
Evidence of Tolerance to Silica-Based Desiccant Dusts in a Pyrethroid-Resistant Strain of Cimex lectularius (Hemiptera: Cimicidae)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Insects
2.2. Products
2.3. Desiccant Dust Assays
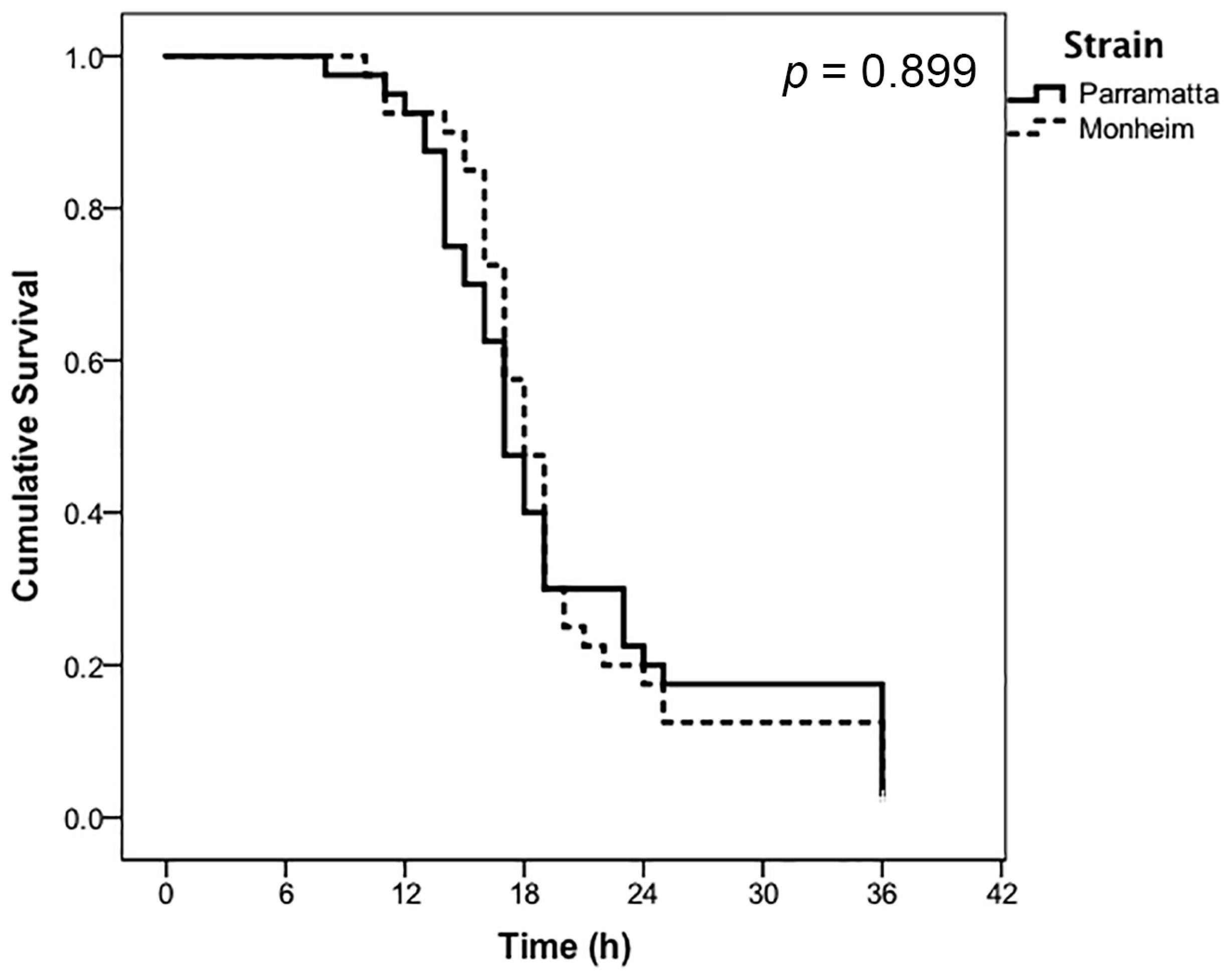
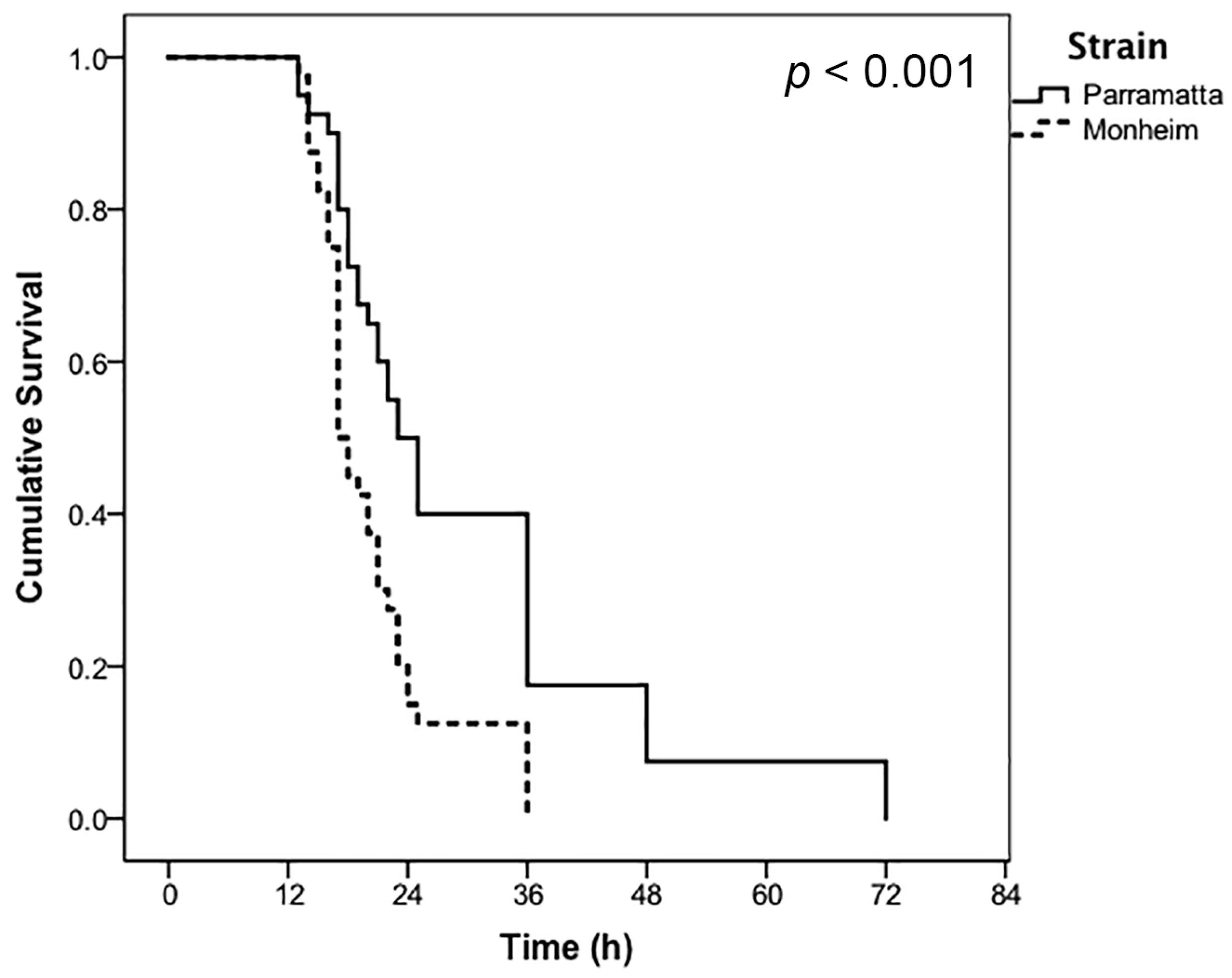
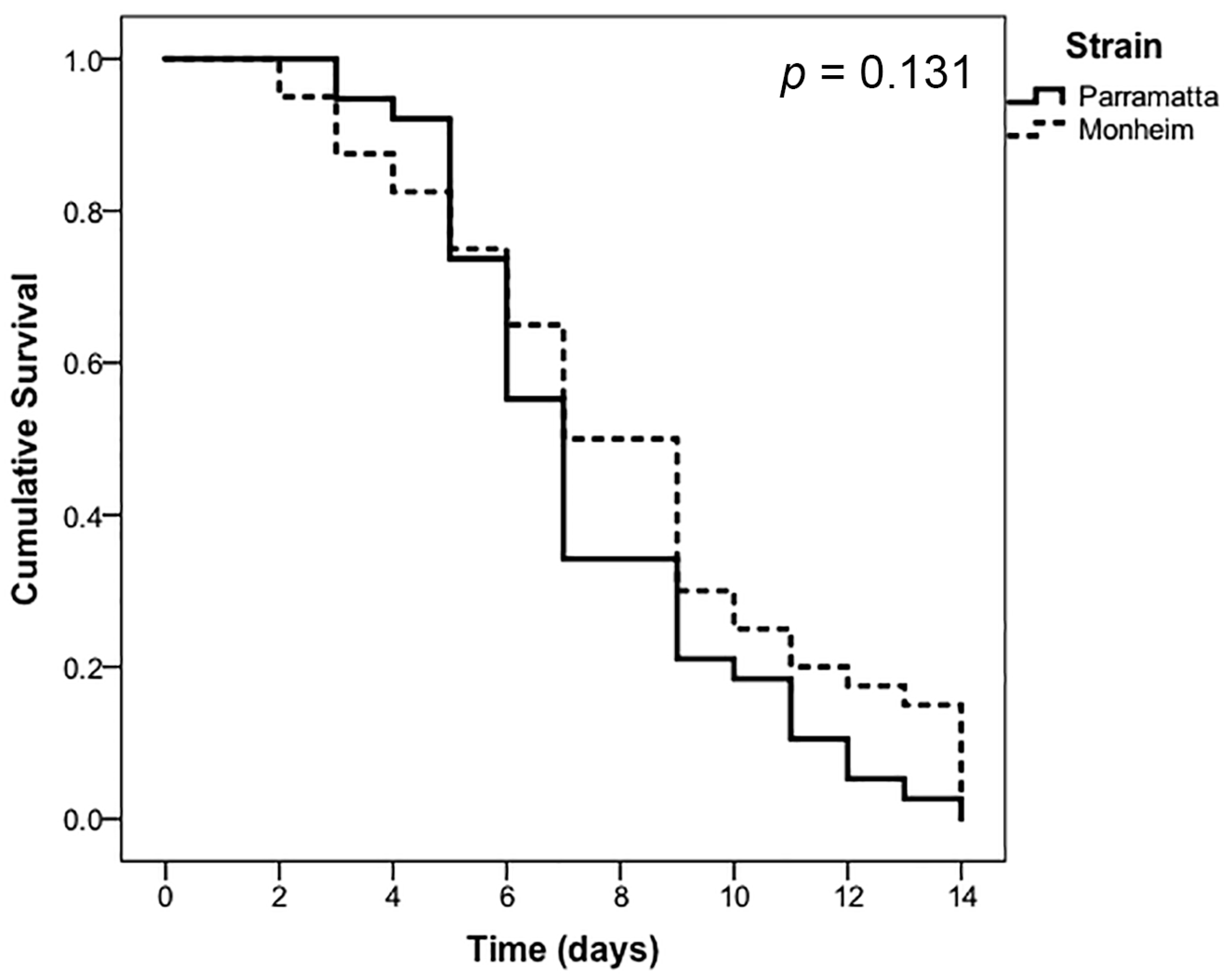
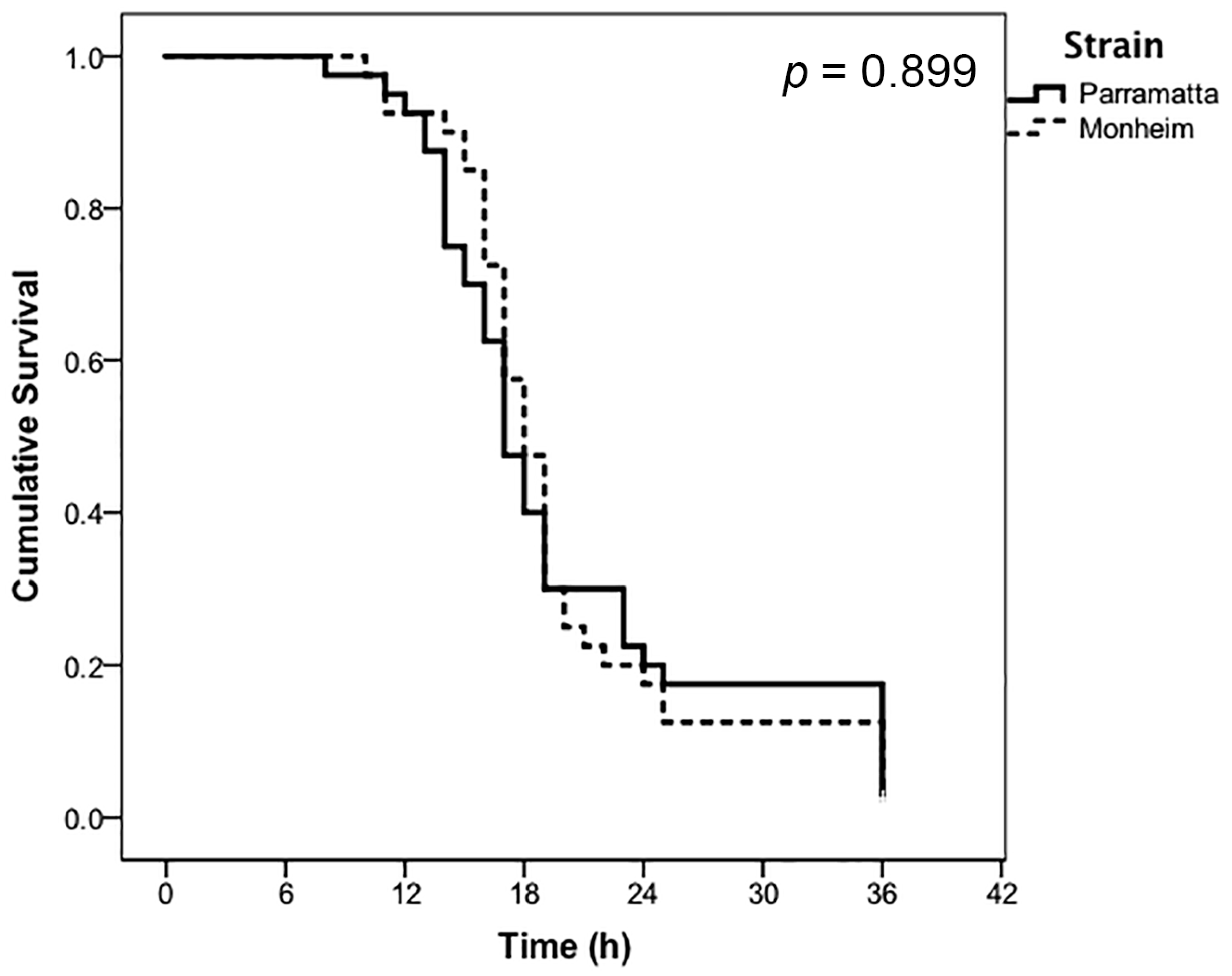
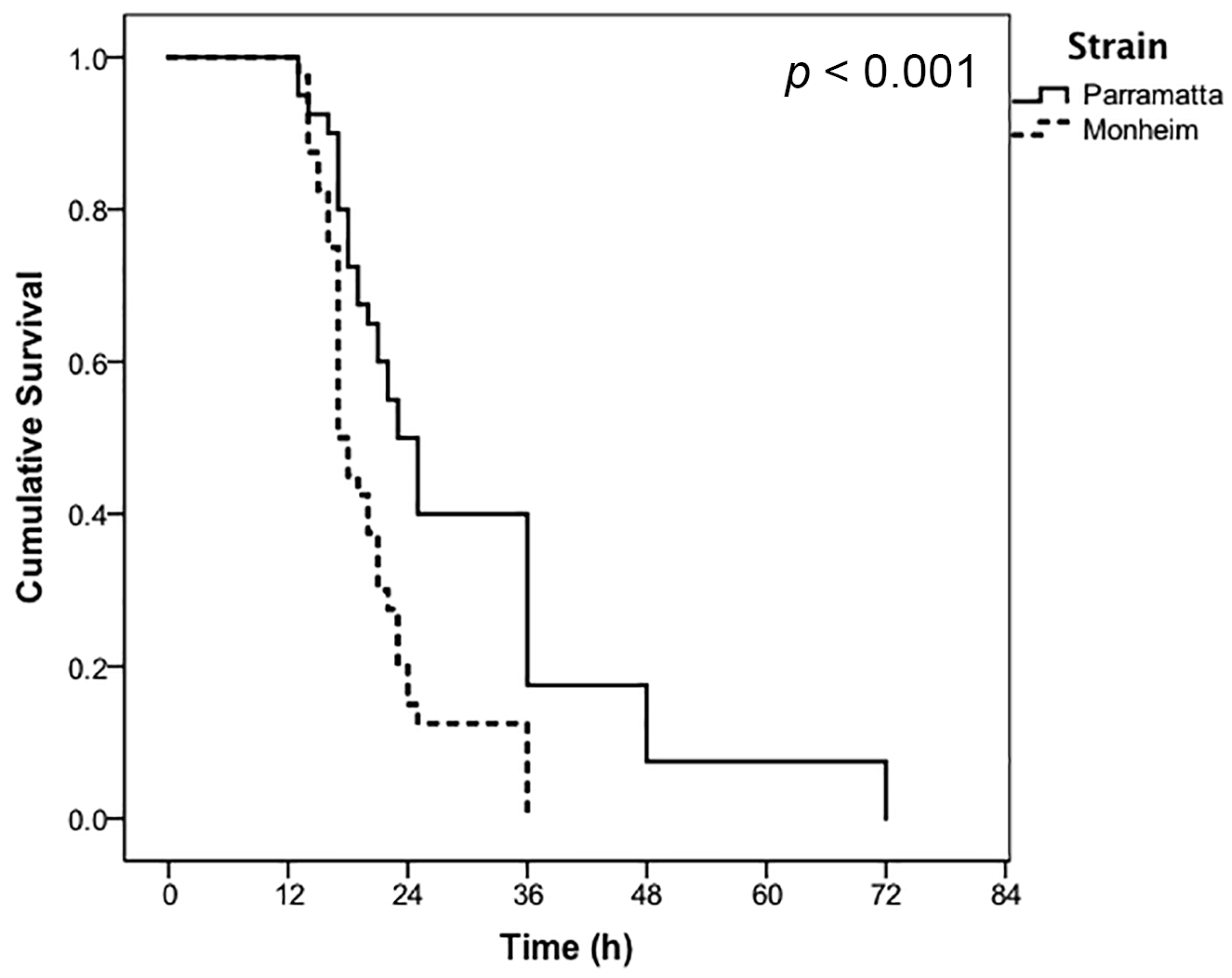
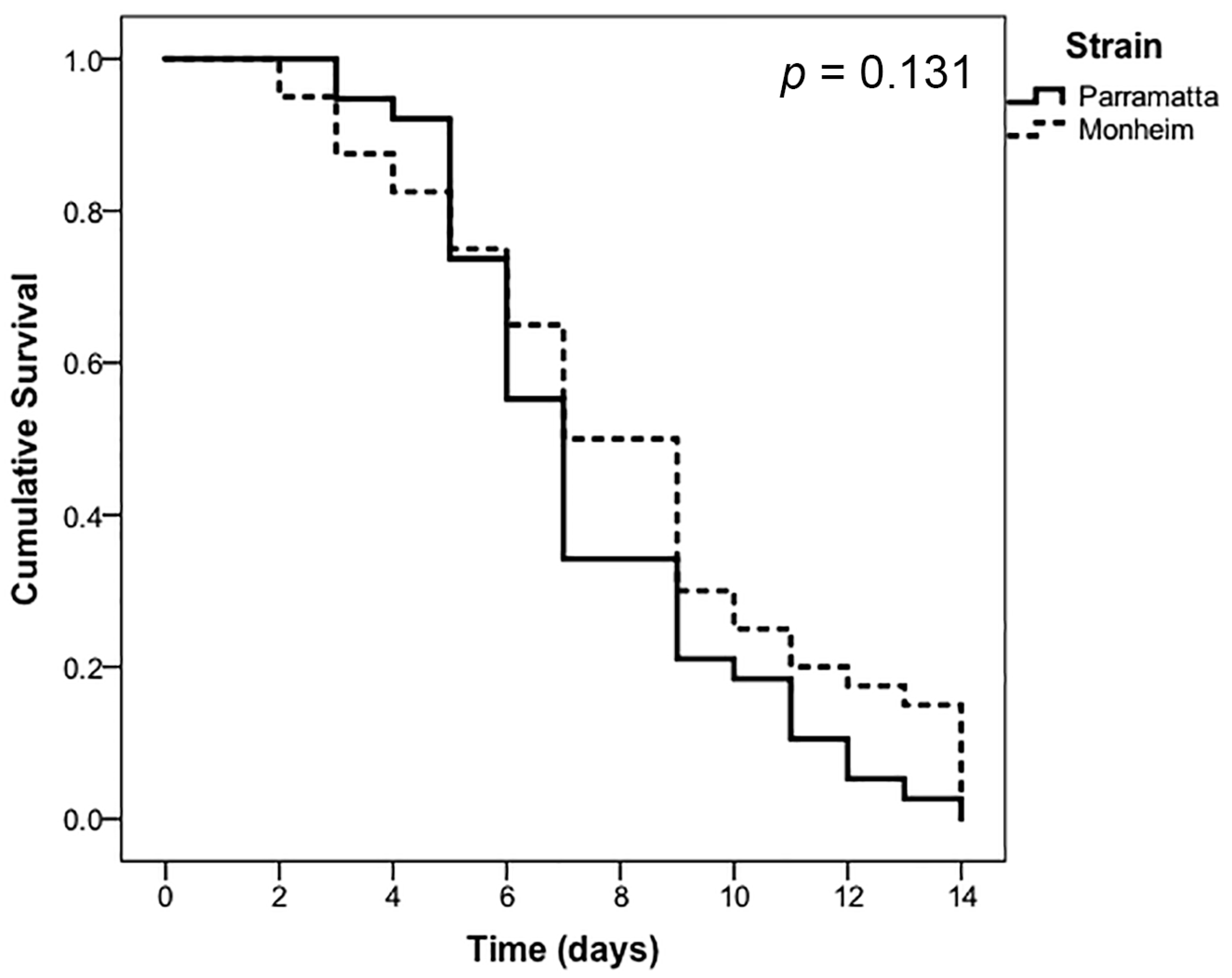
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Doggett, S.L.; Dwyer, D.E.; Peñas, P.F.; Russell, R.C. Bed bugs: Clinical relevance and control options. Clin. Microbiol. Rev. 2012, 25, 164–192. [Google Scholar] [CrossRef] [PubMed]
- Minocha, R.; Wang, C.; Dang, K.; Webb, C.E.; Peñas, P.F.; Doggett, S.L. Systemic and erythrodermic reactions following repeated exposure to bites from the Common bed bug, Cimex lectularius. Aust. Entomol. 2016, in press. [Google Scholar] [CrossRef]
- Doggett, S.L. A Code of Practice for the Control of Bed Bug Infestations in Australia, 4th ed.; Department of Medical Entomology and The Australian Environmental Pest Managers Association: Sydney, Australia, 2013; p. 44. [Google Scholar]
- Doggett, S.L.; Orton, C.J.; Lilly, D.G.; Russell, R.C. Bed bugs: The Australian response. Insects 2011, 2, 96–111. [Google Scholar] [CrossRef] [PubMed]
- Lilly, D.G.; Zalucki, M.P.; Orton, C.J.; Russell, R.C.; Webb, C.E.; Doggett, S.L. Confirmation of insecticide resistance in Cimex lectularius (Hemiptera: Cimicidae) in Australia. Aust. Entomol. 2015, 54, 96–99. [Google Scholar] [CrossRef]
- Romero, A.; Anderson, T.D. High levels of resistance in the common bed bug, Cimex lectularius (Hemiptera: Cimicidae), to neonicotinoid insecticides. J. Med. Entomol. 2016, 53, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Kilpinen, O.; Kristensen, M.; Vagn Jensen, K.M. Resistance differences between chlorpyrifos and synthetic pyrethroids in Cimex lectularius population from Denmark. Parasitol. Res. 2011, 109, 1461–1464. [Google Scholar] [CrossRef] [PubMed]
- Karunaratne, S.H.P.P.; Damayanthi, B.T.; Fareena, M.H.J.; Imbuldeniya, V.; Hemingway, J. Insecticide resistance in the tropical bedbug Cimex hemipterus. Pestic. Biochem. Physiol. 2007, 88, 102–107. [Google Scholar] [CrossRef]
