Color Difference and Memory Recall in Free-Flying Honeybees: Forget the Hard Problem


Abstract
:1. Introduction
2. Experimental Section
2.1. General Conditions
2.1.1. Group 1 Testing with Similar Colors
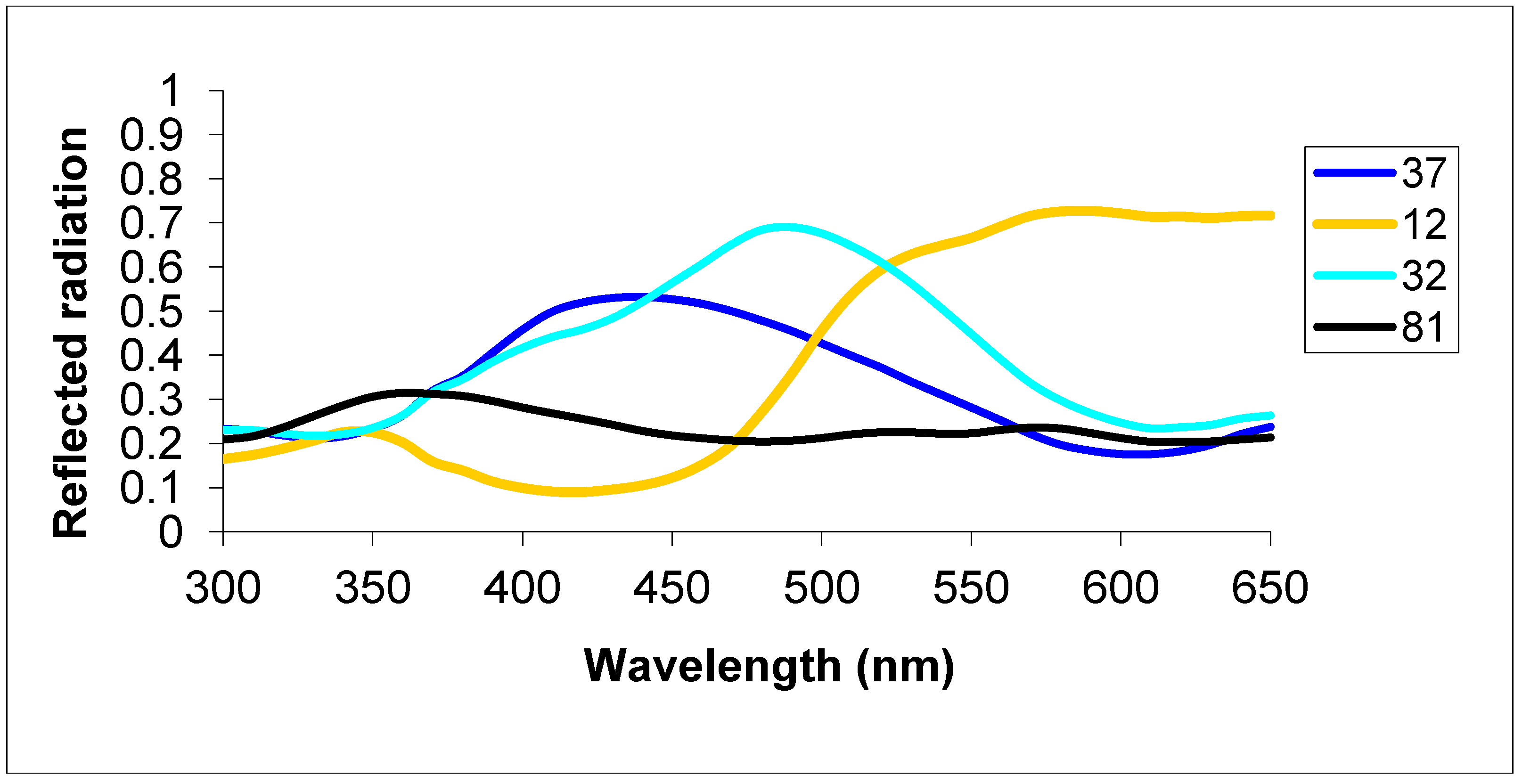
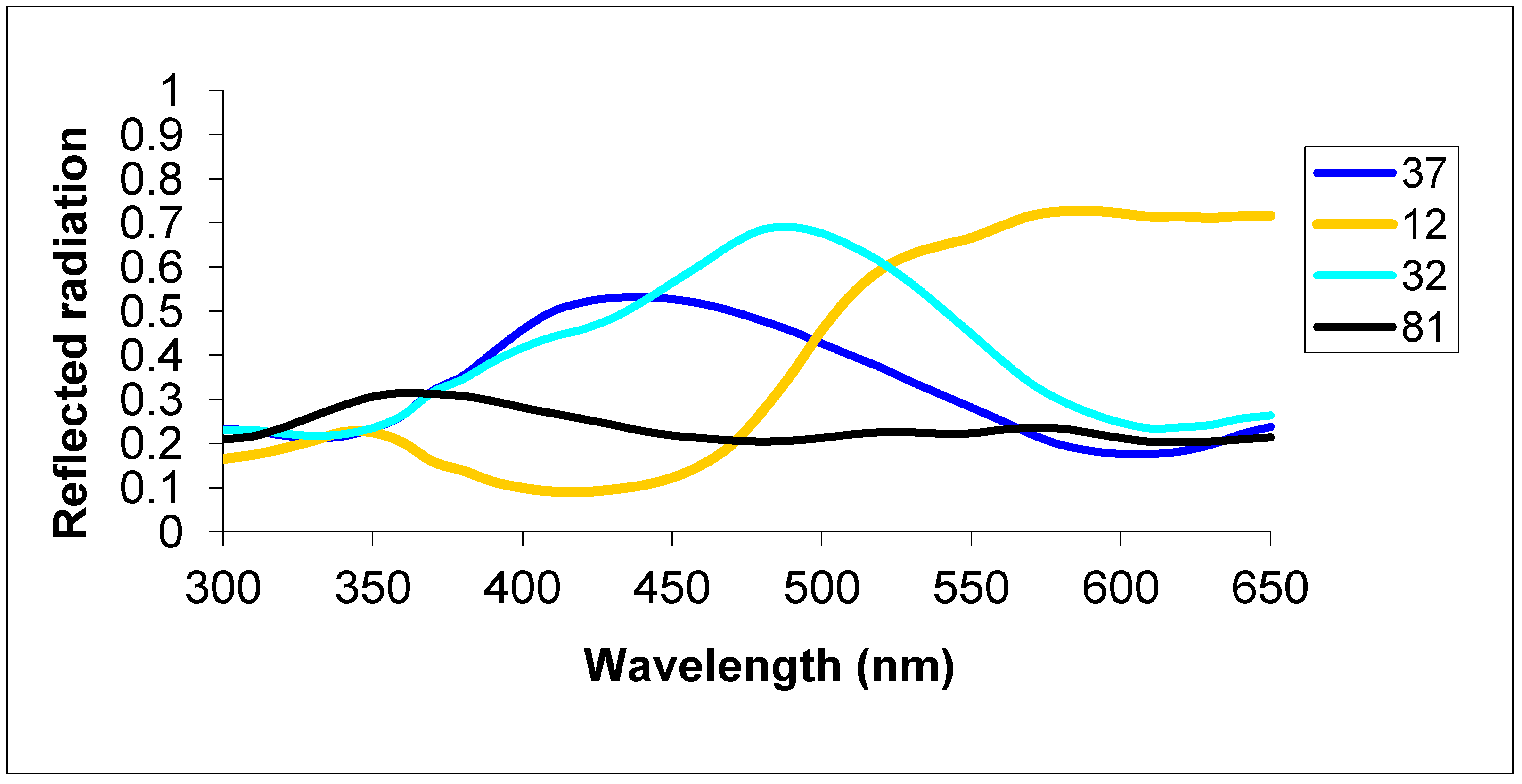
2.1.2. Group 2 Testing with Dissimilar Colors
3. Results and Discussion
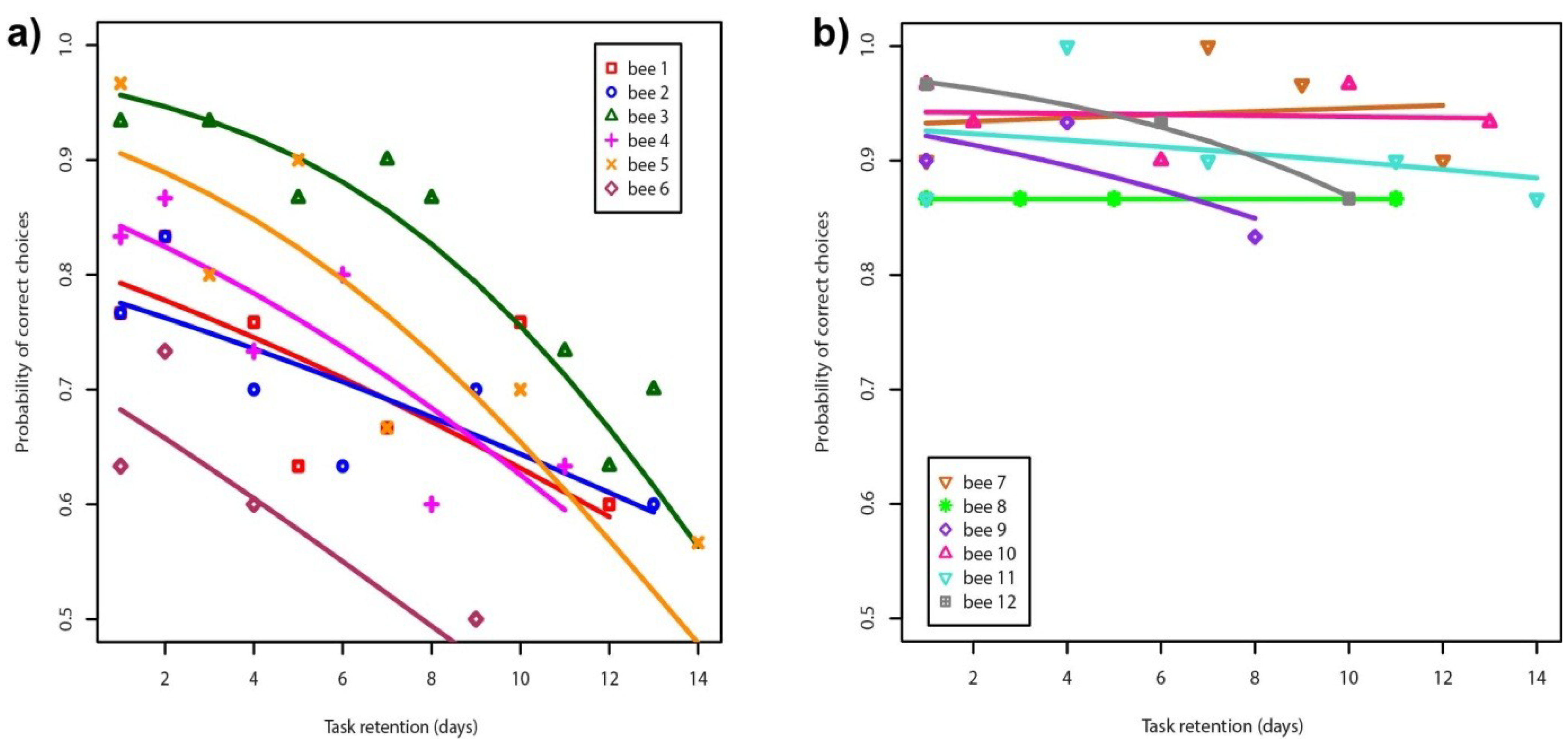

{kind=link}
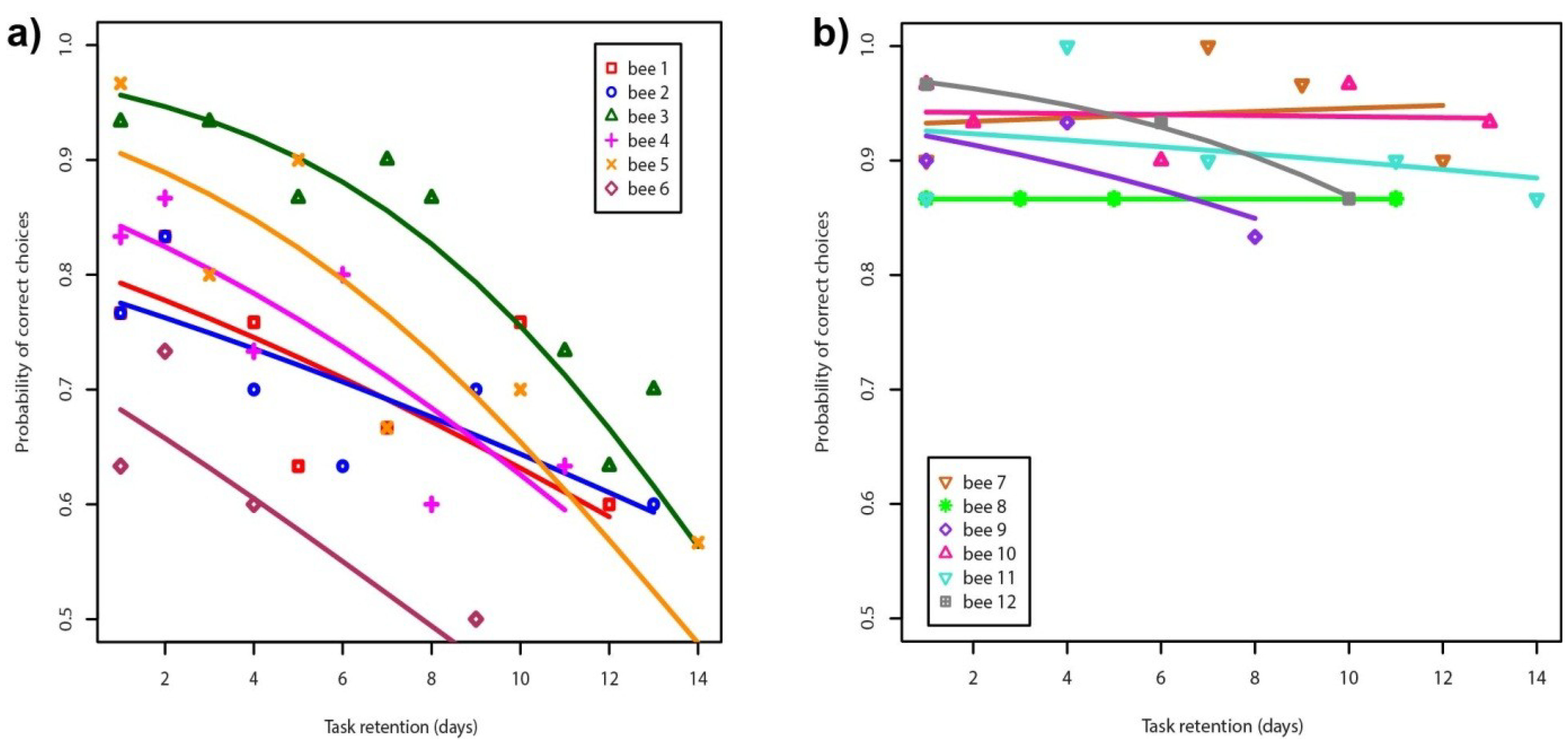{kind=link}
| Bee | Intercept | Task Retention (Days) † | p-Value § |
|---|---|---|---|
| Hard task | |||
| 1 (7 days) | 1.32 (0.75, 0.192) | −0.06 (−0.14, 0.015) | 0.111 |
| 2 (6 days) | 1.31 (0.74, 1.91) | −0.07 (−0.15, 0.00) | 0.066 |
| 3 (9 days) | 3.31 (2.44, 4.33) | −0.22 (−0.31, −0.14) | <0.000 ** |
| 4 (6 days) | 1.80 (1.15, 2.53) | −0.13 (−0.23, −0.03) | 0.01 ** |
| 5 (7 days) | 2.44 (1.72, 3.26) | −0.18 (−0.26, −0.10) | <0.000 ** |
| 6 (5 days) | 0.88 (0.27, 1.51) | −0.11 (−0.22, −0.00) | 0.04 ** |
| Easy task | |||
| 7 (4 days) | 2.60 (1.31, 4.39) | 0.02 (−0.17, 0.21) | 0.802 |
| 8 (4 days) | 1.87 (1.04, 2.82) | 5.11 × 10−17 (0.14, 0.15) | 1.000 |
| 9 (3 days) | 2.57 (1.37, 4.1) | −0.10 (−0.35, 0.12) | 0.371 |
