
Sodium Channel Mutations and Pyrethroid Resistance in Aedes aegypti

Abstract
:1. Introduction
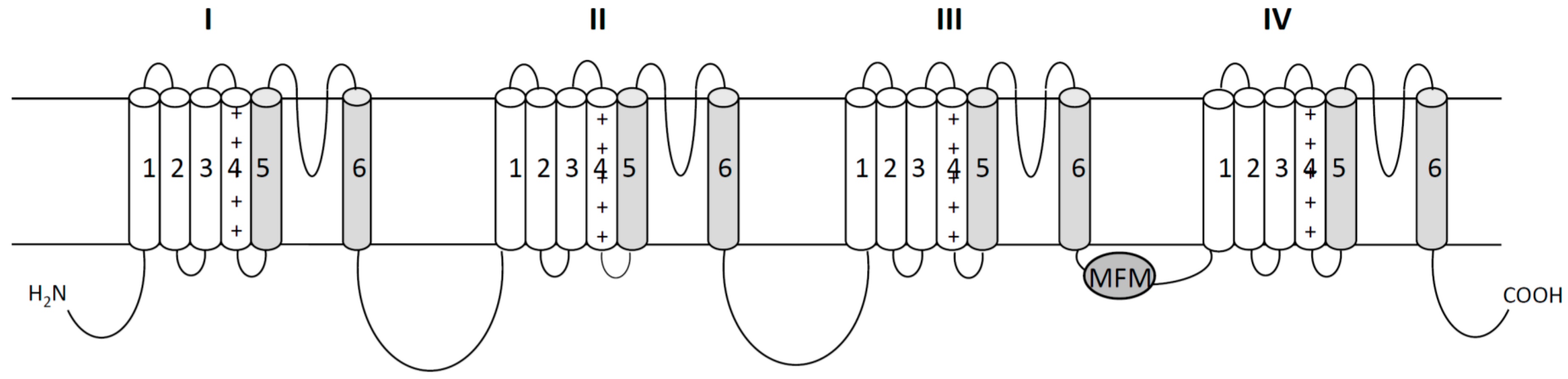
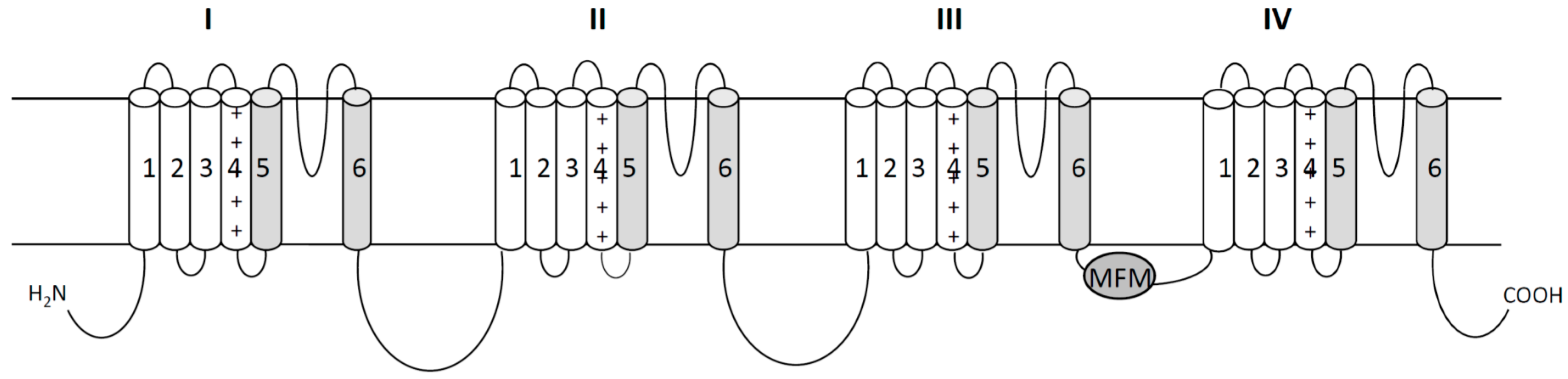
2. Voltage Gated Sodium Channels as Targets of Pyrethroid Insecticides
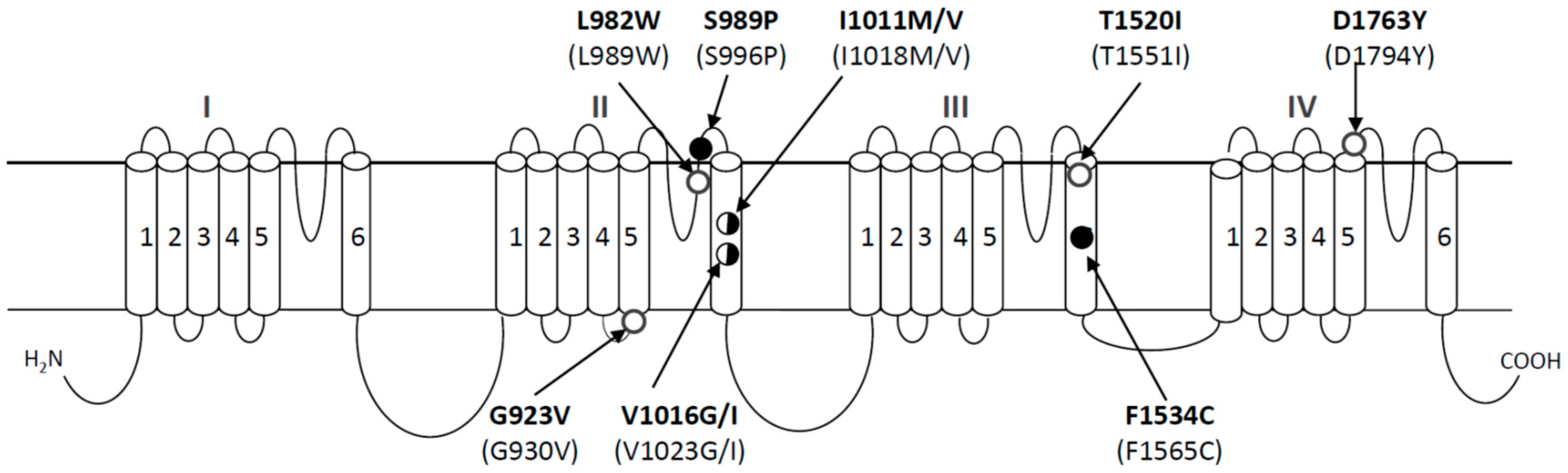
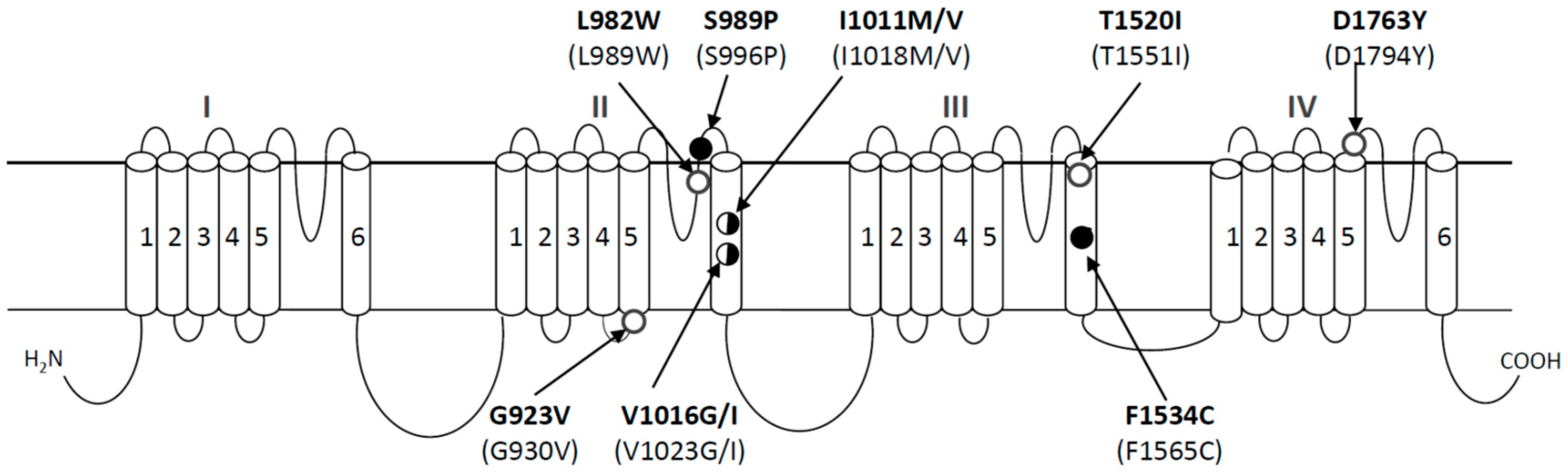
3. Identification of Single Nucleotide Polymorphisms (SNPs) in the Sodium Channel That Are Associated with Pyrethroid Resistance
4. Functional Conformation of kdr Mutations in Xenopus Oocytes
5. Evolution of kdr Mutations in Aedes aegypti
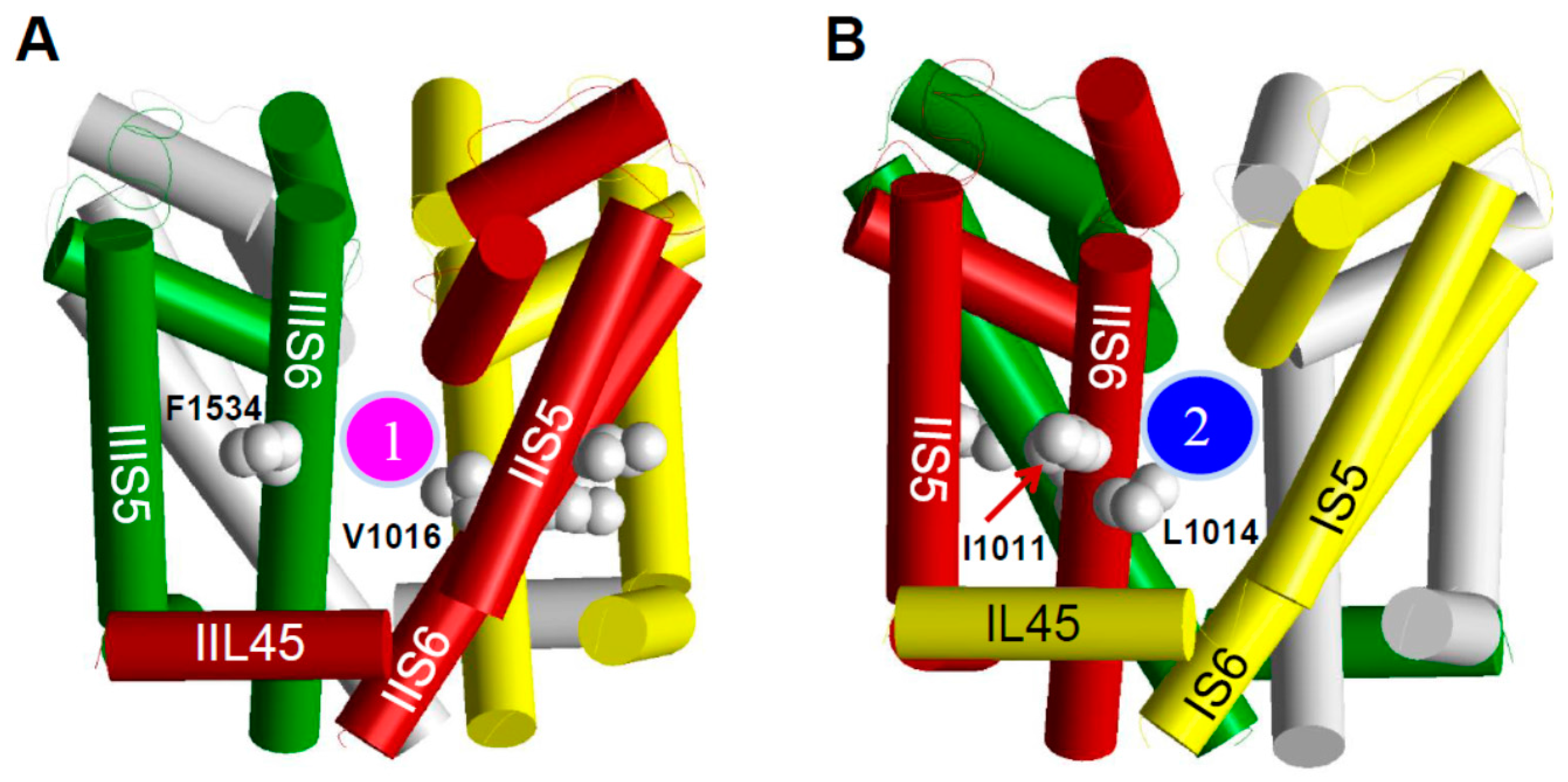
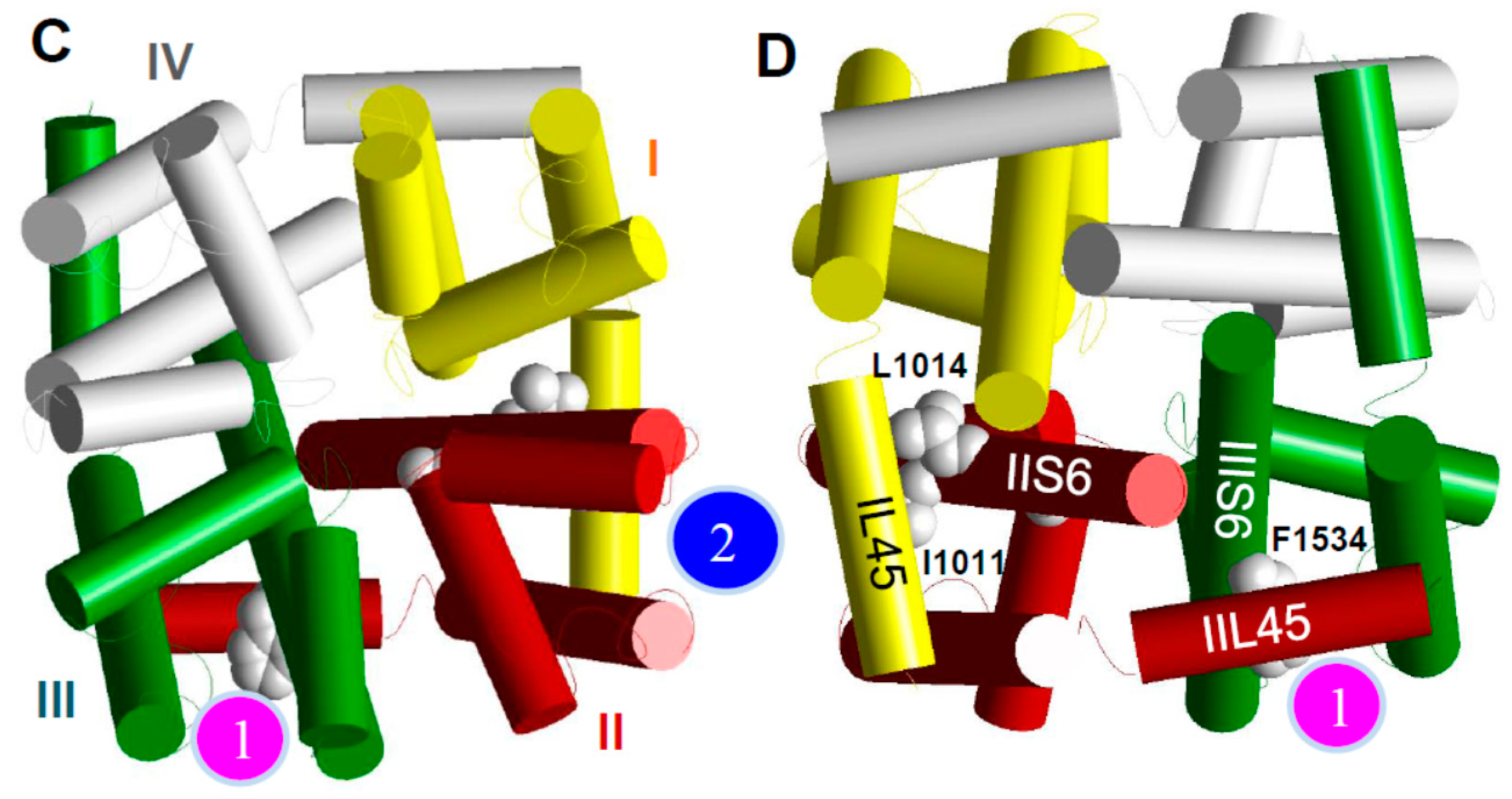
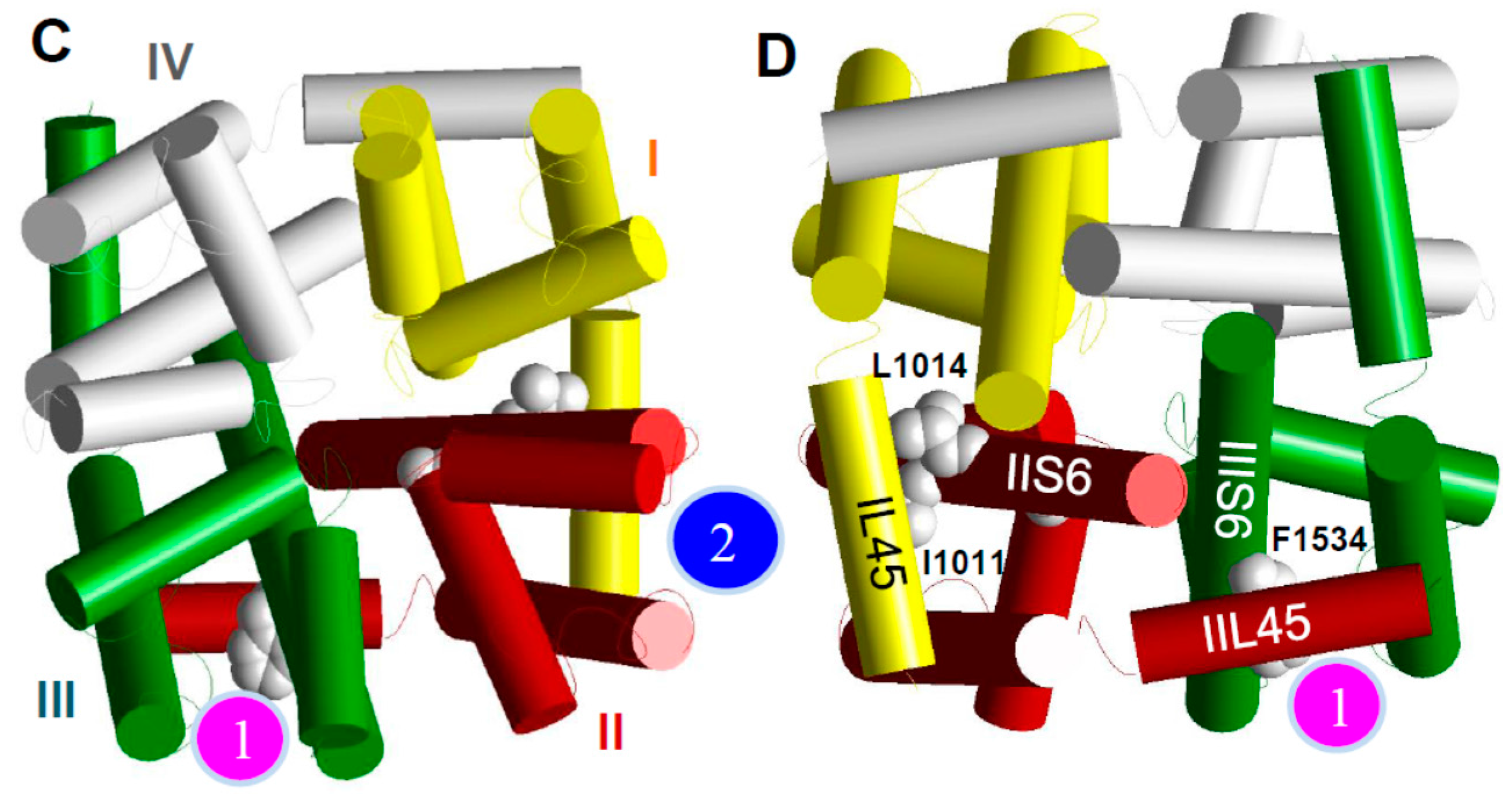
6. The kdr Mutations from Aedes aegypti Likely Confer Resistance to Pyrethroids by Reducing Pyrethroid Binding to One of the Two Pyrethroid Receptor Sites on the Sodium Channel
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Elliott, M. Synthetic Pyrethroids; American Chemical Society: Washington, DC, USA, 1977; pp. 1–28. [Google Scholar]
- Ranson, H.; Lissenden, N. Insecticide resistance in african Anopheles mosquitoes: A worsening situation that needs urgent action to maintain malaria control. Trends Parasitol. 2016, 32, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.; Du, Y.; Rinkevich, F.; Nomura, Y.; Xu, P.; Wang, L.; Silver, K.; Zhorov, B.S. Molecular biology of insect sodium channels and pyrethroid resistance. Insect Biochem. Mol. Biol. 2014, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Narahashi, T.; Ginsburg, K.S.; Nagata, K.; Song, J.H.; Tatebayashi, H. Ion channels as targets for insecticides. Neurotoxicology 1998, 19, 581–590. [Google Scholar] [PubMed]
- Silver, K.S.; Du, Y.; Nomura, Y.; Oliveira, E.E.; Salgado, V.L.; Zhorov, B.S.; Dong, K. Voltage-Gated Sodium Channels as Insecticide Target Sites; Elsevier: Burlington, MA, USA, 2014; Volume 46, pp. 389–433. [Google Scholar]