- Suwannayod, S.; Chanbang, Y.; Buranapanichpan, S. The life cycle and effectiveness of insecticides against the bed bugs of Thailand. Southeast Asian J. Trop. 2010, 41, 548–554. [Google Scholar]
- Tawatsin, A.; Thavara, U.; Chompoosri, J.; Phusup, Y.; Jonjang, N.; Khumsawads, C.; Bhakdeenuan, P.; Sawanpanyalert, P.; Asavadachanukorn, P.; Mulla, M.S.; et al. Insecticide resistance in bedbugs in Thailand and laboratory evaluation of insecticides for the control of Cimex hemipterus and Cimex lectularius (Hemiptera: Cimicidae). J. Med. Entomol. 2011, 48, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Adelman, Z.N.; Kilcullen, K.A.; Koganemaru, R.; Anderson, M.A.E.; Anderson, T.D.; Miller, D.M. Deep sequencing of pyrethroid-resistant bed bugs reveals multiple mechanisms of resistance within a single population. PLoS ONE 2011, 6, e26228. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.F.; Cowles, R.S. Susceptibility of Cimex lectularius (Hemiptera: Cimicidae) to pyrethroid insecticides and to insecticidal dusts with or without pyrethroid insecticides. J. Econ. Entomol. 2012, 105, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Dang, K.; Toi, C.S.; Lilly, D.G.; Bu, W.; Doggett, S.L. Detection of knockdown resistance (kdr) mutations in the common bed bug, Cimex lectularius (Hemiptera: Cimicidae) in Australia. Pest. Manag. Sci. 2015, 71, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Dang, K.; Toi, C.S.; Lilly, D.G.; Lee, C.Y.; Naylor, R.; Tawatsin, A.; Thavara, U.; Bu, W.; Doggett, S.L. Identification of putative kdr mutations in the tropical bed bug, Cimex hemipterus (Hemiptera: Cimicidae). Pest. Manag. Sci. 2015, 71, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.R.; Goodman, M.H.; Potter, M.F.; Haynes, K.F. Population variation in and selection for resistance to pyrethroid-neonicotinoid insecticides in the bed bug. Sci. Rep. 2014, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Koganemaru, R.; Miller, D.M.; Adelman, Z.N. Robust cuticular penetration resistance in the common bed bug (Cimex lectularius L.) correlates with increased steady-state transcript levels of CPR-type cuticle protein genes. Pestic. Biochem. Physiol. 2013, 106, 190–197. [Google Scholar]