| 10 (5 days) | 2.80 (1.76, 4.15) | −0.01 (−0.16, 0.14) | 0.916 |
| 11 (5 days) | 2.56 (1.56, 3.80) | −0.04 (−0.16, 0.08) | 0.532 |
| 12 (3 days) | 3.60 (1.97, 6.10) | −0.17 (−0.45, 0.05) | 0.142 |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends. Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Fenster, C.B.; Armbruster, W.S.; Wilson, P.; Dudash, M.R.; Thomson, J.D. Pollination syndromes and floral specialization. Annu. Rev. Ecol. Syst. 2004, 35, 375–403. [Google Scholar] [CrossRef]
- Dafni, A. Mimicry and deception in pollination. Annu. Rev. Ecol. Syst. 1984, 15, 259–278. [Google Scholar] [CrossRef]
- Reinhard, J.; Srinivasan, M.V.; Guez, D.; Zhang, S.W. Floral scents induce recall of navigational and visual memories in honeybees. J. Exp. Biol. 2004, 207, 4371–4381. [Google Scholar] [CrossRef] [PubMed]
- Morawetz, L.; Svoboda, A.; Spaethe, J.; Dyer, A.G. Blue colour preference in honeybees distracts visual attention for learning closed shapes. J. Comp. Physiol. A 2013, 199, 817–827. [Google Scholar] [CrossRef]