- Soderlund, D.M. Sodium channels. In Comprehensive Molecular Insect Science; Gilbert, L.I., Iatrou, K., Gill, S.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; Volume 5, pp. 1–24. [Google Scholar]
- Soderlund, D.M.; Bloomquist, J.R. Neurotoxic actions of pyrethroid insecticides. Ann. Rev. Entomol. 1989, 34, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Rinkevich, F.D.; Du, Y.; Dong, K. Diversity and convergence of sodium channel mutations involved in resistance to pyrethroids. Pestic. Biochem. Physiol. 2013, 106, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.B.; Kasai, S.; Scott, J.G. Pyrethroid resistance in Aedes aegypti and Aedes albopictus: Important mosquito vectors of human diseases. Pestic. Biochem. Physiol. 2016, 133, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Soderlund, D.M. Molecular mechanisms of pyrethroid insecticide neurotoxicity: Recent advances. Arch. Toxicol. 2012, 86, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Narahashi, T. Neuronal ion channels as the target sites of insecticides. Pharmacol. Toxicol. 1996, 78, 1–14. [Google Scholar] [CrossRef]
- Catterall, W.A. Structure and function of voltage-gated sodium channels at atomic resolution. Exp. Physiol. 2014, 99, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, M.; Ohyama, K.; Dunlap, D.Y.; Matsumura, F. Cloning and sequencing of the para-type sodium channel gene from susceptible and kdr-resistant german cockroaches (Blattella germanica) and house fly (Musca domestica). Mol. Gen. Genet. 1996, 252, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.S.; Martinez-Torres, D.; Hick, C.A.; Devonshire, A.L. Identification of mutations in the housefly para-type sodium channel gene associated with knockdown resistance (kdr) to pyrethroid insecticides. Mol. Gen. Genet. 1996, 252, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Dong, K. A single amino acid change in the para sodium channel protein is associated with knockdown-resistance (kdr) to pyrethroid insecticides in german cockroach. Insect Biochem. Mol. Biol. 1997, 27, 93–100. [Google Scholar] [CrossRef]