- Myamba, J.; Maxwell, C.A.; Asidi, A.; Curtis, C.F. Pyrethroid resistance in tropical bedbugs, Cimex hemipterus, associated with use of treated bednets. Med. Vet. Entomol. 2002, 16, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Potter, M.F.; Potter, D.A.; Haynes, K.F. Insecticide resistance in the bed bug: A factor in the pest’s sudden resurgence? J. Med. Entomol. 2007, 44, 175–178. [Google Scholar] [PubMed]
- Zhu, F.; Gujar, H.; Gordon, J.R.; Haynes, K.F.; Potter, M.F.; Palli, S.R. Bed bugs evolved unique adaptive strategy to resist pyrethroid insecticides. Sci. Rep. 2013. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Sams, S.; Moural, T.; Haynes, K.F.; Potter, M.F.; Palli, S.R. RNA interference of NADPH-cytochrome P450 reductase results in reduced insecticide resistance in the bed bug, Cimex lectularius. PLoS ONE 2012, 7, e31037. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Wigginton, J.; Romero, A.; Moore, A.; Ferguson, K.; Palli, R.; Potter, M.F.; Haynes, K.F.; Palli, S.R. Widespread distribution of knockdown resistance mutations in the bed bug, Cimex lectularius (Hemiptera: Cimicidae), populations in the United States. Arch. Insect Biochem. 2010, 73, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.J.; Miller, D.M. Laboratory evaluations of insecticide product efficacy for control of Cimex lectularius. J. Econ. Entomol. 2006, 99, 2080–2086. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.E.; Miller, D.M. Insecticide resistance in eggs and first instars of the bed bug, Cimex lectularius (Hemiptera: Cimicidae). Insects 2015, 6, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Doggett, S.L.; Russell, R.C. The resurgence of bed bugs, Cimex spp. (Hemiptera: Cimicidae) in Australia. In Proceedings of the Sixth International Conference on Urban Pests, Budapest, Hungary, 13–16 July 2008; Robinson, W.H., Bajomi, D., Eds.; OOK-Press: Veszprém, Budapest, Hungary, 2008; pp. 407–425. [Google Scholar]