- Giurfa, M.; Eichmann, B.; Menzel, R. Symmetry perception in an insect. Nature 1996, 382, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Giurfa, M.; Vorobyev, M.; Brandt, R.; Posner, B.; Menzel, R. Discrimination of coloured stimuli by honeybees: Alternative use of achromatic and chromatic signals. J. Comp. Physiol. A 1997, 180, 235–243. [Google Scholar] [CrossRef]
- Hempel de Ibarra, N.; Giurfa, M.; Vorobyev, M. Detection of coloured patterns by honeybees through chromatic and achromatic cues. J. Comp. Physiol. A 2001, 187, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Chittka, L.; Menzel, R. The evolutionary adaptation of flower colors and the insect pollinators’ color vision systems. J. Comp. Physiol. A 1992, 171, 171–181. [Google Scholar] [CrossRef]
- Dyer, A.G.; Boyd-Gerny, S.; McLoughlin, S.; Rosa, M.G.P.; Simonov, V.; Wong, B.B.M. Parallel evolution of angiosperm colour signals: Common evolutionary pressures linked to hymenopteran vision. Proc. R. Soc. Biol. Sci. B 2012, 279, 3606–3615. [Google Scholar] [CrossRef]
- Shrestha, M.; Dyer, A.G.; Bhattarai, P.; Burd, M. Flower colour and phylogeny along an altitudinal gradient in the Himalayas of Nepal. J. Ecol. 2014, 102, 126–135. [Google Scholar] [CrossRef]
- Briscoe, A.D.; Chittka, L. The evolution of color vision in insects. Annu. Rev. Entomol. 2001, 46, 471–510. [Google Scholar] [CrossRef] [PubMed]