- Kim, H.; Baek, J.H.; Lee, W.-J.; Lee, S.-H. Frequency detection of pyrethroid resistance allele in Anopheles sinensis populations by real-time PCR amplification of specific allele (rtpasa). Pestic. Biochem. Physiol. 2007, 87, 54–61. [Google Scholar] [CrossRef]
- Lol, J.C.; Castellanos, M.E.; Liebman, K.A.; Lenhart, A.; Pennington, P.M.; Padilla, N.R. Molecular evidence for historical presence of knock-down resistance in Anopheles albimanus, a key malaria vector in Latin America. Parasites Vectors 2013. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Li, C.; Wang, Z.; Liu, M.; Dong, Y.; Feng, X.; Wu, Z.; Guo, X.; Xing, D.; Zhang, Y. First detection of multiple knockdown resistance (kdr)-like mutations in voltage-gated sodium channel using three new genotyping methods in Anopheles sinensis from guangxi province, China. J. Med. Entomol. 2012, 49, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, C.; Xing, D.; Yu, Y.; Liu, N.; Xue, R.; Dong, Y.; Zhao, T. Detection and widespread distribution of sodium channel alleles characteristic of insecticide resistance in Culex pipiens complex mosquitoes in China. Med. Vet. Entomol. 2012, 26, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhong, D.; Zhang, D.; Shi, L.; Zhou, G.; Gong, M.; Zhou, H.; Sun, Y.; Ma, L.; He, J. Molecular ecology of pyrethroid knockdown resistance in Culex pipiens pallens mosquitoes. PLoS ONE 2010, 5, e11681. [Google Scholar] [CrossRef] [PubMed]
- Enayati, A.A.; Vatandoost, H.; Ladonni, H.; Townson, H.; Hemingway, J. Molecular evidence for a kdr-like pyrethroid resistance mechanism in the malaria vector mosquito anopheles stephensi. Med. Vet. Entomol. 2003, 17, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Karunaratne, S.; Hawkes, N.J.; Perera, M.D.B.; Ranson, H.; Hemingway, J. Mutated sodium channel genes and elevated monooxygenases are found in pyrethroid resistant populations of Sri Lankan malaria vectors. Pestic. Biochem. Physiol. 2007, 88, 108–113. [Google Scholar] [CrossRef]