- Wang, C.L.; Saltzmann, K.; Chin, E.; Bennett, G.W.; Gibb, T. Characteristics of Cimex lectularius (Hemiptera: Cimicidae), infestation and dispersal in a high-rise apartment building. J. Econ. Entomol. 2010, 103, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.F.; Haynes, K.F.; Romero, A.; Hardebeck, E.; Wickemeyer, W. Is there a new bed bug answer? Pest Control Technol. 2008, 36, 118–124. [Google Scholar]
- Singh, N.; Wang, C.L.; Cooper, R. Effectiveness of a reduced-risk insecticide based bed bug management program in low-income housing. Insects 2013, 4, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.F.; Haynes, K.F.; Fredericks, J. Bed bugs across America: The 2015 bugs without borders survey. Pestworld 2015, 7, 4–14. [Google Scholar]
- Wang, C.L.; Gibb, T.; Bennett, G.W. Evaluation of two least toxic integrated pest management programs for managing bed bugs (Heteroptera: Cimicidae) with discussion of a bed bug intercepting device. J. Med. Entomol. 2009, 46, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Saltzmann, K.; Bennett, G.; Gibb, T. Comparison of three bed bug management strategies in a low-income apartment building. Insects 2012, 3, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Singh, N.; Cooper, R.A.; Liu, C.F.; Buczkowski, G. Evaluation of an insecticide dust band treatment method for controlling bed bugs. J. Econ. Entomol. 2013, 106, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Ebeling, W. Sorptive dusts for pest control. Annu. Rev. Entomol. 1971, 16, 123–158. [Google Scholar] [CrossRef] [PubMed]
- Golob, P. Current status and future perspectives for inert dusts for control of stored product insects. J. Stored Prod. Res. 1997, 33, 69–79. [Google Scholar] [CrossRef]
- Quarles, W.; Winn, P.S. Diatomaceous earth and stored product pests. IPM Pract. 1996, 18, 1–10. [Google Scholar]
- Subramanyam, B.; Roesli, R. Inert dusts. In Alternatives to Pesticides in Stored-Product IPM; Subramanyam, B., Hagstrum, D.W., Eds.; Springer: Boston, MA, USA, 2000; pp. 321–380. [Google Scholar]
- Ebeling, W. Wood destroying insects and fungi. In Urban Entomology; University of California, Division of Agriculture Science: Berkeley, CA, USA, 1975; pp. 128–167. [Google Scholar]