- Chittka, L.; Thomson, J.D.; Waser, N.M. Flower constancy, insect psychology, and plant evolution. Naturwissenschaften 1999, 86, 361–377. [Google Scholar] [CrossRef]
- Menzel, R. Behavioral access to short-term memory in bees. Nature 1979, 281, 368–369. [Google Scholar] [CrossRef] [PubMed]
- Dyer, A.G.; Chittka, L. Biological significance of distinguishing between similar colours in spectrally variable illumination: Bumbleebees (Bombus terrestris) as a case study. J. Comp. Physiol. A 2004, 190, 105–114. [Google Scholar] [CrossRef]
- Peitsch, D.; Fietz, A.; Hertel, H.; Souza, J.; Ventura, D.F.; Menzel, R. The spectral input systems of hymenopteran insects and their receptor-based colour vision. J. Comp. Physiol. A 1992, 170, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Von Frisch, K. Der Farbensinn und Formensinn der Biene. Zool. Jb. Abt. Allg. Zool. Physiol. 1914, 35, 1–188. (in German). [Google Scholar]
- Daumer, K. Reizmetrische Untersuchungen des Farbensehens der Bienen. Z. Vergl. Physiol. 1956, 38, 413–478. (in German). [Google Scholar]
- Von Helversen, O. Zur spektralen Unterschiedsempfindlichkeit der Honigbiene. J. Comp. Physiol. A 1972, 80, 439–472. (in German). [Google Scholar] [CrossRef]
- Menzel, R. Untersuchungen zum Erlernen von Spektralfarben durch die Honigbiene (Apis mellifica). Z. Vergl. Physiol. 1967, 56, 22–62. (in German). [Google Scholar] [CrossRef]
- Giurfa, M.; Nunez, J.; Chittka, L.; Menzel, R. Colour preference of flower-naive honeybees. J. Comp. Physiol. A 1995, 177, 247–259. [Google Scholar] [CrossRef]
- Dyer, A.G.; Chittka, L. Fine colour discrimination requires differential conditioning in bumblebees. Naturwissenschaften 2004, 91, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Giurfa, M. Conditioning procedure and color discrimination in the honeybee Apis mellifera. Naturwissenschaften 2004, 91, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Dyer, A.G.; Murphy, A.H. Honeybees choose “incorrect” colors that are similar to target flowers in preference to novel colors. Isr. J. Plant Sci. 2009, 57, 203–210. [Google Scholar] [CrossRef]
- Menzel, R. Das Gedächtnis der Honigbiene für Spektralfarben. I. Kurzzeitiges und langzeitiges Behalten. Z. Vergl. Physiol. 1968, 60, 82–102. (in German). [Google Scholar]