- Luleyap, H.U.; Alptekin, D.; Kasap, H.; Kasap, M. Detection of knockdown resistance mutations in Anopheles sacharovi (diptera: Culicidae) and genetic distance with Anopheles gambiae (diptera: Culicidae) using cdna sequencing of the voltage-gated sodium channel gene. J. Med. Entomol. 2002, 39, 870–874. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Torres, D.; Chandre, F.; Williamson, M.S.; Darriet, F.; Berge, J.B.; Devonshire, A.L.; Guillet, P.; Pasteur, N.; Pauron, D. Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae s.s. Insect Mol. Biol. 1998, 7, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Torres, D.; Chevillon, C.; Brun-Barale, A.; Bergé, J.B.; Pasteur, N.; Pauron, D. Voltage-dependent Na+ channels in pyrethroid-resistant Culex pipiens L. mosquitoes. Pestic. Sci. 1999, 55, 1012–1020. [Google Scholar] [CrossRef]
- Singh, O.P.; Dykes, C.L.; Das, M.K.; Pradhan, S.; Bhatt, R.M.; Agrawal, O.P.; Adak, T. Presence of two alternative kdr-like mutations, L1014F and L1014S, and a novel mutation, V1010L, in the voltage gated Na+ channel of Anopheles culicifacies from Orissa, India. Malar J. 2010. [Google Scholar] [CrossRef] [PubMed]
- Stump, A.D.; Atieli, F.K.; Vulule, J.M.; Besansky, N.J. Dynamics of the pyrethroid knockdown resistance allele in western kenyan populations of Anopheles gambiae in response to insecticide-treated bed net trials. Am. J. Trop. Med. Hyg. 2004, 70, 591–596. [Google Scholar] [PubMed]
- Verhaeghen, K.; van Bortel, W.; Trung, H.D.; Sochantha, T.; Keokenchanh, K.; Coosemans, M. Knockdown resistance in Anopheles vagus, An. Sinensis, An. Paraliae and An. Peditaeniatus populations of the Mekong region. Parasites Vectors 2010. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Liu, H.; Zhang, L.; Liu, N. Resistance in the mosquito, Culex quinquefasciatus, and possible mechanisms for resistance. Pest Manag. Sci. 2005, 61, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Ranson, H.; Jensen, B.; Vulule, J.M.; Wang, X.; Hemingway, J.; Collins, F.H. Identification of a point mutation in the voltage-gated sodium channel gene of Kenyan Anopheles gambiae associated with resistance to DDT and pyrethroids. Insect Mol. Biol. 2000, 9, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Brengues, C.; Hawkes, N.J.; Chandre, F.; McCarroll, L.; Duchon, S.; Guillet, P.; Manguin, S.; Morgan, J.C.; Hemingway, J. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med. Vet. Entomol. 2003, 17, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Saavedra-Rodriguez, K.; Urdaneta-Marquez, L.; Rajatileka, S.; Moulton, M.; Flores, A.E.; Fernandez-Salas, I.; Bisset, J.; Rodriguez, M.; McCall, P.J.; Donnelly, M.J. A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol. Biol. 2007, 16, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Kawada, H.; Higa, Y.; Komagata, O.; Kasai, S.; Tomita, T.; Thi Yen, N.; Loan, L.L.; Sanchez, R.A.; Takagi, M. Widespread distribution of a newly found point mutation in voltage-gated sodium channel in pyrethroid-resistant Aedes aegypti populations in Vietnam. PLoS Negl. Trop. Dis. 2009, 3, e527. [Google Scholar] [CrossRef] [PubMed]
- Yanola, J.; Somboon, P.; Walton, C.; Nachaiwieng, W.; Somwang, P.; Prapanthadara, L.A. High-throughput assays for detection of the F1534C mutation in the voltage-gated sodium channel gene in permethrin-resistant Aedes aegypti and the distribution of this mutation throughout Thailand. Trop. Med. Int. Health 2011, 16, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Yanola, J.; Somboon, P.; Prapanthadara, L. A novel point mutation in the Aedes aegypti voltage-gated sodium channel gene associated with permethrin resistance. In Proceedings of the 2nd International Conference on Dengue and Dengue Haemorhagic Fever, Phuket, Thailand, 15–17 October 2008.