- McLaughlin, A. Laboratory trials on desiccant dust insecticides. In Proceedings of the 6th International Working Conference on Stored-Product Protection, Canberra, Australia, 17–23 April 1994; Highley, E., Wright, E.J., Banks, H.J., Champ, B.R., Eds.; CAB International: Wallingford, UK, 1994; pp. 638–645. [Google Scholar]
- Prasantha, B.D.R.; Reichmuth, C.; Adler, C.; Felgentreu, D. Lipid adsorption of diatomaceous earths and increased water permeability in the epicuticle layer of the cowpea weevil Callosobruchus maculatus (F.) and the bean weevil Acanthoscelides obtectus (Say) (Chrysomelidae). J. Stored Prod. Res. 2015, 64, 36–41. [Google Scholar] [CrossRef]
- Korunic, Z. Diatomaceous earths, a group of natural insecticides. J. Stored Prod. Res. 1998, 34, 87–97. [Google Scholar] [CrossRef]
- Malia, H.A.E.; Rosi-Denadai, C.A.; Guedes, N.M.P.; Martins, G.F.; Guedes, R.N.C. Diatomaceous earth impairment of water balance in the maize weevil, Sitophilus zeamais. J. Pest Sci. 2016, 89, 945–954. [Google Scholar] [CrossRef]
- Akhtar, Y.; Isman, M.B. Efficacy of diatomaceous earth and a DE-aerosol formulation against the common bed bug, Cimex lectularius Linnaeus in the laboratory. J. Pest. Sci. 2016, 89, 1013–1021. [Google Scholar] [CrossRef]
- Benoit, J.B.; Phillips, S.A.; Croxall, T.J.; Christensen, B.S.; Yoder, J.A.; Denlinger, D.L. Addition of alarm pheromone components improves the effectiveness of desiccant dusts against Cimex lectularius. J. Med. Entomol. 2009, 46, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Donahue, W.A., Jr.; Showler, A.T.; Donahue, M.W.; Vinson, B.E.; Hui, L.; Osbrink, W.L.A. Knockdown and lethal effects of eight commercial nonconventional and two pyrethroid insecticides against moderately permethrin-resistant adult bed bugs, Cimex lectularius (L.)(Hemiptera: Cimicidae). Biopestic. Int. 2015, 11, 108–117. [Google Scholar]
- Goddard, J. Long-term efficacy of various natural or “green” insecticides against bed bugs: A double-blind study. Insects 2014, 5, 942–951. [Google Scholar] [PubMed]
- Potter, M.F.; Haynes, K.F.; Gordon, J.R.; Washburn, L.; Washburn, M.; Hardin, T. Silica gel: A better bed bug dessicant. Pest Control Technol. 2014, 42, 76–84. [Google Scholar]
- Romero, A.; Potter, M.F.; Haynes, K.F. Are dusts the bed bug bullet. Pest Manag. Prof. 2009, 77, 22–30. [Google Scholar]