- Menzel, R. Memory dynamics in the honeybee. J. Comp. Physiol. A 1999, 185, 323–340. [Google Scholar] [CrossRef]
- Reinhard, J.; Srinivasan, M.V.; Zhang, S. Complex memories in honeybees: Can there be more than two? J. Comp. Physiol. A 2006, 192, 409–416. [Google Scholar]
- Dyer, A.G.; Paulk, A.C.; Reser, D.H. Colour processing in complex environments: Insights from the visual system of bees. Proc. R. Soc. Biol. Sci. B 2011, 278, 952–959. [Google Scholar] [CrossRef]
- Dyer, A.G.; Dorin, A.; Reinhardt, V.; Garcia, J.E.; Rosa, M.G.P. Bee reverse-learning behavior and intra-colony differences: Simulations based on behavioural experiments reveal benefits of diversity. Ecol. Model. 2014, 277, 119–131. [Google Scholar] [CrossRef]
- Chittka, L. The color hexagon: A chromaticity diagram based on photoreceptor excitations as a generalized representation of color opponency. J. Comp. Physiol. A 1992, 170, 533–543. [Google Scholar]
- Dyer, A.G.; Neumeyer, C. Simultaneous and successive colour discrimination in the honeybee (Apis mellifera). J. Comp. Physiol. A 2005, 191, 547–557. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing; v 3.0.2; R Core Team; R Foundation for Statistical Computing: Vienna, Austria, 2013.
