96, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gariepy, T.D.; Kuhlmann, U.; Gillott, C.; Erlandson, M. Parasitoids, predators and PCR: The use of diagnostic molecular markers in biological control of Arthropods. J. Appl. Entomol. 2007, 131, 225–240. [Google Scholar] [CrossRef]
- Desneux, N.; Starý, P.; Delebecque, C.J.; Gariepy, T.D.; Barta, R.J.; Hoelmer, K.A.; Heimpel, G.E. Cryptic species of parasitoids attacking the soybean aphid (Hemiptera: Aphididae) in Asia: Binodoxys communis and binodoxys koreanus (Hymenoptera: Braconidae: Aphidiinae). Ann. Entomol. Soc. Am. 2009, 102, 925–936. [Google Scholar] [CrossRef]
- Zheng, X.; Yu, X.; Lu, Z.; Chen, J.; Xu, H.; Ju, R. Parasitization adaptability of Anagrus optabilis on Nilaparvata lugens. Chin. J. Biol. Control 2003, 19, 136–138. [Google Scholar]
- Zheng, X.; Lu, Y.; Zhu, P.; Zhang, F.; Tian, J.; Xu, H.; Chen, G.; Nansen, C.; Lu, Z. Use of banker plant system for sustainable management of the most important insect pest in rice fields in China. Sci. Rep. 2017, 7, 45581. [Google Scholar] [CrossRef] [PubMed]
- Meier, R.; Zhang, G.; Ali, F. The use of mean instead of smallest interspecific distances exaggerates the size of the “Barcoding Gap” and leads to misidentification. Syst. Biol. 2008, 57, 809–813. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Host | Examined Individuals |
|---|---|---|
| Paracentrobia andoi | GLH | 50 |
| Oligosita aesopi | BPH | 41 |
| Oligosita naias | BPH | 7 |
| Oligosita shibuyae | BPH | 2 |
| Anagrus frequens | BPH | 33 |
| Anagrus optabilis | BPH | 6 |
| Anagrus flaveolus | BPH | 11 |
| Gonatocerus orientalis | GLH | 50 |
| Gene | Parasitoid Group | Paracentrobia spp. | Oligosita spp. | Anagrus spp. | Gonatocerus spp. | ||||
| p | df | p | df | p | df | p | df | ||
| 28S gene | Chalcidoidea congeneric | 0.003 | 2 | 0.003 | 2 | 0.021 | 2 | n.s. | 2 |
| Chalcidoidea conspecific | n.s. | 2 | n.s. | 2 | <0.001 | 2 | 0.05 | 2 | |
| COI gene | Chalcidoidea congeneric | 0.017 | 2 | 0.002 | 2 | <0.001 | 2 | n.s. | 2 |
| Chalcidoidea conspecific | n.s. | 2 | n.s. | 2 | <0.001 | 2 | 0.006 | 2 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sann, C.; Wemheuer, F.; Beaurepaire, A.; Daniel, R.; Erler, S.; Vidal, S. Preliminary Investigation of Species Diversity of Rice Hopper Parasitoids in Southeast Asia. Insects 2018, 9, 19. https://doi.org/10.3390/insects9010019
Sann C, Wemheuer F, Beaurepaire A, Daniel R, Erler S, Vidal S. Preliminary Investigation of Species Diversity of Rice Hopper Parasitoids in Southeast Asia. Insects. 2018; 9(1):19. https://doi.org/10.3390/insects9010019
Chicago/Turabian StyleSann, Christina, Franziska Wemheuer, Alexis Beaurepaire, Rolf Daniel, Silvio Erler, and Stefan Vidal. 2018. "Preliminary Investigation of Species Diversity of Rice Hopper Parasitoids in Southeast Asia" Insects 9, no. 1: 19. https://doi.org/10.3390/insects9010019
APA StyleSann, C., Wemheuer, F., Beaurepaire, A., Daniel, R., Erler, S., & Vidal, S. (2018). Preliminary Investigation of Species Diversity of Rice Hopper Parasitoids in Southeast Asia. Insects, 9(1), 19. https://doi.org/10.3390/insects9010019