- Singh, N.; Wang, C.; Wang, D.; Cooper, R.; Zha, C. Comparative efficacy of selected dust insecticides for controlling Cimex lectularius (Hemiptera: Cimicidae). J. Econ. Entomol. 2016, 109, 1819–1826. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.F.; Haynes, K.F.; Christensen, C.; Neary, T.J.; Turner, C.R.; Washburn, L.; Washburn, M. Where do bed bugs stand when the dust settles? Pest Control Technol. 2013, 41, 72–78. [Google Scholar]
- Aak, A.; Roligheten, E.; Rukke, B.A.; Birkemoe, T. Desiccant dust and the use of CO2 gas as a mobility stimulant for bed bugs: A potential control solution? J. Pest Sci. 2016. [Google Scholar] [CrossRef]
- Lilly, D.G.; Latham, S.L.; Webb, C.E.; Doggett, S.L. Cuticle thickening in a pyrethroid-resistant strain of the common bed bug, Cimex lectularius L. (Hemiptera: Cimicidae). PLoS ONE 2016, 11, e0153302. [Google Scholar] [CrossRef] [PubMed]
- Usinger, R.L. Monograph of Cimicidae (Hemiptera–Heteroptera); Entomological Society of America: College Park, MD, USA, 1966. [Google Scholar]
- Lilly, D.G.; Dang, K.; Webb, C.E.; Doggett, S.L. Evidence for metabolic pyrethroid resistance in the common bed bug (Hemiptera: Cimicidae). J. Econ. Entomol. 2016, 109, 1364–1368. [Google Scholar] [CrossRef] [PubMed]
- Dang, K.; Lilly, D.G.; Bu, W.; Doggett, S.L. Simple, rapid and cost-effective technique for the detection of pyrethroid resistance in bed bugs, Cimex spp. (Hemiptera: Cimicidae). Aust. Entomol. 2015, 54, 191–196. [Google Scholar]
- WHO. Test Procedures for Insecticide Resistance Monitoring, Bio-Efficacy and Persistence of Insecticides on Treated Surfaces; WHO/CDS/CPC/MAL/98.12; World Health Organization: Geneva, Switzerland, 1998; pp. 1–43. [Google Scholar]
- Yu, S.J. Insecticide resistance. In The Toxicology and Biochemistry of Insecticides, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 257–321. [Google Scholar]
- Benoit, J.B.; Del Grosso, N.A.; Yoder, J.A.; Denlinger, D.L. Resistance to dehydration between bouts of blood feeding in the bed bug, Cimex lectularius, is enhanced by water conservation, aggregation, and quiescence. Am. J. Trop. Med. Hyg. 2007, 76, 987–993. [Google Scholar] [PubMed]