- Reser, D.H.; Wijesekara Witharanage, R.; Rosa, M.G.P.; Dyer, A.G. Honeybees (Apis mellifera) learn color discriminations via differential conditioning independent of long wavelength (green) photoreceptor modulation. PLoS One 2012, 7, e48577. [Google Scholar] [CrossRef] [PubMed]
- Paulk, A.C.; Dacks, A.M.; Phillips-Portillo, J.; Fellous, J.M.; Gronenberg, W. Visual processing in the central bee brain. J. Neurosci. 2009, 29, 9987–9999. [Google Scholar] [CrossRef] [PubMed]
- Paulk, A.C.; Phillips-Portillo, J.; Dacks, A.M.; Fellous, J.; Gronenberg, W. The processing of colour, motion, and stimulus timing are anatomically segregated in the bumblebee brain. J. Neurosci. 2008, 28, 6319–6332. [Google Scholar]
- Burns, J.G.; Dyer, A.G. Diversity of speed accuracy strategies benefits social insects. Curr. Biol. 2008, 18, R953–R954. [Google Scholar] [CrossRef] [PubMed]
- Kelber, A. What a hawkmoth remembers after hibernation depends on innate preferences and conditioning situation. Behav. Ecol. 2010, 21, 1093–1097. [Google Scholar] [CrossRef]
- Balkenius, A.; Kelber, A.; Balkenius, C. A model of selection between stimulus and place strategy in a hawkmoth. Adapt. Behav. 2004, 12, 21–35. [Google Scholar] [CrossRef]
- Avargue`s-Weber, A.; de Brito Sanchez, M.G.; Giurfa, M.; Dyer, A.G. Aversive reinforcement improves visual discrimination learning in free flying honeybees. PLoS One 2010, 5, e15370. [Google Scholar]
- Rodríguez-Gironés, M.A.; Trillo, A.; Corcobado, G. Long Term Effects of Aversive Reinforcement on Colour Discrimination Learning in Free-Flying Bumblebees. PLoS One 2013, 8, e71551. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dyer, A.G.; Garcia, J.E. Color Difference and Memory Recall in Free-Flying Honeybees: Forget the Hard Problem. Insects 2014, 5, 629-638. https://doi.org/10.3390/insects5030629
Dyer AG, Garcia JE. Color Difference and Memory Recall in Free-Flying Honeybees: Forget the Hard Problem. Insects. 2014; 5(3):629-638. https://doi.org/10.3390/insects5030629
Chicago/Turabian StyleDyer, Adrian G., and Jair E. Garcia. 2014. "Color Difference and Memory Recall in Free-Flying Honeybees: Forget the Hard Problem" Insects 5, no. 3: 629-638. https://doi.org/10.3390/insects5030629
APA StyleDyer, A. G., & Garcia, J. E. (2014). Color Difference and Memory Recall in Free-Flying Honeybees: Forget the Hard Problem. Insects, 5(3), 629-638. https://doi.org/10.3390/insects5030629