
Responses of Crop Pests and Natural Enemies to Wildflower Borders Depends on Functional Group


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
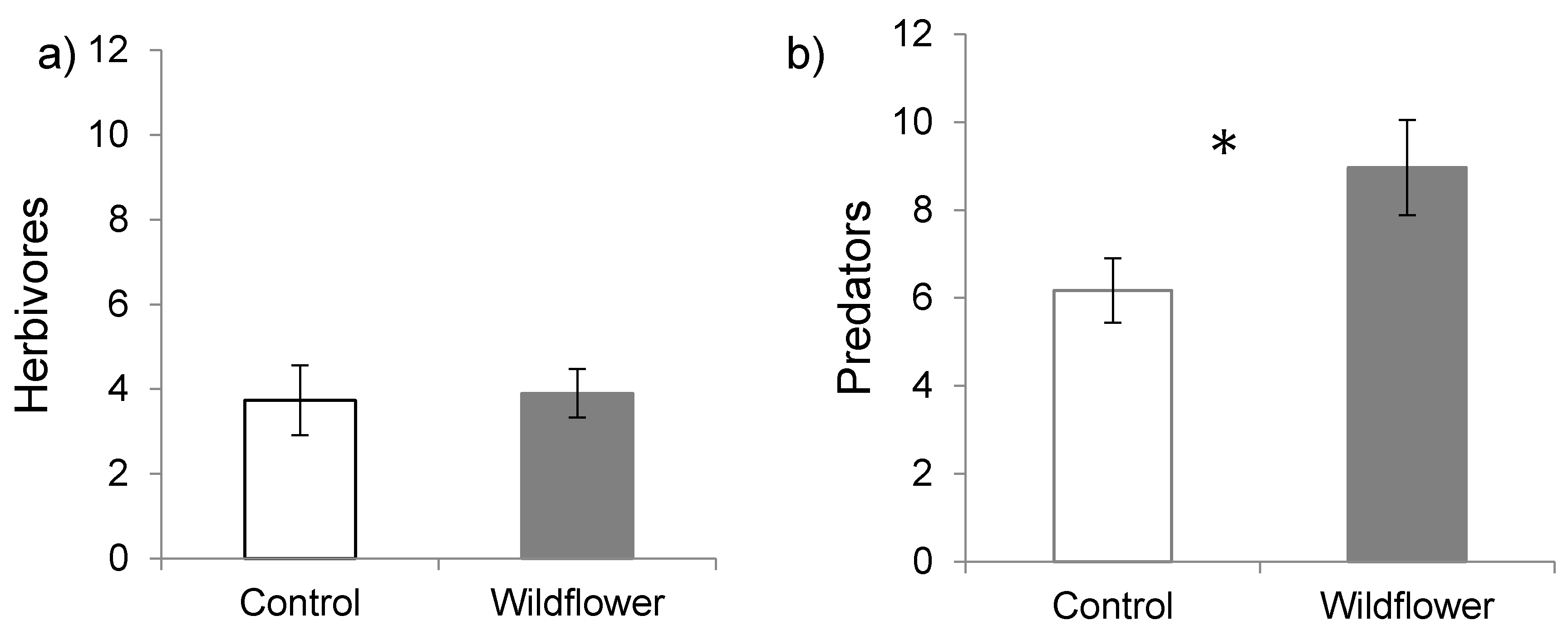
3. Results
4. Discussion
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Altieri, M. The ecological role of biodiversity in agroecosystem. Agric. Ecosyst. Environ. 1999, 74, 19–31. [Google Scholar] [CrossRef]
- Pywell, R.F.; Heard, M.S.; Woodcock, B.A.; Hinsley, S.; Ridding, L.; Nowakowski, M.; Bullock, J.M. Wildlife-friendly farming increases crop yield: Evidence for ecological intensification. Proc. R. Soc. B Biol. Sci. 2015, 282. [Google Scholar] [CrossRef] [PubMed]
- Holzschuh, A.; Dudenhöffer, J.H.; Tscharntke, T. Landscapes with wild bee habitats enhance pollination, fruit set and yield of sweet cherry. Biol. Conserv. 2012, 153, 101–107. [Google Scholar] [CrossRef]
- Thies, C.; Tscharntke, T. Landscape structure and biological control in agroecosystems. Science 1999, 285, 893–895. [Google Scholar] [CrossRef] [PubMed]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity—Ecosystem service management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; Day, J.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Tscharntke, T.; Clough, Y.; Wanger, T.C.; Jackson, L.; Motzke, I.; Perfecto, I.; Vandermeer, J.; Whitbread, A. Global food security, biodiversity conservation and the future of agricultural intensification. Biol. Conserv. 2012, 151, 53–59. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef] [PubMed]
- Rusch, A.; Chaplin-Kramer, R.; Gardiner, M.M.; Hawro, V.; Holland, J.; Landis, D.; Thies, C.; Tscharntke, T.; Weisser, W.W.; Winqvist, C.; et al. Agricultural landscape simplification reduces natural pest control: A quantitative synthesis. Agric. Ecosyst. Environ. 2016, 221, 198–204. [Google Scholar] [CrossRef]
- Chaplin-Kramer, R.; O’Rourke, M.E.; Blitzer, E.J.; Kremen, C. A meta-analysis of crop pest and natural enemy response to landscape complexity. Ecol. Lett. 2011, 14, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, A.K.; Landis, D.A.; Wratten, S.D. Maximizing ecosystem services from conservation biological control: The role of habitat management. Biol. Control 2008, 45, 254–271. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef] [PubMed]
- van Emden, H.F.; Emden, V.; Helmut, F. Conservation Biological Control: From Theory to Practice. In Proceedings of the International Symposium on Biological Control of Arthropods, Honolulu, HI, USA, 14–18 January 2002. [Google Scholar]
- Walton, N.J.; Isaacs, R. Influence of native flowering plant strips on natural enemies and herbivores in adjacent blueberry fields. Environ. Entomol. 2011, 40, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Blaauw, B.R.; Isaacs, R. Wildflower plantings enhance the abundance of natural enemies and their services in adjacent blueberry fields. Biol. Control 2015, 91, 94–103. [Google Scholar] [CrossRef]
- Pfiffner, L.; Luka, H.; Schlatter, C.; Juen, A.; Traugott, M. Impact of wildflower strips on biological control of cabbage lepidopterans. Agric. Ecosyst. Environ. 2009, 129, 310–314. [Google Scholar] [CrossRef]
- Balmer, O.; Géneau, C.E.; Belz, E.; Weishaupt, B.; Förderer, G.; Moos, S.; Ditner, N.; Juric, I.; Luka, H. Wildflower companion plants increase pest parasitation and yield in cabbage fields: Experimental demonstration and call for caution. Biol. Control 2014, 76, 19–27. [Google Scholar] [CrossRef]