- Yanola, J.; Somboon, P.; Walton, C.; Nachaiwieng, W.; Prapanthadara, L.-A. A novel F1552/C1552 point mutation in the Aedes aegypti voltage-gated sodium channel gene associated with permethrin resistance. Pestic. Biochem. Physiol. 2010, 96, 127–131. [Google Scholar] [CrossRef]
- Srisawat, R.; Komalamisra, N.; Eshita, Y.; Zheng, M.; Ono, K.; Itoh, T.Q.; Matsumoto, A.; Petmitr, S.; Rongsriyam, Y. Point mutations in domain II of the voltage-gated sodium channel gene in deltamethrin-resistant Aedes aegypti (diptera: Culicidae). Appl. Entomol. Zool. 2010, 45, 275–282. [Google Scholar] [CrossRef]
- Srisawat, R.; Komalamisra, N.; Apiwathnasorn, C.; Paeporn, P.; Roytrakul, S.; Rongsriyam, Y.; Eshita, Y. Field-collected permethrin-resistant Aedes aegypti from central thailand contain point mutations in the domain IIS6 of the sodium channel gene (kdr). Southeast Asian J. Trop. Med. Public Health 2012, 43, 1380–1386. [Google Scholar] [PubMed]
- Stenhouse, S.A.; Plernsub, S.; Yanola, J.; Lumjuan, N.; Dantrakool, A.; Choochote, W.; Somboon, P. Detection of the V1016G mutation in the voltage-gated sodium channel gene of Aedes aegypti (diptera: Culicidae) by allele-specific PCR assay, and its distribution and effect on deltamethrin resistance in Thailand. Parasites Vectors 2013. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Shen, W.K.; Wang, T.T.; Lin, Y.H.; Hsu, E.L.; Dai, S.M. A novel amino acid substitution in a voltage-gated sodium channel is associated with knockdown resistance to permethrin in Aedes aegypti. Insect Biochem. Mol. Biol. 2009, 39, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Ishak, I.H.; Jaal, Z.; Ranson, H.; Wondji, C.S. Contrasting patterns of insecticide resistance and knockdown resistance (kdr) in the dengue vectors Aedes aegypti and Aedes albopictus from Malaysia. Parasites Vectors 2015. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.C.; Chiang, L.P.; Tan, C.H.; Vythilingam, I.; Lam-Phua, S.G.; Ng, L.C. Low efficacy of delthamethrin-treated net against singapore Aedes aegypti is associated with kdr-type resistance. Trop. Biomed. 2015, 32, 140–150. [Google Scholar] [PubMed]
- Li, C.; Kaufman, P.; Xue, R.; Zhao, M.; Wang, G; Yan, T.; Guo, X.; Zhang, Y.; Dong, Y.; Xing, D. Relationship between insecticide resistance and kdr mutations in the dengue vector Aedes aegypti in southern China. Parasites Vectors 2015. [Google Scholar] [CrossRef] [PubMed]
- Kawada, H.; Oo, S.Z.; Thaung, S.; Kawashima, E.; Maung, Y.N.; Thu, H.M.; Thant, K.Z.; Minakawa, N. Co-occurrence of point mutations in the voltage-gated sodium channel of pyrethroid-resistant Aedes aegypti populations in Myanmar. PLoS Negl. Trop. Dis. 2014, 8, e3032. [Google Scholar] [CrossRef] [PubMed]
- Wuliandari, J.R.; Lee, S.F.; White, V.L.; Tantowijoyo, W.; Hoffmann, A.A.; Endersby-Harshman, N.M. Association between three mutations, F1565C, V1023G and S996P, in the voltage-sensitive sodium channel gene and knockdown resistance in Aedes aegypti from yogyakarta, Indonesia. Insects 2015, 6, 658–685. [Google Scholar] [CrossRef] [PubMed]
- Prapanthadara, L.-A.; Promtet, N.; Koottathep, S.; Somboon, P.; Suwonkerd, W.; McCarroll, L.; Hemingway, J. Mechanisms of DDT and permethrin resistance in Aedes aegypti from Chiang Mai, Thailand. Dengue Bull. 2002, 26, 185–189. [Google Scholar]
- Rajatileka, S.; Black, W.C.; Saavedra-Rodriguez, K.; Trongtokit, Y.; Apiwathnasorn, C.; McCall, P.J.; Ranson, H. Development and application of a simple colorimetric assay reveals widespread distribution of sodium channel mutations in Thai populations of Aedes aegypti. Acta Trop. 2008, 108, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, L.C.; Ponce, G.; Saavedra-Rodriguez, K.; Lopez, B.; Flores, A.E. Frequency of V1016I and F1534C mutations in the voltage-gated sodium channel gene in Aedes aegypti in Venezuela. Pest Manag. Sci. 2015, 71, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Dusfour, I.; Zorrilla, P.; Guidez, A.; Issaly, J.; Girod, R.; Guillaumot, L.; Robello, C.; Strode, C. Deltamethrin resistance mechanisms in Aedes aegypti populations from three french overseas territories worldwide. PLoS Negl. Trop. Dis. 2015, 9, e0004226. [Google Scholar] [CrossRef] [PubMed]
- Bona, A.C.; Chitolina, R.F.; Fermino, M.L.; de Castro Poncio, L.; Weiss, A.; Lima, J.B.; Paldi, N.; Bernardes, E.S.; Henen, J.; Maori, E. Larval application of sodium channel homologous dsRNA restores pyrethroid insecticide susceptibility in a resistant adult mosquito population. Parasites Vectors 2016, 9. [Google Scholar] [CrossRef] [PubMed]