- Rigaux, M.; Haubruge, E.; Fields, P.G. Mechanisms for tolerance to diatomaceous earth between strains of Tribolium castaneum. Entomol. Exp. Appl. 2001, 101, 33–39. [Google Scholar] [CrossRef]
- Ebeling, W. Inorganic insecticides and dusts. In Understanding and Controlling the German Cockroach; Rust, M.K., Owens, J.M., Reierson, D.A., Eds.; Oxford University Press: Oxford, UK; New York, NY, USA, 1995; pp. 193–230. [Google Scholar]
- Akhtar, Y.; Isman, M.B. Horizontal transfer of diatomaceous earth and botanical insecticides in the common bed bug, Cimex lectularius L.; Hemiptera: Cimicidae. PLoS ONE 2013, 8, e75626. [Google Scholar] [CrossRef] [PubMed]
- Polanco, A.M.; Miller, D.M.; Brewster, C.C. Survivorship during starvation for Cimex lectularius L. Insects 2011, 2, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, A.G.; Rajpurohit, S. Cuticular lipids and water balance. In Insect Hydrocarbons: Biology, Biochemistry, and Chemical Ecology; Blomquist, G.J., Bagnères, A.-G., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2010; pp. 100–120. [Google Scholar]
- Chown, S.L.; Gibbs, A.G.; Hetz, S.K.; Klok, C.J.; Lighton, J.R.B.; Marais, E. Discontinuous gas exchange in insects: A clarification of hypotheses and approaches. Physiol. Biochem. Zool. 2006, 79, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, M.C.; Gibbs, A.G. Discontinuous gas exchange in insects. Respir. Physiol. 2006, 154, 18–29. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Strain | Treatment | Label Rate | Replicate | Label Rate Dose (g) | Actual Dose (g) | Deviation (g) | Deviation (%) |
|---|---|---|---|---|---|---|---|
| Parramatta | CimeXa | 0.5× | 1 | 0.0194 | 0.0195 | 0.0001 | 0.52% |
| 2 | 0.0194 | 0.0195 | 0.0001 | 0.52% | |||
| 3 | 0.0194 | 0.0197 | 0.0003 | 1.55% | |||
| 4 | 0.0194 | 0.0197 | 0.0003 | 1.55% | |||
| CimeXa | 1.0× | 1 | 0.0388 | 0.0387 | −0.0001 | −0.26% | |
| 2 | 0.0388 | 0.0390 | 0.0002 | 0.52% | |||
| 3 | 0.0388 | 0.0390 | 0.0002 | 0.52% | |||
| 4 | 0.0388 | 0.0394 | 0.0006 | 1.55% | |||
| Bed Bug Killer | 1.0× | 1 | 0.0254 | 0.0253 | −0.0001 | −0.39% | |
| 2 | 0.0254 | 0.0253 | −0.0001 | −0.39% | |||
| 3 | 0.0254 | 0.0249 | −0.0005 | −1.97% | |||