- Tschumi, M.; Albrecht, M.; Collatz, J.; Dubsky, V.; Entling, M.H.; Najar-Rodriguez, A.J.; Jacot, K. Tailored flower strips promote natural enemy biodiversity and pest control in potato crops. J. Appl. Ecol. 2016, 53, 1169–1176. [Google Scholar] [CrossRef]
- Tschumi, M.; Albrecht, M.; Entling, M.H.; Jacot, K. High effectiveness of tailored flower strips in reducing pests and crop plant damage. Proc. Biol. Sci. 2015. [Google Scholar] [CrossRef] [PubMed]
- Balzan, M.V.; Bocci, G.; Moonen, A.C. Augmenting flower trait diversity in wildflower strips to optimise the conservation of arthropod functional groups for multiple agroecosystem services. J. Insect Conserv. 2014, 18, 713–728. [Google Scholar] [CrossRef]
- Olson, D.M.; Wäckers, F.L. Management of field margins to maximize multiple ecological services. J. Appl. Ecol. 2007, 44, 13–21. [Google Scholar] [CrossRef]
- Berndt, L.A.; Wratten, S.D.; Scarratt, S.L. The influence of floral resource subsidies on parasitism rates of leafrollers (Lepidoptera: Tortricidae) in New Zealand vineyards. Biol. Control 2006, 37, 50–55. [Google Scholar] [CrossRef]
- Phillips, B.W.; Gardiner, M.M. Does local habitat management or large-scale landscape composition alter the biocontrol services provided to pumpkin agroecosystems? Biol. Control 2016, 92, 181–194. [Google Scholar] [CrossRef]
- Tscharntke, T.; Karp, D.S.; Chaplin-Kramer, R.; Batáry, P.; DeClerck, F.; Gratton, C.; Hunt, L.; Ives, A.; Jonsson, M.; Larsen, A.; et al. When natural habitat fails to enhance biological pest control—Five hypotheses. Biol. Conserv. 2016, 204, 449–458. [Google Scholar] [CrossRef]
- Jonsson, M.; Straub, C.S.; Didham, R.K.; Buckley, H.L.; Case, B.S.; Hale, R.J.; Gratton, C.; Wratten, S.D. Experimental evidence that the effectiveness of conservation biological control depends on landscape complexity. J. Appl. Ecol. 2015, 52, 1274–1282. [Google Scholar] [CrossRef]
- Wilson, H.; Miles, A.F.; Daane, K.M.; Altieri, M.A. Landscape diversity and crop vigor outweight influence of local diversification on biological control of a vineyard pest. Ecosphere 2017, 8. [Google Scholar] [CrossRef]
- Bommarco, R.; Miranda, F.; Bylund, H.; Björkman, C. Insecticides suppress natural enemies and increase pest damage in cabbage. J. Econ. Entomol. 2011, 104, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Forehand, A.L.M.; Orr, D.B.; Linker, H.M. Insect communities associated with beneficial insect habitat plants in North Carolina. Environ. Entomol. 2006, 35, 1541–1549. [Google Scholar] [CrossRef]
- Feeny, P. Plant apparency and chemical defense. In Biochemical Interactions Between Plants and Insects; Wallace, J., Mansell, R., Eds.; Springer: London, UK, 1976; pp. 1–40. [Google Scholar]
- Castagneyrol, B.; Giffard, B.; Christelle, P. Plant apparency, an overlooked driver of associational resistance to insect herbivory. J. Ecol. 2013, 418–429. [Google Scholar] [CrossRef]
- Young, O. Host plants of the tarnished plant bug, Lygus lineolaris (Heteroptera: Miridae). Ann. Entomol. Soc. Am. 1986, 79, 747–762. [Google Scholar] [CrossRef]
- Schaefers, G.A. Yield effects of tarnished plant bug feeding on june-bearing strawberry varieties in New York State. J. Econ. Entomol. 1980, 73, 721–725. [Google Scholar] [CrossRef]
- Bostainian, N.J. The Tarnished Plant Bug and Strawberry Production; Agriculture and Agri-Food Canada: Ottawa, Cannada, 1994. [Google Scholar]