- Chapadense, F.G.; Fernandes, E.K.; Lima, J.B.; Martins, A.J.; Silva, L.C.; Rocha, W.T.; Santos, A.H.; Cravo, P. Phenotypic and genotypic profile of pyrethroid resistance in populations of the mosquito Aedes aegypti from Goiania, central west Brazil. Rev. Soc. Bras. Med. Trop. 2015, 48, 607–609. [Google Scholar] [CrossRef] [PubMed]
- Linss, J.G.; Brito, L.P.; Garcia, G.A.; Araki, A.S.; Bruno, R.V.; Lima, J.B.; Valle, D.; Martins, A.J. Distribution and dissemination of the Val1016Ile and Phe1534Cys kdr mutations in Aedes aegypti brazilian natural populations. Parasites Vectors 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Aponte, H.A.l.; Penilla, R.P.; Dzul-Manzanilla, F.; Che-Mendoza, A.; López, A.D.; Solis, F.; Manrique-Saide, P.; Ranson, H.; Lenhart, A.; McCall, P.J. The pyrethroid resistance status and mechanisms in Aedes aegypti from the guerrero state, Mexico. Pestic. Biochem. Physiol. 2013, 107, 226–234. [Google Scholar] [CrossRef]
- Cornel, A.J.; Holeman, J.; Nieman, C.C.; Lee, Y.; Smith, C.; Amorino, M.; Brisco, K.K.; Barrera, R.; Lanzaro, G.C.; Mulligan, F.S., III. Surveillance, insecticide resistance and control of an invasive Aedes aegypti (diptera: Culicidae) population in California. F1000Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Kawada, H.; Higa, Y.; Futami, K.; Muranami, Y.; Kawashima, E.; Osei, J.H.; Sakyi, K.Y.; Dadzie, S.; de Souza, D.K.; Appawu, M. Discovery of point mutations in the voltage-gated sodium channel from african Aedes aegypti populations: Potential phylogenetic reasons for gene introgression. PLoS Negl. Trop. Dis. 2016, 10, e0004780. [Google Scholar] [CrossRef] [PubMed]
- Kushwah, R.B.; Dykes, C.L.; Kapoor, N.; Adak, T.; Singh, O.P. Pyrethroid-resistance and presence of two knockdown resistance (kdr) mutations, F1534C and a novel mutation T1520I, in Indian Aedes aegypti. PLoS Negl. Trop. Dis. 2015, 9, e3332. [Google Scholar] [CrossRef] [PubMed]
- Kasai, S.; Ng, L.C.; Lam-Phua, S.G.; Tang, C.S.; Itokawa, K.; Komagata, O.; Kobayashi, M.; Tomita, T. First detection of a putative knockdown resistance gene in major mosquito vector, Aedes albopictus. Jpn. J. Infect. Dis. 2011, 64, 217–221. [Google Scholar] [PubMed]
- Chen, H.; Li, K.; Wang, X.; Yang, X.; Lin, Y.; Cai, F.; Zhong, W.; Lin, C.; Lin, Z.; Ma, Y. First identification of kdr allele F1534S in VGSC gene and its association with resistance to pyrethroid insecticides in Aedes albopictus populations from Haikou city, Hainan island, China. Infect. Dis. Poverty 2016. [Google Scholar] [CrossRef] [PubMed]
- Marcombe, S.; Farajollahi, A.; Healy, S.P.; Clark, G.G.; Fonseca, D.M. Insecticide resistance status of united states populations of Aedes albopictus and mechanisms involved. PLoS ONE 2014, 9, e101992. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Valles, S.M.; Dong, K. Novel point mutations in the german cockroach para sodium channel gene are associated with knockdown resistance (kdr) to pyrethroid insecticides. Insect Biochem. Mol. Biol. 2000, 30, 991–997. [Google Scholar] [CrossRef]
- Hodgdon, H.E.; Yoon, K.S.; Previte, D.J.; Kim, H.J.; Aboelghar, G.E.; Lee, S.H.; Clark, J.M. Determination of knockdown resistance allele frequencies in global human head louse populations using the serial invasive signal amplification reaction. Pest Manag. Sci. 2010, 66, 1031–1040. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.; Liyanapathirana, M.; Agossa, F.R.; Weetman, D.; Ranson, H.; Donnelly, M.J.; Wilding, C.S. Footprints of positive selection associated with a mutation (N1575Y) in the voltage-gated sodium channel of Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2012, 109, 6614–6619. [Google Scholar] [CrossRef] [PubMed]
- Head, D.; McCaffery, A.; Callaghan, A. Novel mutations in the para-homologous sodium channel gene associated with phenotypic expression of nerve insensitivity resistance to pyrethroids in Heliothine lepidoptera. Insect Mol. Biol. 1998, 7, 191–196. [Google Scholar] [CrossRef]
- Hu, Z.; Du, Y.; Nomura, Y.; Dong, K. A sodium channel mutation identified in Aedes aegypti selectively reduces cockroach sodium channel sensitivity to type I, but not type II pyrethroids. Insect Biochem. Mol. Biol. 2011, 41, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Vera-Maloof, F.Z.; Saavedra-Rodriguez, K.; Elizondo-Quiroga, A.E.; Lozano-Fuentes, S.; Black Iv, W.C. Coevolution of the Ile1,016 and Cys1,534 mutations in the voltage gated sodium channel gene of Aedes aegypti in Mexico. PLoS Negl. Trop. Dis. 2015, 9, e0004263. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Nomura, Y.; Satar, G.; Hu, Z.; Nauen, R.; He, S.Y.; Zhorov, B.S.; Dong, K. Molecular evidence for dual pyrethroid-receptor sites on a mosquito sodium channel. Proc. Natl. Acad. Sci. USA 2013, 110, 11785–11790. [Google Scholar] [CrossRef] [PubMed]