| 4 | 0.0254 | 0.0254 | 0.0000 | 0.00% | |||
| Monheim | CimeXa | 0.5× | 1 | 0.0194 | 0.0194 | 0.0000 | 0.00% |
| 2 | 0.0194 | 0.0195 | 0.0001 | 0.52% | |||
| 3 | 0.0194 | 0.0192 | −0.0002 | −1.03% | |||
| 4 | 0.0194 | 0.0190 | −0.0004 | −2.06% | |||
| CimeXa | 1.0× | 1 | 0.0388 | 0.0393 | 0.0005 | 1.29% | |
| 2 | 0.0388 | 0.0386 | −0.0002 | −0.52% | |||
| 3 | 0.0388 | 0.0390 | 0.0002 | 0.52% | |||
| 4 | 0.0388 | 0.0383 | −0.0005 | −1.29% | |||
| Bed Bug Killer | 1.0× | 1 | 0.0254 | 0.0255 | 0.0001 | 0.39% | |
| 2 | 0.0254 | 0.0254 | 0.0000 | 0.00% | |||
| 3 | 0.0254 | 0.0255 | 0.0001 | 0.39% | |||
| 4 | 0.0254 | 0.0257 | 0.0003 | 1.18% |
| Treatment | Label Rate | Strain | LT50 (h) | 95% CI (Lower) | 95% CI (Upper) |
|---|---|---|---|---|---|
| CimeXa | 0.5× | Parramatta | 30.00 * | 25.07 | 34.93 |
| 0.5× | Monheim | 20.47 | 18.40 | 22.55 | |
| 1.0× | Parramatta | 20.17 | 17.65 | 22.70 | |
| 1.0× | Monheim | 20.00 | 17.86 | 22.14 |
| Treatment | Label Rate | Strain | LT50 (Days) | 95% CI (Lower) | 95% CI (Upper) |
|---|---|---|---|---|---|
| Bed Bug Killer | 1.0× | Parramatta | 7.42 | 6.54 | 8.30 |
| 1.0× | Monheim | 8.12 | 6.99 | 9.25 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lilly, D.G.; Webb, C.E.; Doggett, S.L. Evidence of Tolerance to Silica-Based Desiccant Dusts in a Pyrethroid-Resistant Strain of Cimex lectularius (Hemiptera: Cimicidae). Insects 2016, 7, 74. https://doi.org/10.3390/insects7040074
Lilly DG, Webb CE, Doggett SL. Evidence of Tolerance to Silica-Based Desiccant Dusts in a Pyrethroid-Resistant Strain of Cimex lectularius (Hemiptera: Cimicidae). Insects. 2016; 7(4):74. https://doi.org/10.3390/insects7040074
Chicago/Turabian StyleLilly, David G., Cameron E. Webb, and Stephen L. Doggett. 2016. "Evidence of Tolerance to Silica-Based Desiccant Dusts in a Pyrethroid-Resistant Strain of Cimex lectularius (Hemiptera: Cimicidae)" Insects 7, no. 4: 74. https://doi.org/10.3390/insects7040074
APA StyleLilly, D. G., Webb, C. E., & Doggett, S. L. (2016). Evidence of Tolerance to Silica-Based Desiccant Dusts in a Pyrethroid-Resistant Strain of Cimex lectularius (Hemiptera: Cimicidae). Insects, 7(4), 74. https://doi.org/10.3390/insects7040074