- Balzan, M.V.; Bocci, G.; Moonen, A.C. Utilisation of plant functional diversity in wildflower strips for the delivery of multiple agroecosystem services. Entomol. Exp. Appl. 2016, 158, 304–319. [Google Scholar] [CrossRef]
- Connell, W.A. Stelidota geminata (Say) infestations of strawberries (Coleoptera: Nitidulidae). Entomol. News 1980, 91, 55–56. [Google Scholar]
- Weber, R.G.; Connell, W.A. Stelidota geminata (Say): Studies of its biology (Coleoptera: Nitidulidae). Ann. Entomol. Soc. Am. 1975, 68, 649–653. [Google Scholar] [CrossRef]
- Loughner, R.L.; Loeb, G.M.; Demchak, K.; Schloemann, S. Evaluation of strawberry sap beetle (Coleoptera: Nitidulidae) use of habitats surrounding strawberry plantings as food resources and overwintering sites. Environ. Entomol. 2007, 36, 1059–1065. [Google Scholar] [CrossRef]
- Day, W.H. Evaluation of biological control of the tarnished plant bug (Hemiptera: Miridae) in alfalfa by the introduced parasite Peristenus digoneutis (Hymenoptera: Braconidae). Environ. Entomol. 1996, 25, 512–518. [Google Scholar] [CrossRef]
- Weiss, M.J.; Williams, R.N. Some host-parasite relationships of Microctonus nitidulidid and Stelidota geminata. Ann. Entomol. Soc. Am. 1980, 73, 323–326. [Google Scholar] [CrossRef]
- Tuell, J.K.; Fiedler, A.K.; Landis, D.; Isaacs, R.; Tuell, J.K.; Fiedler, A.K.; Landis, D. Visitation by Wild and Managed Bees (Hymenoptera : Apoidea) to Eastern U.S. Native Plants for Use in Conservation Programs. Environ. Entomol. 2008, 37, 707–718. [Google Scholar] [CrossRef]
- Isaacs, R.; Tuell, J.; Fiedler, A.; Gardiner, M.; Landis, D. Maximizing arthropod-mediated ecosystem services in agricultural landscapes: The role of native plants. Front. Ecol. Environ. 2009, 7, 196–203. [Google Scholar] [CrossRef]
- Baars, M.A. Catches in pitfall traps in relation to mean densities of carabid beetles. Oecologia 1979, 41, 25–46. [Google Scholar] [CrossRef] [PubMed]
- Spence, J.R.; Niemelä, J.K. Sampling Carabid Asseblages with pitfall traps: The madness and the method. Can. Entomol. 1994, 126, 881–894. [Google Scholar] [CrossRef]
- Lang, A.-G. The pitfalls of pitfalls: A comparison of pitfall trap catches and absolute density estimates of epigeal invertebrate predators in arable land. J. Pest Sci. 2000, 73, 99–106. [Google Scholar]
- Hunter, M.D.; Price, P.W. Playing Chutes and Ladders : Heterogeneity and the relative roles of bottom-up and top-down forces in natural communities. Ecology 1992, 73, 724–732. [Google Scholar]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCabe, E.; Loeb, G.; Grab, H. Responses of Crop Pests and Natural Enemies to Wildflower Borders Depends on Functional Group. Insects 2017, 8, 73. https://doi.org/10.3390/insects8030073
McCabe E, Loeb G, Grab H. Responses of Crop Pests and Natural Enemies to Wildflower Borders Depends on Functional Group. Insects. 2017; 8(3):73. https://doi.org/10.3390/insects8030073
Chicago/Turabian StyleMcCabe, Ellie, Gregory Loeb, and Heather Grab. 2017. "Responses of Crop Pests and Natural Enemies to Wildflower Borders Depends on Functional Group" Insects 8, no. 3: 73. https://doi.org/10.3390/insects8030073
APA StyleMcCabe, E., Loeb, G., & Grab, H. (2017). Responses of Crop Pests and Natural Enemies to Wildflower Borders Depends on Functional Group. Insects, 8(3), 73. https://doi.org/10.3390/insects8030073