- Hirata, K.; Komagata, O.; Itokawa, K.; Yamamoto, A.; Tomita, T.; Kasai, S. A single crossing-over event in voltage-sensitive Na+ channel genes may cause critical failure of dengue mosquito control by insecticides. PLoS Negl. Trop. Dis. 2014, 8, e3085. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.F.; Rajatileka, S.; Ranson, H. Pyrethroid resistance in Aedes aegypti from Grand Cayman. Am. J. Trop. Med. Hyg. 2010, 83, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Nomura, Y.; Zhorov, B.S.; Dong, K. Evidence for dual binding sites for 1,1,1-trichloro-2,2-bis(p-chlorophenyl)ethane (DDT) in insect sodium channels. J. Biol. Chem. 2016, 291, 4638–4648. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Smith, T.J.; Knipple, D.C.; Soderlund, D.M. Mutations in the house fly Vssc1 sodium channel gene associated with super-kdr resistance abolish the pyrethroid sensitivity of Vssc1/tipE sodium channels expressed in xenopus oocytes. Insect Biochem. Mol. Biol. 1999, 29, 185–194. [Google Scholar] [PubMed]
- Du, Y.; Lee, J.E.; Nomura, Y.; Zhang, T.; Zhorov, B.S.; Dong, K. Identification of a cluster of residues in transmembrane segment 6 of domain III of the cockroach sodium channel essential for the action of pyrethroid insecticides. Biochem. J. 2009, 419, 377–385. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, A.O.; Khambay, B.P.; Williamson, M.S.; Field, L.M.; Wallace, B.A.; Davies, T.G. Modelling insecticide-binding sites in the voltage-gated sodium channel. Biochem. J. 2006, 396, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Usherwood, P.N.; Davies, T.G.; Mellor, I.R.; O'Reilly, A.O.; Peng, F.; Vais, H.; Khambay, B.P.; Field, L.M.; Williamson, M.S. Mutations in DIIS5 and the DIIS4-S5 linker of drosophila melanogaster sodium channel define binding domains for pyrethroids and DDT. FEBS Lett. 2007, 581, 5485–5492. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Nomura, Y.; Zhorov, B.S.; Dong, K. Rotational symmetry of two pyrethroid receptor sites in the mosquito sodium channel. Mol. Pharmacol. 2015, 88, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Zhorov, B.S.; Dong, K. Elucidation of pyrethroid and DDT receptor sites in the voltage-gated sodium channel. Neurotoxicology 2016, in press. [Google Scholar] [CrossRef] [PubMed]
- Vais, H.; Williamson, M.S.; Goodson, S.J.; Devonshire, A.L.; Warmke, J.W.; Usherwood, P.N.; Cohen, C.J. Activation of drosophila sodium channels promotes modification by deltamethrin. Reductions in affinity caused by knock-down resistance mutations. J. Gen. Physiol. 2000, 115, 305–318. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutation 1 | Original Numbering 2 | Year 3 | Country | Reference |
|---|---|---|---|---|
| L982W | L75W | 2003 | Vietnam | [31] |
| I1011M + G923V | I104M + G16V | 2003 | Brazil, Guyana, and Martinique | [31] |
| I1011V | 2007 | Multiple Latin American Thailand | [32] [47] | |
| V1016G | V109G | 2003 | Indonesia | [31] |
| Thailand | [31,46,47] | |||
| V1016G + S989P | 2010 | Thailand | [37,38,39] | |
| Malaysia | [41] | |||
| V1016G + F1534C | 2015 | Singapore | [42] | |
| V1016G + F1534C + S989P | 2011 | Thailand | [34] | |
| Myanmar | [44] | |||
| Indonesia | [45] | |||
| China | [43] | |||
| V1016G + D1763Y | D1794Y | 2009 | Taiwan | [40] |
| V1016I | Multiple Latin American | [32] | ||
| F1534C | F1269C | 2008 | Vietnam | [33] |
| F1552C | Thailand | [34,35,36] | ||
| F1534C + V1016I | 2013 | Venezuela | [48] | |
| French overseas territories | [49] | |||
| Brazil | [50,51,52] | |||
| Mexico | [53] | |||
| United States | [54] | |||
| Ghana | [55] | |||
| F1534C + T1520I | 2015 | India | [56] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.; Nomura, Y.; Zhorov, B.S.; Dong, K. Sodium Channel Mutations and Pyrethroid Resistance in Aedes aegypti. Insects 2016, 7, 60. https://doi.org/10.3390/insects7040060
Du Y, Nomura Y, Zhorov BS, Dong K. Sodium Channel Mutations and Pyrethroid Resistance in Aedes aegypti. Insects. 2016; 7(4):60. https://doi.org/10.3390/insects7040060
Chicago/Turabian StyleDu, Yuzhe, Yoshiko Nomura, Boris S. Zhorov, and Ke Dong. 2016. "Sodium Channel Mutations and Pyrethroid Resistance in Aedes aegypti" Insects 7, no. 4: 60. https://doi.org/10.3390/insects7040060
APA StyleDu, Y., Nomura, Y., Zhorov, B. S., & Dong, K. (2016). Sodium Channel Mutations and Pyrethroid Resistance in Aedes aegypti. Insects, 7(4), 60. https://doi.org/10.3390/insects7040060